
Improving Ergometrine Production by easO and easP Knockout in Claviceps paspali


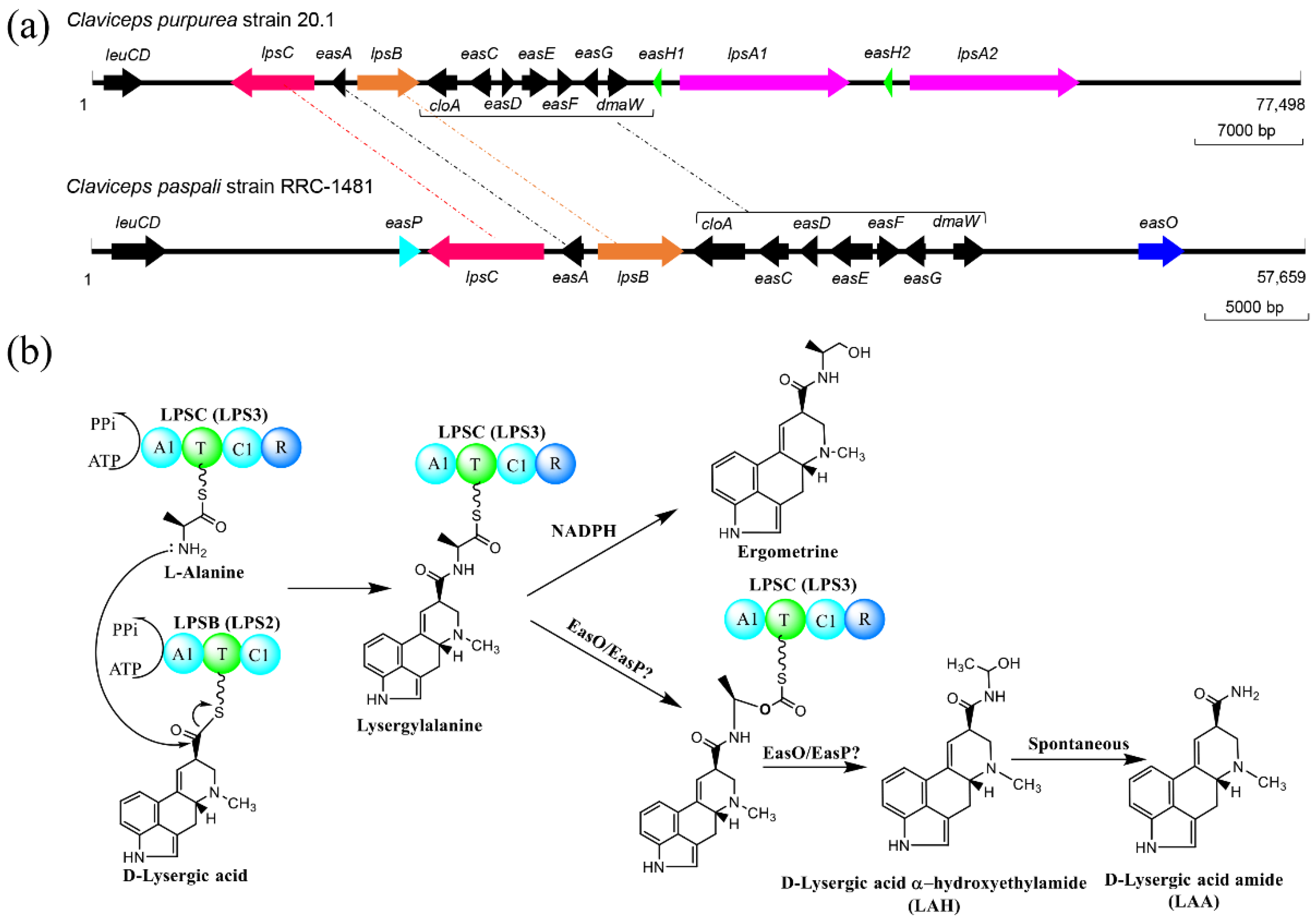{kind=link}
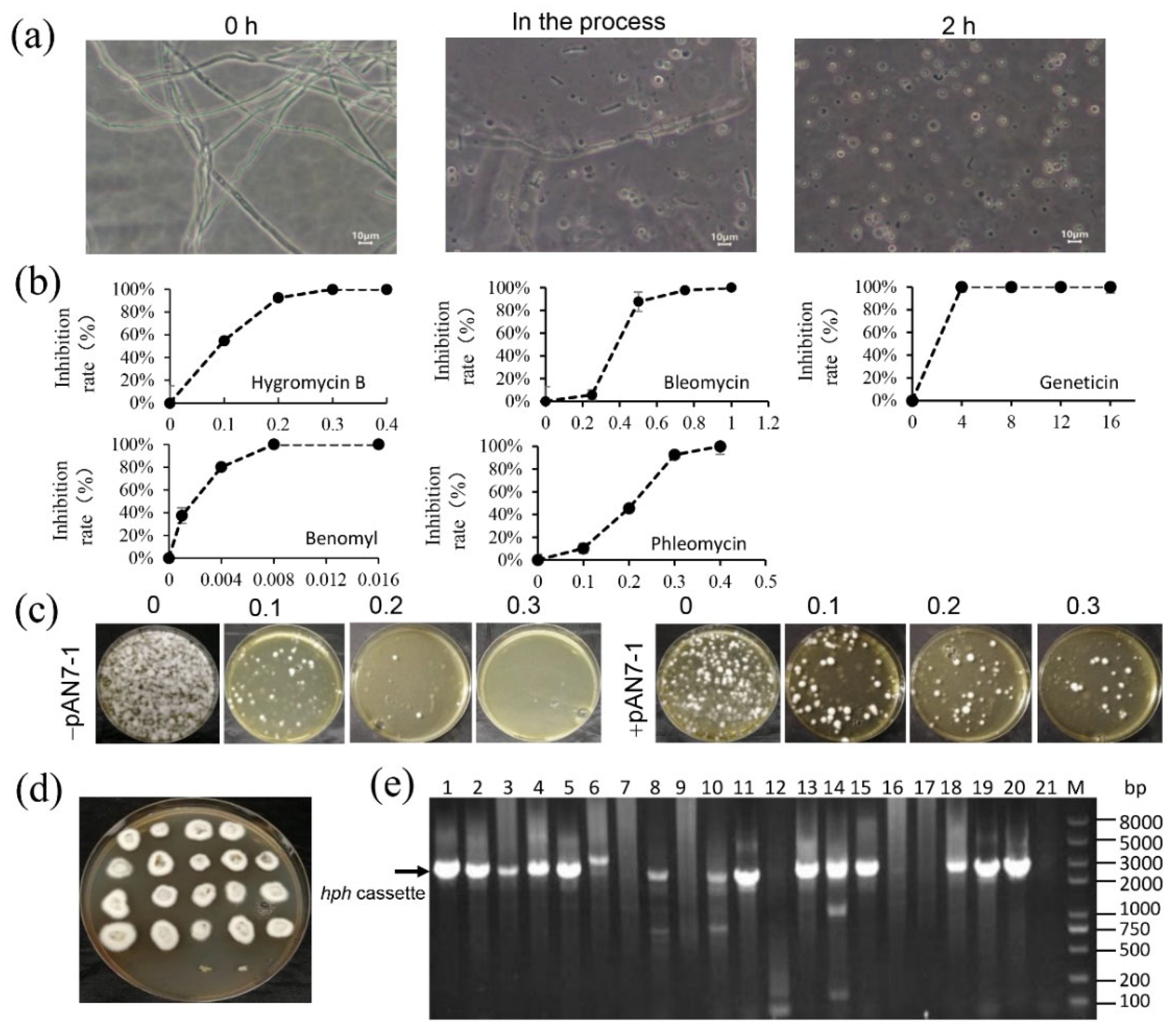{kind=link}
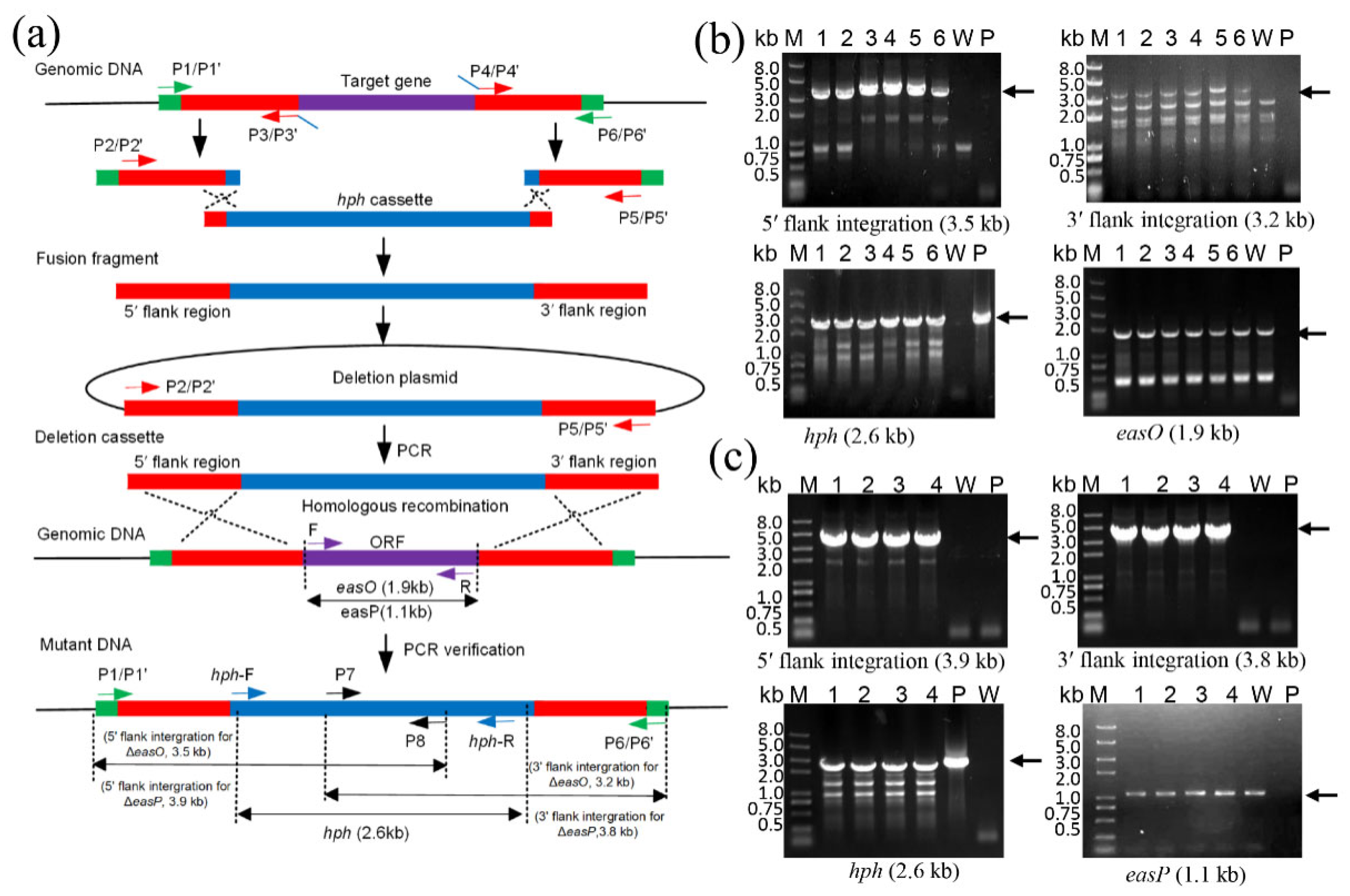{kind=link}
{kind=link}
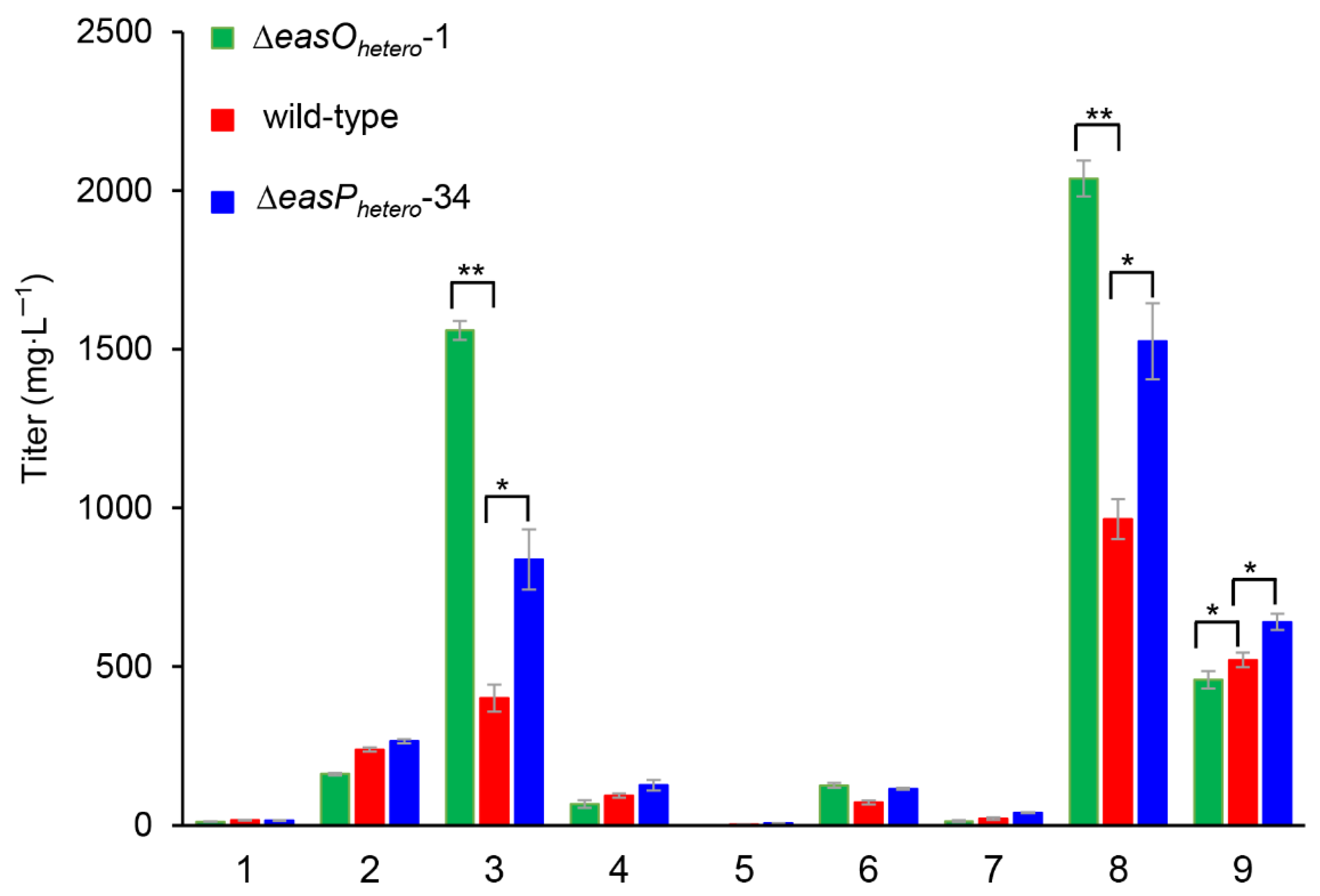{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Establishment of the Protoplast-Mediated Transformation System
2.3. Construction of Deletion Cassettes and Characterization of ΔeasO and ΔeasP Mutants
2.4. Detection and Analysis of Ergot Alkaloid Production for C. paspali Mutants
2.5. Heterologous Expression in A. nidulans A1145
2.6. Product Analysis for Transformants of A. nidulans
2.7. Statistical Analysis
3. Results
3.1. The Protoplast-Mediated Transformation of C. paspali
3.2. Knockout of easO and easP and Characterization of ΔeasO and ΔeasP Mutants
3.3. Analysis of Ergot Alkaloid Production
3.4. Heterologous Expression of the Ergot Alkaloid Biosynthetic Genes in A. nidulans
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doi, Y.; Wakana, D.; Takeda, H.; Tanaka, E.; Hosoe, T. Production of Ergot Alkaloids by the Japanese Isolate Claviceps purpurea Var. Agropyri on Rice Medium. Adv. Microbiol. 2022, 12, 254–269. [Google Scholar] [CrossRef]
- Wallwey, C.; Li, S. Ergot Alkaloids: Structure Diversity, Biosynthetic Gene Clusters and Functional Proof of Biosynthetic Genes. Nat. Prod. Rep. 2011, 28, 496–510. [Google Scholar] [CrossRef]
