
Natural Substrates and Culture Conditions to Produce Pigments from Potential Microbes in Submerged Fermentation




Abstract
:1. Introduction
2. Pigmented Microbial Distribution in Evolutionary Perspective
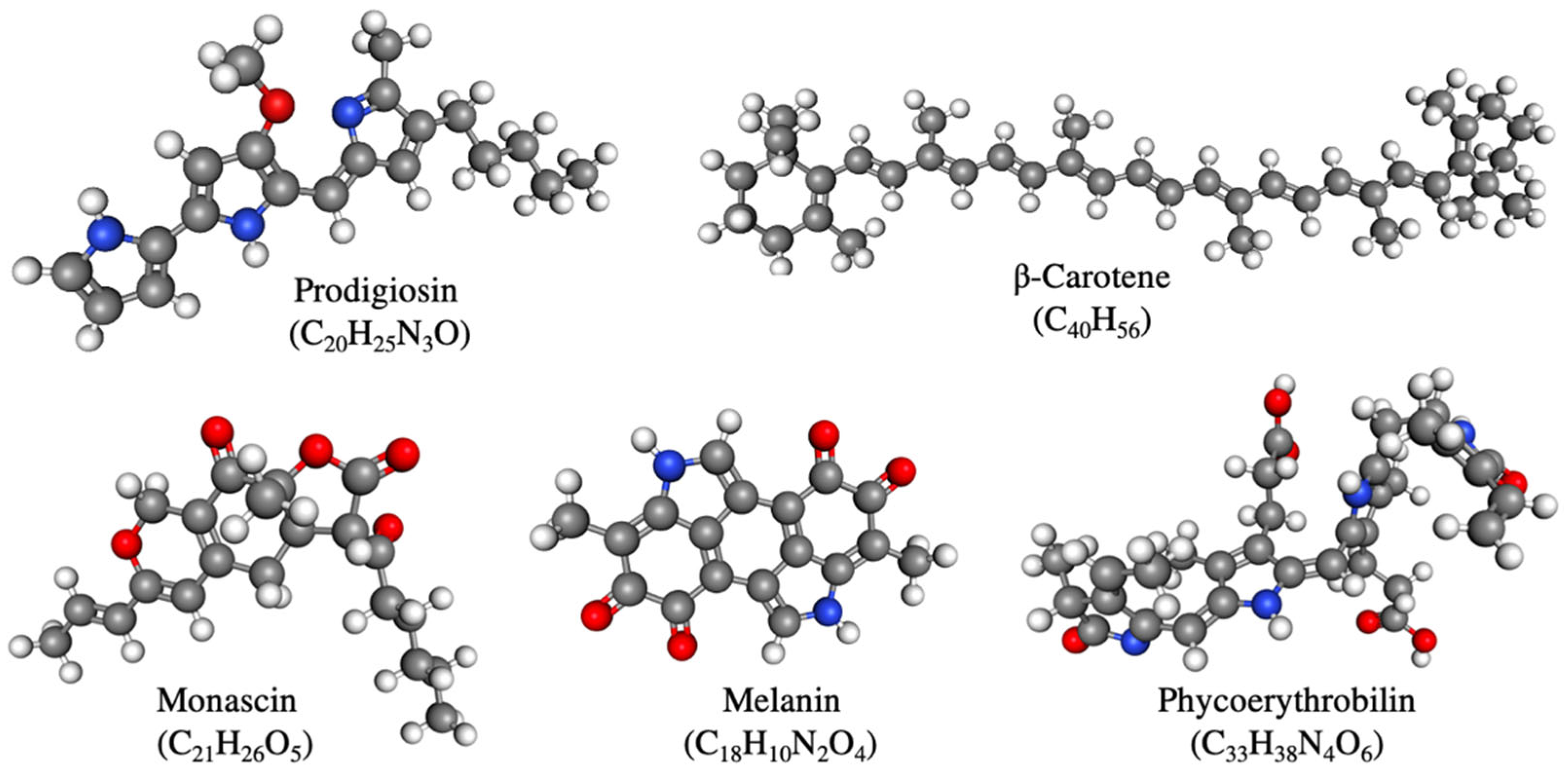
3. Important Pigments for Submerged Fermentation
4. Biosynthetic Pathways of Major Microbial Pigments
5. High Pigment Yielding Natural Substrates

{kind=link}
{kind=link}
{kind=link}
| Pigment Microbe | Species | Substrate | Pigment | Production Rate | Method | Reference |
|---|---|---|---|---|---|---|
| Bacillus safensis | Fruit waste of pineapple, orange, and pomegranate | Melanin | 6.96 mg/mL | Shake flask | [108] | |
| Bacillus subtilis | Corn steep liquor | Riboflavin | 26.8 mg/L | Shake flask | [109] | |
| Bacteria | Chromobacterium vaccinii | Rapeseed cake | Violacein | 12.93 mg/L | Shake flask | [110] |
| Chromobacterium violaceum | Liquid pineapple waste | Violacein | 16.25 mg/mL | 1 L Bioreactor | [111] | |
| Chromobacterium violaceum | Sugarcane bagasse | Violacein | 820 mg/L | Shake flask | [112] | |
| Chryseobacterium artocarpi | Liquid pineapple waste | Flexirubin | 540 mg/L | Shake flask | [113] | |
| Pseudomonas sp. | Vegetable waste | Melanin | 2.79 mg/mL | Shake flask | [114] | |
| Pseudomonas aeruginosa | Cotton seed meal | Pyocyanin | 4 μg/mL | Shake flask | [115] | |
| Pseudomonas aeruginosa | Grape seed | Pyocyanin | 4 μg/mL | Shake flask | [115] | |
| Sarcina sp. | Apple pomace | Carotenoid | 12.87 mg/100g | Shake flask | [116] | |
| Serratia marcescens | Cassava wastewater | Prodigiosin | 49,500 mg/L | Shake flask | [98] | |
| Serratia marcescens | Peanut oil cake | Prodigiosin | 40,000 mg/L | Shake flask | [99] | |
| Serratia marcescens | Tannery fleshing | Prodigiosin | 33,000 mg/L | Shake flask | [117] | |
| Serratia marcescens | Peanut seed broth | Prodigiosin | 38.75 mg/mL | Shake flask | [61] | |
| Serratia marcescens | Peanut seed oil | Prodigiosin | 0.02 gm/mL | Shake flask | [118] | |
| Serratia marcescens | Brown sugar | Prodigiosin | 8 mg/mL | 5 L Bioreactor | [119] | |
| Serratia marcescens | Peanut powder and olive oil | Prodigiosin | 15,420.9 mg/L | Shake flask | [120] | |
| Serratia marcescens | Powdered peanut | Prodigiosin | 1595.09 mg/L | Shake flask | [121] | |
| Serratia marcescens | Wheat bran and sunflower oil | Prodigiosin | 240 mg/L | Shake flask | [122] | |
| Serratia marcescens | Powdered peanut | Prodigiosin | 39 mg/mL | Shake flask | [61] | |
| Serratia marcescens | Sesame seed | Prodigiosin | 17 mg/mL | Shake flask | [61] | |
| Streptomyces sp. | Dairy processing wastewater | Prodigiosin | 47,000 mg/L | Shake flask | [123] | |
| Fungi | Aspergillus carbonarius | Apple, black carrot, pomegranate, red beet pulps | Melanin | 61.84 U/gm | Shake flask | [124] |
| Blakeslea trispora | Cheese whey | Carotenoid | 405 mg/L | 1.4 L glass bioreactor | [125] | |
| Eremothecium gossypii(=Ashbya gossypii) | Corn steep liquor | Riboflavin | 13.7 gm/L | Shake flask | [126] | |
| Monascus purpureus | Rice husk hydrolysate | Monascus | 72.1 U/mL | Shake flask | [127] | |
| Monascus purpureus | Potato pomace | Monascus | 47.9 U/mL | Shake flask | [64] | |
| Monascus purpureus | Whey powder | Monascus | 38.4 U/mL | Shake flask | [128] | |
| Monascus purpureus | Corncob hydrolysate | Monascus | 25.80 U/mL | Shake flask | [100] | |
| Monascus purpureus | Corncob | Monascus | 133.77 U/mL | Shake flask | [129] | |
