
A Novel Salt-Tolerant L-Glutaminase: Efficient Functional Expression, Computer-Aided Design, and Application




Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Source, Plasmids, and Strains
2.2. Bioinformatics Analysis and Screening for Positive Variants
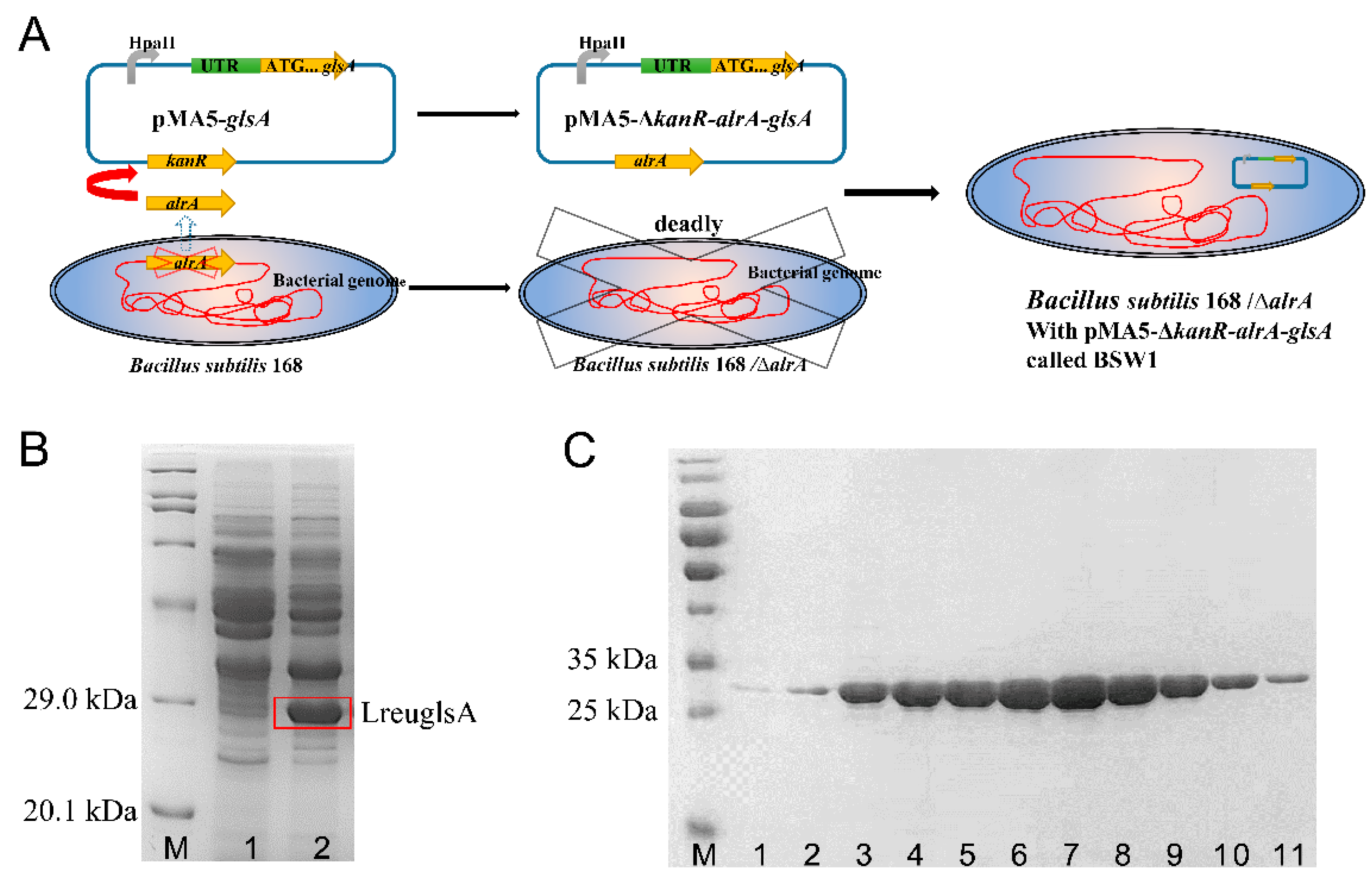
2.3. Construction of a Food Safe Recombinant B. Subtilis 168 Strain
2.4. Overexpression through Synthetic Biology Components
2.5. L-Glutaminase Expression and Purification
2.6. Enzyme Activity Assay
2.7. Fermentation and Application of L-Glutaminase in Soy Sauce Brewing
3. Results and Discussion
3.1. A Novel L-Glutaminase Displays Significant Salt Tolerance
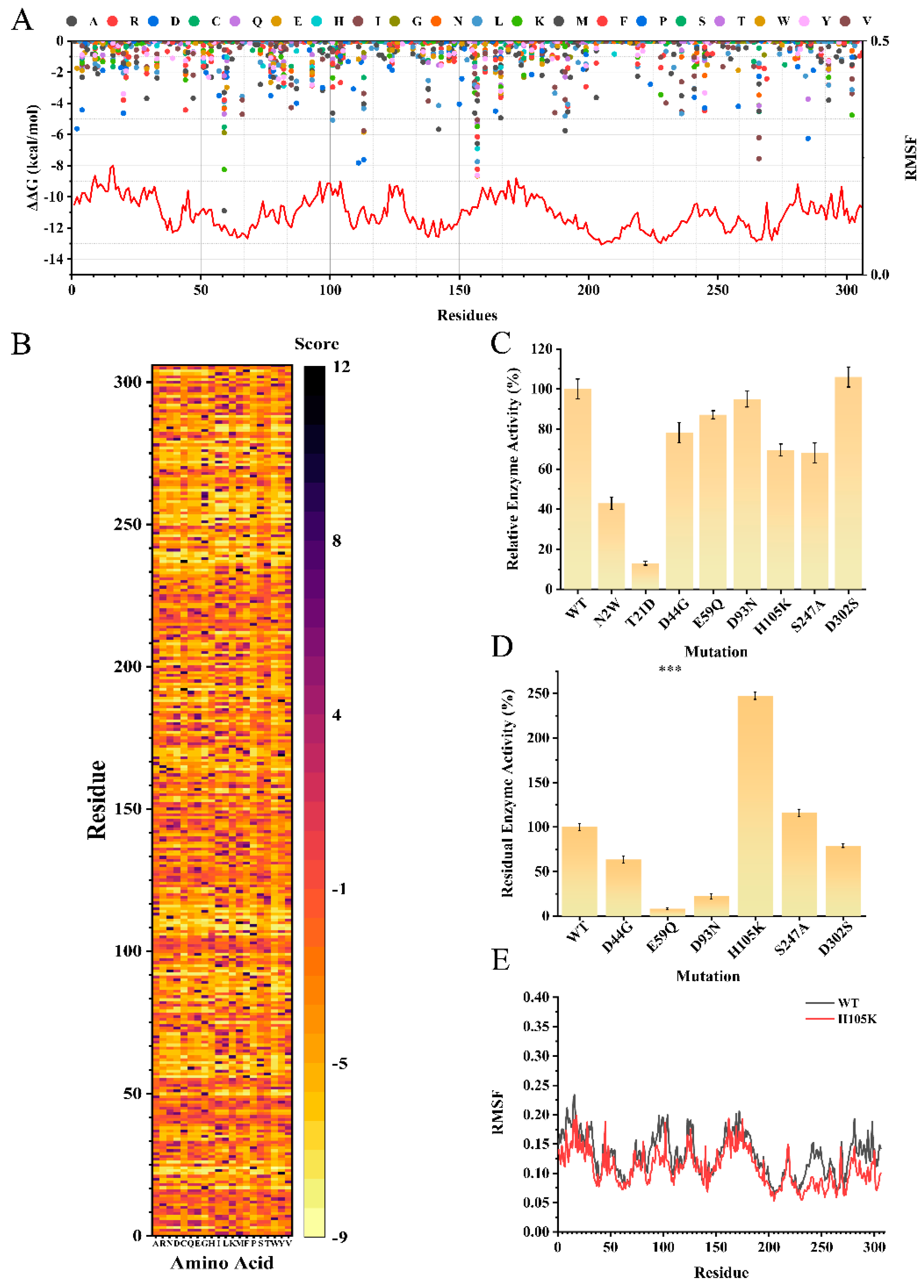
3.2. Structural Prediction and Identification of the Active Center of LreuglsA
3.3. A H105K Mutant Is Designed Using Computer-Aided Mutation Prediction to Increase Thermostability
3.4. Improving Enzyme Production with a Portable 5′-UTR
3.5. Immobilized BSW3-H105K Cells Significantly Increase Glutamate Production during Soy Sauce Brewing
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Halpern, B.P. Glutamate and the flavor of foods. J. Nutr. 2000, 130, 910S–914S. [Google Scholar] [CrossRef] [PubMed]
- Geha, R.S.; Beiser, A.; Ren, C.; Patterson, R.; Greenberger, P.A.; Grammer, L.C.; Ditto, A.M.; Harris, K.E.; Shaughnessy, M.A.; Yarnold, P.R.; et al. Review of alleged reaction to monosodium glutamate and outcome of a multicenter double-blind placebo-controlled study. J. Nutr. 2000, 130, 1058S–1062S. [Google Scholar] [CrossRef] [PubMed]
- Graudal, N.A.; Hubeck-Graudal, T.; Jurgens, G. Effects of low sodium diet versus high sodium diet on blood pressure, renin, aldosterone, catecholamines, cholesterol, and triglyceride. Cochrane Database Syst. Rev. 2020, 12, CD004022. [Google Scholar] [CrossRef] [PubMed]