- Motti, R.; Bonanomi, G.; Emrick, S.; Lanzotti, V. Traditional Herbal Remedies Used in women’s Health Care in Italy: A Review. Hum. Ecol. 2019, 47, 941–972. [Google Scholar] [CrossRef]
- Hulvová, H.; Galuszka, P.; Frébortová, J.; Frébort, I. Parasitic Fungus Claviceps as a Source for Biotechnological Production of Ergot Alkaloids. Biotechnol. Adv. 2013, 31, 79–89. [Google Scholar] [CrossRef]
- Ong, J.J.Y.; De Felice, M. Migraine Treatment: Current Acute Medications and their Potential Mechanisms of Action. Neurotherapeutics 2018, 15, 274–290. [Google Scholar] [CrossRef] [Green Version]
- Bonuccelli, U.; Del Dotto, P.; Rascol, O. Role of Dopamine Receptor Agonists in the Treatment of Early Parkinson’s Disease. Parkinsonism Relat Disord. 2009, 15, S44–S53. [Google Scholar] [CrossRef]
- De Deurwaerdère, P.; Di Giovanni, G.; Millan, M.J. Expanding the Repertoire of L-DOPA’s Actions: A Comprehensive Review of its Functional Neurochemistry. Prog. Neurobiol. 2017, 151, 57–100. [Google Scholar] [CrossRef]
- Agriopoulou, S. Ergot Alkaloids Mycotoxins in Cereals and Cereal-Derived Food Products: Characteristics, Toxicity, Prevalence, and Control Strategies. Agronomy 2021, 11, 931. [Google Scholar] [CrossRef]
- Ortel, I.; Keller, U. Combinatorial Assembly of Simple and Complex D-lysergic Acid Alkaloid Peptide Classes in the Ergot Fungus Claviceps purpurea. J. Biol. Chem. 2009, 284, 6650–6660. [Google Scholar] [CrossRef] [Green Version]