| Monascus purpureus | Bakery waste hydrolysate | Monascus | 24.01 U/mL | Shake flask | [101] | |
| Monascus purpureus | Brewer’s spent grain | Monascus | 22.25 U/mL | Shake flask | [102] | |
| Monascus purpureus | Soybean meal | Monascus | 21.45 U/mL | Shake flask | [46] | |
| Monascus purpureus | Rice straw hydrolysate with glucose | Monascus | 21.20 U/mL | Shake flask | [103] | |
| Monascus purpureus | Grape waste | Monascus | 20–22.5 gm/L | Shake flask | [130] | |
| Monascus ruber | Sugarcane bagasse hydrolysate | Monascus | 18.71 U/mL | Shake flask | [104] | |
| Penicillium purpurogenum | Orange peels | Monascus-like | 0.58 U/mL | Shake flask | [131] | |
| Sporidiobolus pararoseus | Corn steep liquor | Carotenoid | 40 gm/L | Shake flask | [132] | |
| Talaromyces atroroseus | Corncob hydrolysate | Monascus | 16.17 U/mL | Shake flask | [133] | |
| Talaromyces purpureogenus | Bengal gram husk | Monascus | 0.565 U/mL | Shake flask | [134] | |
| Yeast | Rhodotorula achenorium | Whey ultrafiltrate | Carotenoid | 262 mg/L | Shake flask | [135] |
| Rhodotorula glutinis | Brewery wastewater | Carotenoid | 1.2 mg/L | Shake flask | [136] | |
| Rhodotorula glutinis | Mung bean waste flour and sweet potato extract | Carotenoid | 3.48 mg/L | Shake flask | [137] | |
| Rhodotorula glutinis | Cassava wastewater | Carotenoid | 0.98 mg/L | Shake flask | [138] | |
| Rhodotorula glutinis | Crude glycerol | Carotenoid | 135.25 mg/L | Shake flask | [139] | |
| Rhodotorula glutinis | Chicken feathers | Carotenoid | 92 mg/L | Shake flask | [140] | |
| Rhodotorula glutinis | Whey | Carotenoid | 46 mg/L | Shake flask | [141] | |
| Rhodotorula rubra | Sugarcane juice | Carotenoid | 30.39 mg/g | Shake flask | [107] | |
| Rhodotorula rubra | Whey ultrafiltrate | Carotenoid | 12.1 mg/L | Shake flask | [142] | |
| Rhodotorula rubra | Whey sugar | Carotenoid | 0.705 OD/ml | Shake flask | [143] | |
| Rhodosporidium mucilaginosa | Potato extract | Carotenoid | 56 mg/L | Shake flask | [141] | |
| Rhodosporidium mucilaginosa | Coffee husk extract | Carotenoid | 21.35 mg/L | Shake flask | [144] | |
| Rhodosporidium mucilaginosa | Coffee pulp extract | Carotenoid | 16.36 mg/L | Shake flask | [144] | |
| Rhodosporidium mucilaginosa | Cassava bagasse | Carotenoid | 12.5 mg/L | Shake flask | [145] | |
| Rhodosporidium mucilaginosa | Onion peels and mung bean husk | Carotenoid | 719.69 μg/g | Shake flask | [146] | |
| Rhodosporidium toruloides | Waste extract | Carotenoid | 62 mg/L | Shake flask | [105] | |
| Rhodosporidium toruloides | Wheat straw hydrolysate | Carotenoid | 24.58 mg/L | Shake flask | [147] | |
| Rhodosporidium toruloides | Carob pulp syrup | Carotenoid | 9.79 μg/L | Shake flask | [148] | |
| Rhodotorula glutinis | Loquat kernel extract | Carotenoid | 62.73–72.36 mg/L | Shake flask | [66] | |
| Rhodotorula glutinis | Waste chicken feathers | Carotenoid | 6.47 mg/g | Shake flask | [140] | |
| Sporodiobolus pararoseus | Corn steep liquor and parboiled rice water | Carotenoid | 0.84 mg/L | Shake flask | [132] | |
| Sporodiobolus pararoseus | Sugarcane molasses and corn steep liquor | Carotenoid | 0.52 mg/L | Shake flask | [149] | |
| Sporidiobolus salmonicolor | Corn maceration and rice parboiling water | Carotenoid | 7.38 mg/L | 2 L Bioreactor | [150] | |
| Sporidiobolus salmonicolor | Cheese whey hydrolysate | Carotenoid | 590.4 μg/L | Shake flask | [151] | |
| Sporidiobolus pararoseus | Corn steep liquor and par- boiled rice water | Carotenoid | 843 μg/L | Shake flask | [132] | |