- Binod, P.; Sindhu, R.; Madhavan, A.; Abraham, A.; Mathew, A.K.; Beevi, U.S.; Sukumaran, R.K.; Singh, S.P.; Pandey, A. Recent developments in l-glutaminase production and applications—An overview. Bioresour. Technol. 2017, 245, 1766–1774. [Google Scholar] [CrossRef]
- Nandakumar, R.; Yoshimune, K.; Wakayama, M.; Moriguchi, M. Microbial glutaminase: Biochemistry, molecular approaches and applications in the food industry. J. Mol. Catal. B Enzym. 2003, 23, 87–100. [Google Scholar] [CrossRef]
- Tadanobu Nakadai, S.N. Use of Glutaminase for Soy Sauce Made by Koji or a Preparation of Proteases from Aspergillus oryzae. J. Ferment. Bioeng. 1989, 67, 158–162. [Google Scholar] [CrossRef]
- Kumar, L.; Singh, B.; Adhikari, D.K.; Mukherjee, J.; Ghosh, D. A temperature and salt-tolerant L-glutaminase from gangotri region of uttarakhand himalaya: Enzyme purification and characterization. Appl. Biochem. Biotechnol. 2012, 166, 1723–1735. [Google Scholar] [CrossRef]
- Nandakumar, R.; Wakayama, M.; Nagano, Y.; Kawamura, T.; Sakai, K.; Moriguchi, M. Overexpression of salt-tolerant glutaminase from Micrococcus luteus K-3 in Escherichia coli and its purification. Protein Expr. Purif. 1999, 15, 155–161. [Google Scholar] [CrossRef]
- Moriguchi, M.; Sakai, K.; Tateyama, R.; Furuta, Y.; Wakayama, M. Isolation and Characterization of Salt-Tolerant Glutaminases from Marine Micrococcus-Luteus K-3. J. Ferment. Bioeng. 1994, 77, 621–625. [Google Scholar] [CrossRef]
- Weingand-Ziade, A.; Gerber-Decombaz, C.; Affolter, M. Functional characterization of a salt- and thermotolerant glutaminase from Lactobacillus rhamnosus. Enzym. Microb. Technol. 2003, 32, 862–867. [Google Scholar] [CrossRef]
- Jeon, J.M.; Lee, H.I.; Han, S.H.; Chang, C.S.; So, J.S. Partial purification and characterization of glutaminase from Lactobacillus reuteri KCTC3594. Appl. Biochem. Biotechnol. 2010, 162, 146–154. [Google Scholar] [CrossRef]
- Amobonye, A.; Singh, S.; Pillai, S. Recent advances in microbial glutaminase production and applications-a concise review. Crit. Rev. Biotechnol. 2019, 39, 944–963. [Google Scholar] [CrossRef] [PubMed]