- Young, C.A.; Schardl, C.L.; Panaccione, D.G.; Florea, S.; Takach, J.E.; Charlton, N.D.; Moore, N.; Webb, J.S.; Jaromczyk, J. Genetics, Genomics and Evolution of Ergot Alkaloid Diversity. Toxins 2015, 7, 1273–1302. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Jia, Y. Ergot Alkaloids: Synthetic Approaches to Lysergic Acid and Clavine Alkaloids. Nat. Prod. Rep. 2017, 34, 411–432. [Google Scholar] [CrossRef]
- Oberti, H.; Dalla Rizza, M.; Reyno, R.; Murchio, S.; Altier, N.; Abreo, E. Diversity of Claviceps paspali Reveals Unknown Lineages and Unique Alkaloid Genotypes. Mycologia 2020, 112, 230–243. [Google Scholar] [CrossRef]
- Uhlig, S.; Rangel-Huerta, O.D.; Divon, H.H.; Rolén, E.; Pauchon, K.; Sumarah, M.W.; Vrålstad, T.; Renaud, J.B. Unraveling the Ergot Alkaloid and Indole Diterpenoid Metabolome in the Claviceps purpurea Species Complex Using LC–HRMS/MS Diagnostic Fragmentation Filtering. J. Agric. Food Chem. 2021, 69, 7137–7148. [Google Scholar] [CrossRef]
- Coyle, C.M.; Cheng, J.Z.; O’Connor, S.E.; Panaccione, D.G. An Old Yellow Enzyme Gene Controls the Branch Point Between Aspergillus fumigatus and Claviceps purpurea Ergot Alkaloid Pathways. Appl. Environ. Microbiol. 2010, 76, 3898–3903. [Google Scholar] [CrossRef] [Green Version]
- Králová, M.; Frébortová, J.; Pěnčík, A.; Frébort, I. Overexpression of Trp-related Genes in Claviceps purpurea Leading to Increased Ergot Alkaloid Production. New Biotechnol. 2021, 61, 69–79. [Google Scholar] [CrossRef]