| Xanthophyllomyces dendrorhous | Sugar beet molasses | Astaxanthin | 40 mg/L | 100 L Bioreactor | [106] | |
| Xanthophyllomyces dendrorhous | Eucalyptus hydrolysate | Astaxanthin | 30.5 mg/L | 2 L Bioreactor | [152] | |
| Xanthophyllomyces dendrorhous | Mustard waste | Astaxanthin | 25.8 mg/L | Shake flask | [153] | |
| Xanthophyllomyces dendrorhous | Date juice | Astaxanthin | 23.8 mg/L | 3 L Bioreactor | [154] | |
| Xanthophyllomyces dendrorhous | Molasses | Astaxanthin | 15.3 mg/L | Shake flask | [155] | |
| Xanthophyllomyces dendrorhous | Grape juice | Astaxanthin | 9.8 μg/mL | Shake flask | [156] | |
| Xanthophyllomyces dendrorhous | Mesquite pods and corn steep liquor | Carotenoid | 293.41 ± 31.12 μg/g | Shake flask | [157] | |
| Microalgae | Haematococcus pluvialis | Primary-treated piggery wastewater | Astaxanthin | 83.9 mg/L | Shake flask | [79] |
| Phormidium autumnale | Slaughterhouse wastewater | Carotenoid | 107,902.5 kg/year | 2 L Bioreactor | [158] |
6. Submerged Culture Conditions for Pigment Production
| Pigment | Substrate | Temperature | pH | Reference |
|---|---|---|---|---|
| Carotenoid | Wheat straw hydrolysate | 30 °C | 5.3 | [147] |
| Coffee husk media | 28 °C | 5.7 | [144] | |
| Corn maceration and rice parboiling water | 25 °C | 4.0 | [150] | |
| Cassava bagasse | 25 °C | 6.0 | [145] | |
| Corn steep liquor and parboiled rice water | 25 °C | 4.0 | [132] | |
| Rice powder | 35 °C | 7.0 | [159] | |
| Mesquite pods and corn steep liquor | 20 °C | 5.5 | [157] | |
| Cheese whey | 26 °C | 7.3 | [125] | |
| Primary-treated piggery wastewater | 23 °C | 7.5 | [79] | |
| Slaughterhouse wastewater | 26 °C | 7.6 | [158] | |
| Flexirubin | Liquid pineapple waste | 30 °C | 7.0 | [113] |
| Melanin | Fruit pulp | 25 °C | 6.5 | [124] |
| Fruit waste | 30.7 °C | 6.8 | [108] | |
| Vegetable waste | 25 °C | 7.0 | [114] | |
| Monascus | Potato pomace | 28 °C | 5.0 | [64] |
| Glucose fermentation media | 30 °C | 5.5 | [103] | |
| Whey medium | 30 °C | 6.0 | [128] | |
| Grape waste | 30 °C | 6.5 | [46] | |
| Brewer’s spent grain media | 30 °C | 5.5–7.5 | [102] | |
| Rice powder | 32 °C | 3.5 | [160] | |
| Monascus-like | Potato dextrose broth | 24 °C | 6.4 | [161] |
| Orange peels | 24 °C | 5.0 | [131] | |
| Prodigiosin | Brown sugar | 25 °C | 7.0 | [119] |
| Cassava wastewater | 28 °C | 7.0 | [98] | |
| Peanut oil | 28 °C | - | [118] | |
| Peanut powder and olive oil | 26 °C | 7.0 | [120] | |
| Powdered peanut broth | 28 °C | 7.0 | [61] | |
| Peanut oil cake | 30 °C | 7.0 | [99] | |
| Wheat bran medium | 30 °C | - | [122] | |
| Pyocyanin | Cotton seed meal media | 37 °C | - | [115] |
| Riboflavin | Corn steep liquor | 37 °C | 7.2 | [109] |
| Corn steep liquor | 28 °C | 6.8 | [126] | |
| Violacein | Liquid pineapple waste | 30 °C | 7.0 | [111] |
| Sugarcane bagasse | 30 °C | 7.0 | [112] |
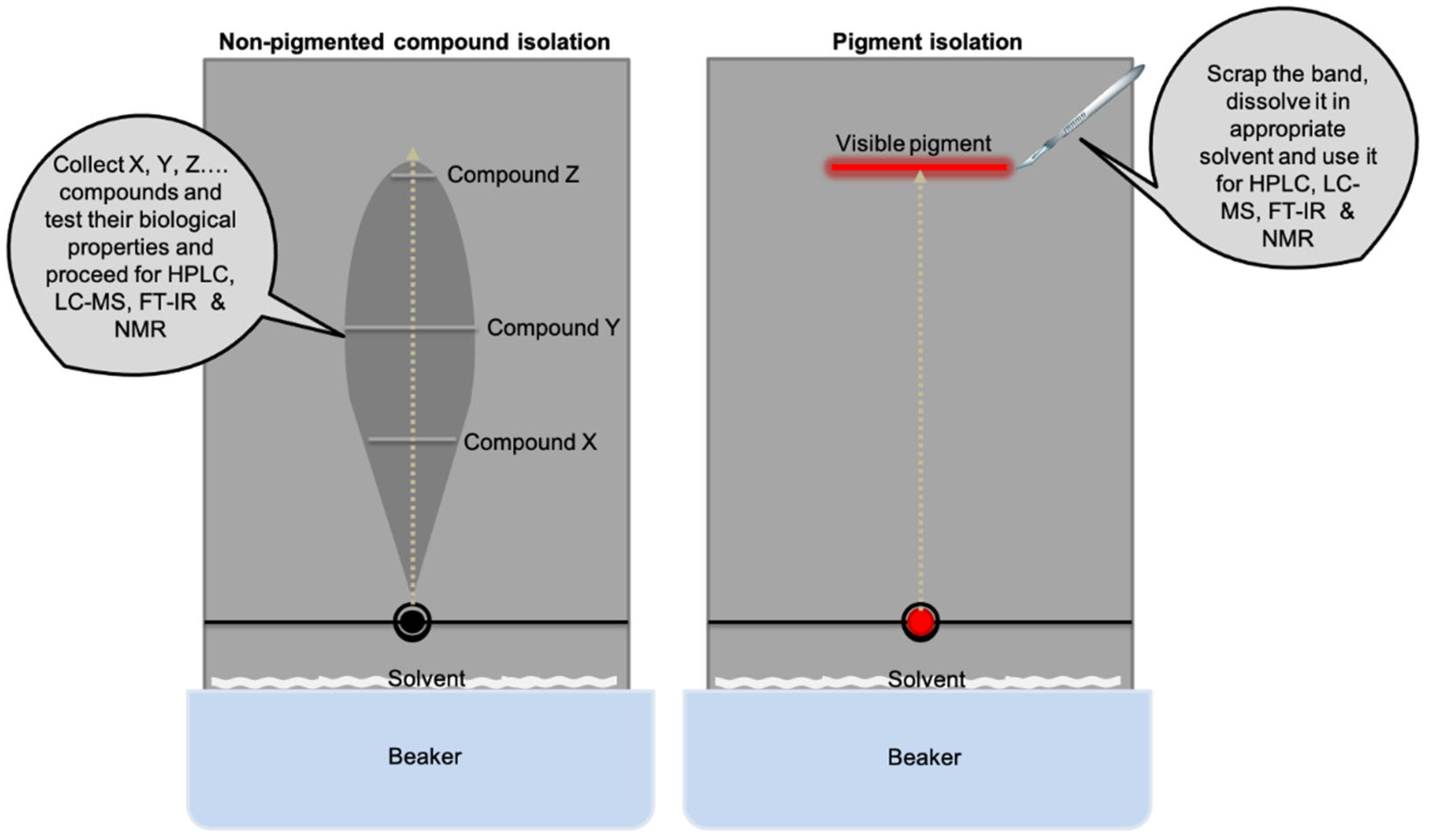
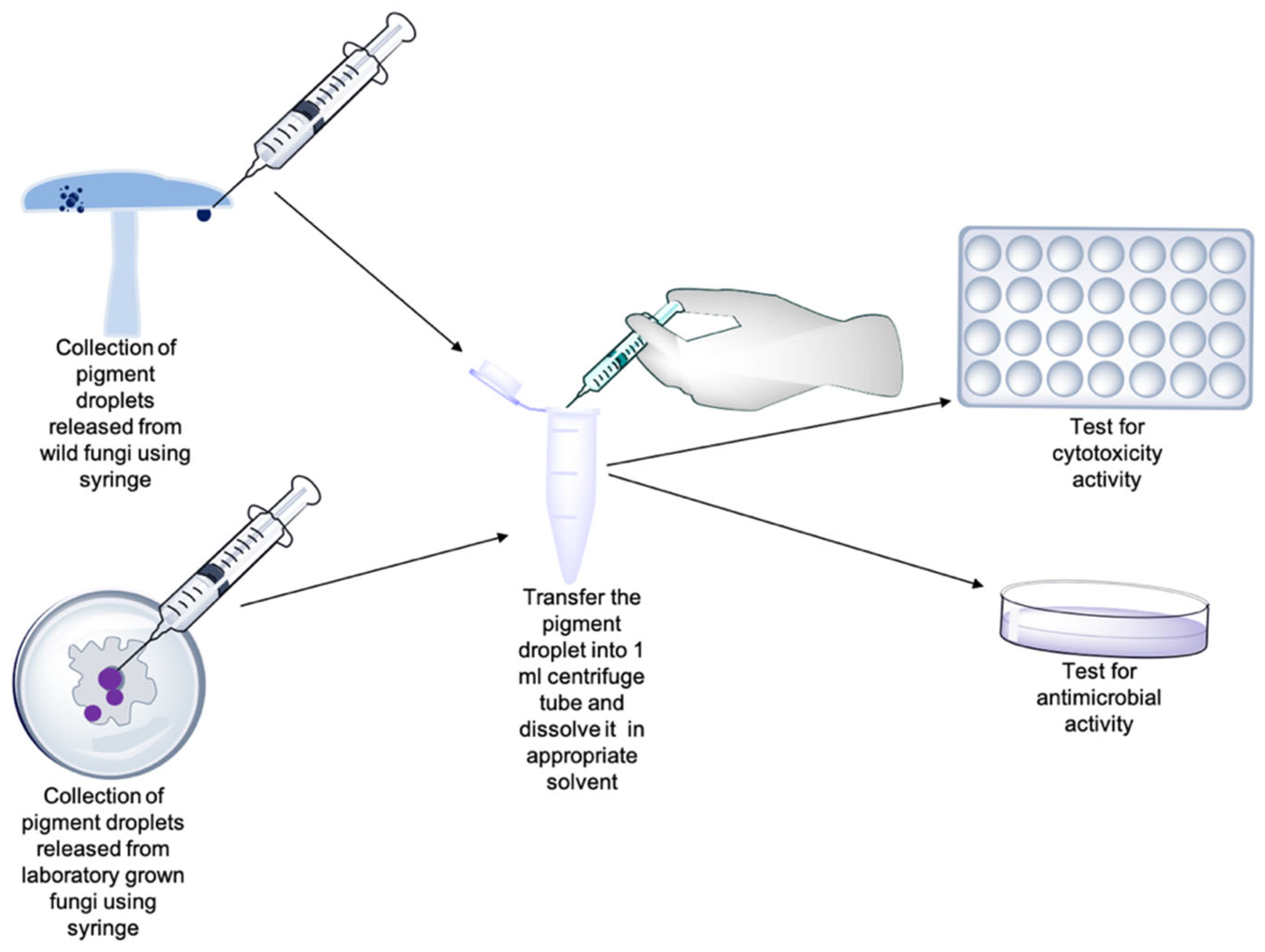
7. Rapid Identification of Microbial Pigments
8. Need for Targeted Drug Research on Microbial Pigments
9. Role of Pigmented Microbes in Climate Change
10. Current Applications of Microbial Pigments
11. Future Directions of Microbial Pigments
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramesh, C.; Vinithkumar, N.V.; Kirubagaran, R.; Venil, C.K.; Dufossé, L. Multifaceted applications of microbial pigments: Current knowledge, challenges and future directions for public health implications. Microorganisms 2019, 7, 186. [Google Scholar] [CrossRef]