- Pu, H.; Wang, Q.; Zhu, F.; Cao, X.; Xin, Y.; Luo, L.; Yin, Z. Cloning, expression of glutaminase fromPseudomonas nitroreducensand application to theanine synthesis. Biocatal. Biotransform. 2012, 31, 1–7. [Google Scholar] [CrossRef]
- Sakhaei, M.; Alemzadeh, I. Enzymatic Synthesis of Theanine in the Presence of L-glutaminase Produced by Trichoderma koningii. Appl. Food Biotechnol. 2017, 4, 113–121. [Google Scholar] [CrossRef]
- Kroemer, G.; Pouyssegur, J. Tumor cell metabolism: Cancer’s Achilles’ heel. Cancer Cell 2008, 13, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Orabi, H.; El-Fakharany, E.; Abdelkhalek, E.; Sidkey, N. Production, optimization, purification, characterization, and anti-cancer application of extracellular L-glutaminase produced from the marine bacterial isolate. Prep. Biochem. Biotechnol. 2020, 50, 408–418. [Google Scholar] [CrossRef]
- Reda, F.M. Kinetic properties of Streptomyces canarius L- Glutaminase and its anticancer efficiency. Braz. J. Microbiol. 2015, 46, 957–968. [Google Scholar] [CrossRef]
- Liu, Y.; Su, A.; Li, J.; Ledesma-Amaro, R.; Xu, P.; Du, G.; Liu, L. Towards next-generation model microorganism chassis for biomanufacturing. Appl. Microbiol. Biotechnol. 2020, 104, 9095–9108. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, L.; Li, J.; Du, G.; Chen, J. Synthetic Biology Toolbox and Chassis Development in Bacillus subtilis. Trends Biotechnol. 2019, 37, 548–562. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Evans, R.; O’Neill, M.; Pritzel, A.; Antropova, N.; Senior, A.; Green, T.; Žídek, A.; Bates, R.; Blackwell, S.; Yim, J.; et al. Protein complex prediction with AlphaFold-Multimer. BioRxiv 2022. [Google Scholar] [CrossRef]