- Haarmann, T.; Ortel, I.; Tudzynski, P.; Keller, U. Identification of the Cytochrome P450 Monooxygenase that Bridges the Clavine and Ergoline Alkaloid Pathways. Chembiochem 2006, 7, 645–652. [Google Scholar] [CrossRef]
- Hicks, C.; Witte, T.E.; Sproule, A.; Lee, T.; Shoukouhi, P.; Popovic, Z.; Menzies, J.G.; Boddy, C.N.; Liu, M.; Overy, D.P. Evolution of the Ergot Alkaloid Biosynthetic Gene Cluster Results in Divergent Mycotoxin Profiles in Claviceps purpurea Sclerotia. Toxins 2021, 13, 861. [Google Scholar] [CrossRef]
- Florea, S.; Panaccione, D.G.; Schardl, C.L. Ergot Alkaloids of the Family Clavicipitaceae. Phytopathology 2017, 107, 504–518. [Google Scholar] [CrossRef] [Green Version]
- Wong, G.; Lim, L.R.; Tan, Y.Q.; Go, M.K.; Bell, D.J.; Freemont, P.S.; Yew, W.S. Reconstituting the Complete Biosynthesis of D-lysergic Acid in Yeast. Nat. Commun. 2022, 13, 712. [Google Scholar] [CrossRef]
- Yan, L.; Liu, Y. Insights into the Mechanism and Enantioselectivity in the Biosynthesis of Ergot Alkaloid Cycloclavine Catalyzed by Aj_EasH From Aspergillus japonicus. Inorg. Chem. 2019, 58, 13771–13781. [Google Scholar] [CrossRef]
- Furukawa, T.; van Rhijn, N.; Fraczek, M.; Gsaller, F.; Davies, E.; Carr, P.; Gago, S.; Fortune-Grant, R.; Rahman, S.; Gilsenan, J.M.; et al. The Negative Cofactor 2 Complex is a Key Regulator of Drug Resistance in Aspergillus fumigatus. Nat. Commun. 2020, 11, 427. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, S.; Matsubara, Y.; Watanabe, K. Alkaloid Biosynthetic Enzyme Generates Diastereomeric Pair via Two Distinct Mechanisms. J. Am. Chem. Soc. 2022, 144, 5485–5493. [Google Scholar] [CrossRef]