- Venil, C.K.; Zakaria, Z.A.; Ahmad, W.A. Bacterial pigments and their applications. Process Biochem. 2013, 48, 1065–1079. [Google Scholar] [CrossRef]
- Sandmann, G. Carotenoids and their biosynthesis in fungi. Molecules 2022, 27, 1431. [Google Scholar] [CrossRef]
- Kalra, R.; Conlan, X.A.; Goel, M. Fungi as a potential source of pigments: Harnessing filamentous fungi. Front. Chem. 2020, 8, 369. [Google Scholar] [CrossRef]
- Lagashetti, A.C.; Dufossé, L.; Singh, S.K.; Singh, P.N. Fungal pigments and their prospects in different industries. Microorganisms 2019, 7, 604. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef]
- Mata-Gómez, L.C.; Montañez, J.C.; Méndez-Zavala, A.; Aguilar, C.N. Biotechnological production of carotenoids by yeasts: An overview. Microb. Cell Fact. 2014, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Kanamoto, H.; Nakamura, K.; Misawa, N. Carotenoid production in oleaginous yeasts. Adv. Exp. Med. Biol. 2021, 1261, 153–163. [Google Scholar]
- Chreptowicz, K.; Mierzejewska, J.; Tkácová, J.; Młynek, M.; Certik, M. Carotenoid-producing yeasts: Identification and characteristics of environmental isolates with a valuable extracellular enzymatic activity. Microorganisms 2019, 7, 653. [Google Scholar] [CrossRef]
- Igreja, W.S.; Maia, F.d.A.; Lopes, A.S.; Chisté, R.C. Biotechnological production of carotenoids using low cost-substrates is influenced by cultivation parameters: A review. Int. J. Mol. Sci. 2021, 22, 8819. [Google Scholar] [CrossRef]
- Lopes, G.; Clarinha, D.; Vasconcelos, V. Carotenoids from cyanobacteria: A biotechnological approach for the topical treatment of psoriasis. Microorganisms 2020, 8, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for high-value products towards human health and nutrition. Mar. Drugs 2019, 17, 304. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.K.; Murray, P. Exploitation of microalgae species for nutraceutical purposes: Cultivation aspects. Fermentation 2018, 4, 46. [Google Scholar] [CrossRef]
- Pailliè-Jiménez, M.E.; Stincone, P.; Brandelli, A. Natural pigments of microbial origin. Front. Sustain. Food Syst. 2020, 4, 590439. [Google Scholar] [CrossRef]
- Nemer, G.; Louka, N.; Vorobiev, E.; Salameh, D.; Nicaud, J.-M.; Maroun, R.G.; Koubaa, M. Mechanical cell disruption technologies for the extraction of dyes and pigments from microorganisms: A review. Fermentation 2021, 7, 36. [Google Scholar] [CrossRef]
- Lyakhovchenko, N.S.; Abashina, T.N.; Polivtseva, V.N.; Senchenkov, V.Y.; Pribylov, D.A.; Chepurina, A.A.; Nikishin, I.A.; Avakova, A.A.; Goyanov, M.A.; Gubina, E.D.; et al. A blue-purple pigment-producing bacterium isolated from the Vezelka river in the city of Belgorod. Microorganisms 2021, 9, 102. [Google Scholar] [CrossRef]
- Sen, T.; Barrow, C.J.; Deshmukh, S.K. Microbial pigments in the food industry—Challenges and the way forward. Front. Nutr. 2019, 6, 7. [Google Scholar] [CrossRef]
- Celedón, R.S.; Díaz, L.B. Natural pigments of bacterial origin and their possible biomedical applications. Microorganisms 2021, 9, 739. [Google Scholar] [CrossRef]
- Choi, S.Y.; Lim, S.; Yoon, K.; Lee, J.I.; Mitchell, R.J. Biotechnological activities and applications of bacterial pigments violacein and prodigiosin. J. Biol. Eng. 2021, 15, 10. [Google Scholar] [CrossRef]
- Han, R.; Xiang, R.; Li, J.; Wang, F.; Wang, C. High-level production of microbial prodigiosin: A review. J. Basic Microbiol. 2021, 61, 506–523. [Google Scholar] [CrossRef]