- Schymkowitz, J.; Borg, J.; Stricher, F.; Nys, R.; Rousseau, F.; Serrano, L. The FoldX web server: An online force field. Nucleic Acids Res. 2005, 33, W382–W388. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Liu, Q.; Qu, G.; Feng, Y.; Reetz, M.T. Utility of B-Factors in Protein Science: Interpreting Rigidity, Flexibility, and Internal Motion and Engineering Thermostability. Chem. Rev. 2019, 119, 1626–1665. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Xia, Y.; Chen, W.; Zhao, J.X.; Tian, F.W.; Zhang, H.; Ding, X.L. Construction of a new food-grade expression system for Bacillus subtilis based on theta replication plasmids and auxotrophic complementation. Appl. Microbiol. Biotechnol. 2007, 76, 643–650. [Google Scholar] [CrossRef]
- Schwanhausser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global quantification of mammalian gene expression control. Nature 2011, 473, 337–342, Erratum in Nature 2013, 495, 126–127. [Google Scholar] [CrossRef]
- Kozak, M. Influences of mRNA secondary structure on initiation by eukaryotic ribosomes. Proc. Natl. Acad. Sci. USA 1986, 83, 2850–2854. [Google Scholar] [CrossRef]
- Viegas, S.C.; Apura, P.; Martinez-Garcia, E.; de Lorenzo, V.; Arraiano, C.M. Modulating Heterologous Gene Expression with Portable mRNA-Stabilizing 5′-UTR Sequences. ACS Synth. Biol. 2018, 7, 2177–2188. [Google Scholar] [CrossRef]
- Xiao, J.; Peng, B.; Su, Z.; Liu, A.; Hu, Y.; Nomura, C.T.; Chen, S.; Wang, Q. Facilitating Protein Expression with PorTable 5′-UTR Secondary Structures in Bacillus licheniformis. ACS Synth. Biol. 2020, 9, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Hofacker, I.L.; Stadler, P.F. Memory efficient folding algorithms for circular RNA secondary structures. Bioinformatics 2006, 22, 1172–1176. [Google Scholar] [CrossRef]
- Codex Alimentarius Commission. Guideline for the Conduct of Food Safety Assessment of Foods Derived from Recombinant-DNA Plants; Codex Alimentarius Commission: Rome, Italy, 2003. [Google Scholar]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [PubMed]