- Jones, A.M.; Steen, C.R.; Panaccione, D.G. Independent Evolution of a Lysergic Acid Amide in Aspergillus species. Appl. Environ. Microbiol. 2021, 87, e1801–e1821. [Google Scholar] [CrossRef]
- Davis, K.A.; Sampson, J.K.; Panaccione, D.G. Genetic Reprogramming of the Ergot Alkaloid Pathway of Metarhizium brunneum. Appl. Environ. Microbiol. 2020, 86, e01251-20. [Google Scholar] [CrossRef]
- Steen, C.R.; Sampson, J.K.; Panaccione, D.G.; Druzhinina, I.S. A Baeyer-Villiger Monooxygenase Gene Involved in the Synthesis of Lysergic Acid Amides Affects the Interaction of the Fungus Metarhizium brunneum with Insects. Appl. Environ. Microbiol. 2021, 87, e721–e748. [Google Scholar] [CrossRef]
- Tang, M.; He, H.; Zhang, F.; Tang, G. Baeyer-Villiger Oxidation of Acyl Carrier Protein-Tethered Thioester to Acyl Carrier Protein-Linked Thiocarbonate Catalyzed by a Monooxygenase Domain in FR901464 Biosynthesis. ACS Catal. 2013, 3, 444–447. [Google Scholar] [CrossRef]
- Tolmie, C.; Smit, M.S.; Opperman, D.J. Native Roles of Baeyer-Villiger Monooxygenases in the Microbial Metabolism of Natural Compounds. Nat. Prod. Rep. 2019, 36, 326–353. [Google Scholar] [CrossRef]
- Castagnoli, N.J.; Corbett, K.; Chain, E.B.; Thomas, R. Biosynthesis of N-(α-hydroxyethyl)-lysergamide, a Metabolite of Claviceps paspali Stevens & Hall. Biochem. J. 1970, 117, 451–455. [Google Scholar] [CrossRef]