- Paul, T.; Bandyopadhyay, T.K.; Mondal, A.; Tiwari, O.N.; Muthuraj, M.; Bhunia, B. A comprehensive review on recent trends in production, purification, and applications of prodigiosin. Biomass Convers. Biorefinery 2022, 12, 1409–1431. [Google Scholar] [CrossRef]
- Durán, N.; Justo, G.Z.; Durán, M.; Brocchi, M.; Cordi, L.; Tasic, L.; Castro, G.R.; Nakazato, G. Advances in Chromobacterium violaceum and properties of violacein-its main secondary metabolite: A review. Biotechnol. Adv. 2016, 34, 1030–1045. [Google Scholar] [CrossRef] [PubMed]
- Foong, L.C.; Loh, C.W.L.; Ng, H.S.; Lan, J.C. Recent development in the production strategies of microbial carotenoids. World J. Microbiol. Biotechnol. 2021, 37, 12. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Cámara, S.; Ibañez, A.; Rubio, S.; Barreiro, C.; Barredo, J.-L. Main carotenoids produced by microorganisms. Encyclopedia 2021, 1, 1223–1245. [Google Scholar] [CrossRef]
- Nelis, H.J.; Leenheer, A.P. De Microbial sources of carotenoid pigments used in foods and feeds. J. Appl. Biotechnol. 1991, 70, 181–191. [Google Scholar] [CrossRef]
- Eman, M.M. Fungal and yeast carotenoids. J. Yeast Fungal Res. 2019, 10, 30–44. [Google Scholar] [CrossRef]
- Ávila-Román, J.; García-Gil, S.; Rodríguez-Luna, A.; Motilva, V.; Talero, E. Anti-inflammatory and anticancer effects of microalgal carotenoids. Mar. Drugs 2021, 19, 531. [Google Scholar] [CrossRef]
- Singh, S.; B.Nimse, S.; Mathew, D.E.; Dhimmar, A.; Sahastrabudhe, H.; Gajjar, A.; A.Ghadge, V.; Kumar, P.; B.Shinde, P. Microbial melanin: Recent advances in biosynthesis, extraction, characterization, and applications. Biotechnol. Adv. 2021, 53, 107773. [Google Scholar] [CrossRef]
- Tran-Ly, A.N.; Reyes, C.; Schwarze, F.W.M.R.; Ribera, J. Microbial production of melanin and its various applications. World J. Microbiol. Biotechnol. 2020, 36, 170. [Google Scholar] [CrossRef]
- Vendruscolo, F.; Bühler, R.M.M.; de Carvalho, J.C.; de Oliveira, D.; Moritz, D.E.; Schmidell, W.; Ninow, J.L. Monascus: A reality on the production and application of microbial pigments. Appl. Biochem. Biotechnol. 2016, 178, 211–223. [Google Scholar] [CrossRef]
- Chaudhary, V.; Katyal, P.; Puniya, A.K.; Panwar, H. Natural pigment from Monascus: The production and therapeutic significance. J. Appl. Microbiol. 2021, 133, 18–38. [Google Scholar] [CrossRef] [PubMed]
- de Morais, M.G.; Prates, D.d.F.; Moreira, J.B.; Duarte, J.H.; Costa, J.A.V. Phycocyanin from microalgae: Properties, extraction and purification, with some recent applications. Ind. Biotechnol. 2018, 14, 30–37. [Google Scholar] [CrossRef]
- Sonani, R.R. Recent advances in production, purification and applications of phycobiliproteins. World J. Biol. Chem. 2016, 7, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Puzorjov, A.; McCormick, A.J. Phycobiliproteins from extreme environments and their potential applications. J. Exp. Bot. 2020, 71, 3827–3842. [Google Scholar] [CrossRef]
- Averianova, L.A.; Balabanova, L.A.; Son, O.M.; Podvolotskaya, A.B.; Tekutyeva, L.A. Production of vitamin B2 (riboflavin) by microorganisms: An overview. Front. Bioeng. Biotechnol. 2020, 8, 570828. [Google Scholar] [CrossRef]
- Sharma, M.; Usmani, Z.; Gupta, V.K.; Bhat, R. Valorization of fruits and vegetable wastes and by-products to produce natural pigments. Crit. Rev. Microbiol. 2021, 41, 535–563. [Google Scholar] [CrossRef]