- Oliva, M.; Dideberg, O.; Field, M.J. Understanding the acylation mechanisms of active-site serine penicillin-recognizing proteins: A molecular dynamics simulation study. Proteins 2003, 53, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.C.; Sim, D.H.; Seo, M.J.; Oh, D.K. Increased Production of Food-Grade d-Tagatose from d-Galactose by Permeabilized and Immobilized Cells of Corynebacterium glutamicum, a GRAS Host, Expressing d-Galactose Isomerase from Geobacillus thermodenitrificans. J. Agric. Food Chem. 2016, 64, 8146–8153. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Optimum pH | Optimum Temp (°C) | Specific Gravity (U/mg) | Thermal Stability | Salt Tolerant | References |
|---|---|---|---|---|---|---|
| A. oryzae | 9 | 41 | 40.12 | NR | + | [6] |
| Bacillus sp. LKG-01 | 11 | 70 | 584.2 | +++ | +++ | [7] |
| M. luteus K-3 | 8 | 50 | 190 | + | +++ | [8] |
| L. rhamnosus | 7 | 50 | NR | ++ | +++ | [10] |
| L. reuteri KCTC3594 | 7.5 | 40 | 16.4 | + | ++ | [11] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Hu, M.; Wang, Q.; Liu, F.; Xu, M.; Zhang, X.; Rao, Z. A Novel Salt-Tolerant L-Glutaminase: Efficient Functional Expression, Computer-Aided Design, and Application. Fermentation 2022, 8, 444. https://doi.org/10.3390/fermentation8090444
Zhang H, Hu M, Wang Q, Liu F, Xu M, Zhang X, Rao Z. A Novel Salt-Tolerant L-Glutaminase: Efficient Functional Expression, Computer-Aided Design, and Application. Fermentation. 2022; 8(9):444. https://doi.org/10.3390/fermentation8090444
Chicago/Turabian StyleZhang, Hengwei, Mengkai Hu, Qing Wang, Fei Liu, Meijuan Xu, Xian Zhang, and Zhiming Rao. 2022. "A Novel Salt-Tolerant L-Glutaminase: Efficient Functional Expression, Computer-Aided Design, and Application" Fermentation 8, no. 9: 444. https://doi.org/10.3390/fermentation8090444
APA StyleZhang, H., Hu, M., Wang, Q., Liu, F., Xu, M., Zhang, X., & Rao, Z. (2022). A Novel Salt-Tolerant L-Glutaminase: Efficient Functional Expression, Computer-Aided Design, and Application. Fermentation, 8(9), 444. https://doi.org/10.3390/fermentation8090444