- Pertot, E.; Gaberc-porekar, V.; Socic, H. Isolation and Characterization of an Alkaloid-Blocked Mutant of Claviceps paspali. J. Basic Microbiol. 1990, 30, 51–56. [Google Scholar] [CrossRef]
- Koide, R.T. Methods in Enzymology; Zimmer, E.A., Roalson, E.H., Eds.; Academic Press: New York, NY, USA, 2005; pp. 58–72. [Google Scholar]
- Kozák, L.; Szilágyi, Z.; Vágó, B.; Kakuk, A.; Tóth, L.; Molnár, I.; Pócsi, I. Inactivation of the Indole-Diterpene Biosynthetic Gene Cluster of Claviceps paspali by Agrobacterium-mediated Gene Replacement. Appl. Microbiol. Biotechnol. 2018, 102, 3255–3266. [Google Scholar] [CrossRef]
- Bai, J.; Yan, D.; Zhang, T.; Guo, Y.; Liu, Y.; Zou, Y.; Tang, M.; Liu, B.; Wu, Q.; Yu, S.; et al. A Cascade of Redox Reactions Generates Complexity in the Biosynthesis of the Protein Phosphatase-2 Inhibitor RubratoxinA. Angew. Chem. Int. Ed. 2017, 56, 4782–4786. [Google Scholar] [CrossRef]
- Szewczyk, E.; Nayak, T.; Oakley, C.E.; Edgerton, H.; Xiong, Y.; Taheri-Talesh, N.; Osmani, S.A.; Oakley, B.R. Fusion PCR and Gene Targeting in Aspergillus nidulans. Nat. Protoc. 2006, 1, 3111–3120. [Google Scholar] [CrossRef]
- Ohashi, M.; Liu, F.; Hai, Y.; Chen, M.; Tang, M.; Yang, Z.; Sato, M.; Watanabe, K.; Houk, K.N.; Tang, Y. SAM-dependent Enzyme-Catalysed Pericyclic Reactions in Natural Product Biosynthesis. Nature 2017, 549, 502–506. [Google Scholar] [CrossRef] [Green Version]