- Pargai, D.; Jahan, S.; Gahlot, M. Functional properties of natural dyed textiles. In Chemistry and Technology of Natural and Synthetic Dyes and Pigments; Samanta, A.K., Awwad, N., Algarni, H.M., Eds.; IntechOpen: London, UK, 2020; pp. 1–19. [Google Scholar]
- Newsome, A.G.; Culver, C.A.; van Breemen, R.B. Nature’s palette: The search for natural blue colorants. J. Agric. Food Chem. 2014, 62, 6498–6511. [Google Scholar] [CrossRef]
- Yadav, A.K.; Jain, C.K.; Malik, D.S. Toxic characterization of textile dyes and effluents in relation to human health hazards. J. Sustain. Environ. Res. 2014, 3, 95–102. [Google Scholar]
- Babitha, S. Microbial pigments. In Biotechnology for Agro-Industrial Residues Utilisation: Utilisation of Agro-Residues; Nigam, P.S., Pandey, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 147–162. ISBN 9781402099410. [Google Scholar]
- Dey, S.; Nagababu, B.H. Applications of food color and bio-preservatives in the food and its effect on the human health. Food Chem. Adv. 2022, 1, 100019. [Google Scholar] [CrossRef]
- Slama, H.B.; Bouket, A.C.; Pourhassan, Z.; Alenezi, F.N.; Silini, A.; Cherif-Silini, H.; Oszako, T.; Luptakova, L.; Golinska, P.; Belbahri, L. Diversity of synthetic dyes from textile industries, discharge impacts and treatment methods. Appl. Sci. 2021, 11, 6255. [Google Scholar] [CrossRef]
- Dufossé, L. Current and potential natural pigments from microorganisms (bacteria, yeasts, fungi, microalgae). In Handbook on Natural Pigments in Food and Beverages Industrial Applications for Omproving Food Color; Carle, R., Schweiggert, R.M., Eds.; Woodhead Publishing: Sawston, UK, 2016; pp. 337–354. [Google Scholar]
- Venil, C.K.; Dufossé, L.; Devi, P.R. Bacterial pigments: Sustainable compounds with market potential for pharma and food industry. Front. Sustain. Food Syst. 2020, 4, 100. [Google Scholar] [CrossRef]
- Ramesh, C.H.; Dufossé, L. Ecological and biotechnological aspects of pigmented Microbes: A way forward in development of food and pharmaceutical grade pigments. Microorganisms 2021, 9, 637. [Google Scholar] [CrossRef]
- Lopes, F.C.; Tichota, D.M.; Pereira, J.Q.; Segalin, J.; Rios, A.d.O.; Brandelli, A. Pigment production by filamentous fungi on agro-industrial byproducts: An eco-friendly alternative. Appl. Biochem. Biotechnol. 2013, 171, 616–625. [Google Scholar] [CrossRef] [PubMed]
- De Medeiros, T.D.M.; Dufossé, L.; Bicas, J.L. Lignocellulosic substrates as starting materials for the production of bioactive biopigments. Food Chem. X. 2022, 13, 100223. [Google Scholar] [CrossRef]
- Lopes, F.C.; Ligabue-Braun, R. Agro-industrial residues: Eco-friendly and inexpensive substrates for microbial pigments production. Front. Sustain. Food Syst. 2021, 5, 589414. [Google Scholar] [CrossRef]
- Sajjad, W.; Din, G.; Rafiq, M.; Iqbal, A.; Khan, S.; Zada, S.; Ali, B.; Kang, S. Pigment production by cold-adapted bacteria and fungi: Colorful tale of cryosphere with wide range applications. Extremophiles 2020, 24, 447–473. [Google Scholar] [CrossRef]
- Nawaz, A.; Chaudhary, R.; Shah, Z.; Dufossé, L.; Fouillaud, M.; Mukhtar, H.; ul Haq, I. An overview on industrial and medical applications of bio-pigments synthesized by marine bacteria. Microorganisms 2021, 9, 11. [Google Scholar] [CrossRef]
- Ramesh, C.; Vinithkumar, N.V.; Kirubagaran, R. Marine pigmented bacteria: A prospective source of antibacterial compounds. J. Nat. Sci. Biol. Med. 2019, 10, 104–113. [Google Scholar] [CrossRef]