- Yan, D.; Chen, Q.; Gao, J.; Bai, J.; Liu, B.; Zhang, Y.; Zhang, L.; Zhang, C.; Zou, Y.; Hu, Y. Complexity and Diversity Generation in the Biosynthesis of Fumiquinazoline-Related Peptidyl Alkaloids. Org. Lett. 2019, 21, 1475–1479. [Google Scholar] [CrossRef]
- Punt, P.J.; Oliver, R.P.; Dingemanse, M.A.; Pouweis, P.H.; van den Handel, C.A.M.J. Transformation of Aspergillus Based On the Hygromycin B Resistance Marker From Escherichia coli. Gene 1987, 56, 117–124. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, Y.-M.; Wen, Y.-H.; Gong, T.; Chen, J.-J.; Chen, T.-J.; Yang, J.-L.; Zhu, P. Improving Ergometrine Production by easO and easP Knockout in Claviceps paspali. Fermentation 2022, 8, 263. https://doi.org/10.3390/fermentation8060263
Qiao Y-M, Wen Y-H, Gong T, Chen J-J, Chen T-J, Yang J-L, Zhu P. Improving Ergometrine Production by easO and easP Knockout in Claviceps paspali. Fermentation. 2022; 8(6):263. https://doi.org/10.3390/fermentation8060263
Chicago/Turabian StyleQiao, Yun-Ming, Yan-Hua Wen, Ting Gong, Jing-Jing Chen, Tian-Jiao Chen, Jin-Ling Yang, and Ping Zhu. 2022. "Improving Ergometrine Production by easO and easP Knockout in Claviceps paspali" Fermentation 8, no. 6: 263. https://doi.org/10.3390/fermentation8060263
APA StyleQiao, Y. -M., Wen, Y. -H., Gong, T., Chen, J. -J., Chen, T. -J., Yang, J. -L., & Zhu, P. (2022). Improving Ergometrine Production by easO and easP Knockout in Claviceps paspali. Fermentation, 8(6), 263. https://doi.org/10.3390/fermentation8060263