- Liu, M.; Peng, F.; Wang, Y.; Zhang, K.; Chen, G.; Fang, C. Kineococcus xinjiangensis sp. nov., isolated from desert sand. Int. J. Syst. Evol. Microbiol. 2009, 59, 1090–1093. [Google Scholar] [CrossRef]
- Mumtaz, R.; Bashir, S.; Numan, M.; Shinwari, Z.K.; Ali, M. Pigments from soil bacteria and their therapeutic properties: A mini review. Curr. Microbiol. 2019, 76, 783–790. [Google Scholar] [CrossRef]
- Daudu, R.; Parker, C.W.; Singh, N.K.; Wood, J.M.; Debieu, M.; O’Hara, N.B.; Mason, C.E.; Venkateswaran, K. Draft genome sequences of Rhodotorula mucilaginosa strains isolated from the international space station. Microbiol. Resour. Announc. 2020, 9, e00570-20. [Google Scholar] [CrossRef] [PubMed]
- Lebeau, J.; Venkatachalam, M.; Fouillaud, M.; Petit, T.; Vinale, F.; Dufossé, L.; Caro, Y. Production and new extraction method of polyketide red pigments produced by ascomycetous fungi from terrestrial and marine habitats. J. Fungi 2017, 3, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatachalam, M.; Magalon, H.; Dufossé, L.; Fouillau, M. Production of pigments from the tropical marine-derived fungi Talaromyces albobiverticillius: New resources for natural red-colored metabolites. J. Food Compos. Anal. 2018, 70, 35–48. [Google Scholar] [CrossRef]
- Majumdar, S.; Priyadarshinee, R.; Kumar, A.; Mandal, T.; Mandal, D.D. Exploring Planococcus sp. TRC1, a bacterial isolate, for carotenoid pigment production and detoxification of paper mill effluent in immobilized fluidized bed reactor. J. Clean. Prod. 2019, 211, 1389–1402. [Google Scholar] [CrossRef]
- Padhan, B.; Poddar, K.; Sarkar, D.; Sarkar, A. Production, purification, and process optimization of intracellular pigment from novel psychrotolerant Paenibacillus sp. BPW19. Biotechnol. Rep. 2021, 29, e00592. [Google Scholar] [CrossRef]
- Khaneja, R.; Perez-Fons, L.; Fakhry, S.; Baccigalupi, L.; Steiger, S.; To, E.; Sandmann, G.; Dong, T.C.; Ricca, E.; Fraser, P.D.; et al. Carotenoids found in Bacillus. J. Appl. Microbiol. 2010, 108, 1889–1902. [Google Scholar] [CrossRef]
- Hayashi, M.; Ishibashi, T.; Kuwahara, D.; Hirasawa, K. Commercial production of astaxanthin with Paracoccus carotinifaciens. Adv. Exp. Med. Biol. 2021, 1261, 11–20. [Google Scholar]
- Giri, A.V.; Anandkumar, N.; Muthukumaran, G.; Pennathur, G. A novel medium for the enhanced cell growth and production of prodigiosin from Serratia marcescens isolated from soil. BMC Microbiol. 2004, 4, 11. [Google Scholar] [CrossRef]
- Wibowo, J.T.; Kellermann, M.Y.; Petersen, L.-E.; Alfiansah, Y.R.; Lattyak, C.; Schupp, P.J. Characterization of an insoluble and soluble form of melanin produced by Streptomyces cavourensis SV 21, a sea cucumber associated bacterium. Mar. Drugs 2022, 20, 54. [Google Scholar] [CrossRef]
- Zhu, Y.; Shang, X.; Yang, L.; Zheng, S.; Liu, K.; Lia, X. Purification, identification and properties of a new blue pigment produced from Streptomyces sp. A1013Y. Food Chem. 2020, 308, 125600. [Google Scholar] [CrossRef]
- Chen, X.; Gui, R.; Li, N.; Wu, Y.; Chen, J.; Wu, X.; Qin, Z.; Yang, S.-T.; Li, X. Production of soluble dietary fibers and red pigments from potato pomace in submerged fermentation by Monascus purpureus. Process Biochem. 2021, 111, 159–166. [Google Scholar] [CrossRef]
- Darwesh, O.M.; Matter, I.A.; Almoallim, H.S.; Alharbi, S.A.; Oh, Y.-K. Isolation and optimization of Monascus ruber OMNRC45 for red pigment production and evaluation of the pigment as a food colorant. Appl. Sci. 2020, 10, 8867. [Google Scholar] [CrossRef]
- Taskin, M.; Erdal, S. Production of carotenoids by Rhodotorula glutinis MT-5 in submerged fermentation using the extract from waste loquat kernels as substrate. J. Sci. Food Agric. 2011, 91, 1440–1445. [Google Scholar] [CrossRef] [PubMed]
- Rekha, R.; Nimsi, K.A.; Manjusha, K.; Sirajudheen, T.K. Marine yeast Rhodotorula paludigena VA 242 a pigment enhancing feed additive for the ornamental fish koi carp. Aquac. Fish. 2022. [Google Scholar] [CrossRef]
- Libkind, D.; Moliné, M.; Tognetti, C. Isolation and selection of new astaxanthin producing strains of Xanthophyllomyces dendrorhous. In Microbial Carotenoids from Fungi: Methods and Protocols, Methods in Molecular Biology; Barredo, J.-L., Ed.; Springer Science+Business Media: New York, NY, USA, 2012; Volume 898, pp. 183–194. [Google Scholar]
- Libkind, D.; Moliné, M.; Colabella, F. Isolation and selection of new astaxanthin-producing strains of Phaffia rhodozyma. In Microbial Carotenoids from Fungi: Methods and Protocols, Methods in Molecular Biology; Barreiro, C., Barredo, J.-L., Eds.; Humana Press: New York, NY, USA, 2018; Volume 1852, pp. 297–310. [Google Scholar]
- Ruiz-Domínguez, M.C.; Jáuregui, M.; Medina, E.; Jaime, C.; Cerezal, P. Rapid green extractions of C-phycocyanin from Arthrospira maxima for functional applications. Appl. Sci. 2019, 9, 1987. [Google Scholar] [CrossRef]
- Zheng, Y.; Mo, L.; Zhang, W.; Duan, Y.; Huang, J.; Chen, C.; Gao, Y.; Shi, X.; Li, F.; Yang, J.; et al. Phycocyanin fluorescent probe from Arthrospira platensis: Preparation and application in LED-CCD fluorescence density strip qualitative detection system. J. Appl. Phycol. 2019, 31, 1107–1115. [Google Scholar] [CrossRef]
- Kissoudi, M.; Sarakatsianos, I.; Samanidou, V. Isolation and purification of food-grade C-phycocyanin from Arthrospira platensis and its determination in confectionery by HPLC with diode array detection. J. Sep. Sci. 2018, 41, 975–981. [Google Scholar] [CrossRef]
- Leema, J.T.M.; Kirubagaran, R.; Vinithkumar, N.V.; Dheenan, P.S.; Karthikayulu, S. High value pigment production from Arthrospira (Spirulina) platensis cultured in seawater. Bioresour. Technol. 2010, 101, 9221–9227. [Google Scholar] [CrossRef]
- Ru, I.T.K.; Sung, Y.Y.; Jusoh, M.; Wahid, M.E.A.; Nagappan, T. Chlorella vulgaris: A perspective on its potential for combining high biomass with high value bioproducts. Appl. Phycol. 2020, 1, 2–11. [Google Scholar] [CrossRef]
- Rahman, D.Y.; Sarian, F.D.; van Wijk, A.; Martinez-Garcia, M.; van der Maarel, M.J.E.C. Thermostable phycocyanin from the red microalga Cyanidioschyzon merolae, a new natural blue food colorant. J. Appl. Phycol. 2017, 29, 1233–1239. [Google Scholar] [CrossRef]
- Eisele, L.E.; Bakhru, S.H.; Liu, X.; MacColl, R.; Edwards, M.R. Studies on C-phycocyanin from Cyanidium caldarium, a eukaryote at the extremes of habitat. Biochim. Biophys. Acta. 2000, 1456, 99–107. [Google Scholar] [CrossRef]
- Baudelet, P.-H.; Gagez, A.-L.; Bérard, J.-B.; Juin, C.; Bridiau, N.; Kaas, R.; Thiéry, V.; Cadoret, J.-P.; Picot, L. Antiproliferative activity of Cyanophora paradoxa pigments in melanoma, breast and lung cancer cells. Mar. Drugs 2013, 11, 4390–4406. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, L.; Hantke, A.; Eriksen, N.T. Purification of the photosynthetic pigment C-phycocyanin from heterotrophic Galdieria sulphuraria. J. Sci. Food Agric. Food Agric. 2013, 93, 2933–2938. [Google Scholar] [CrossRef]
- Kang, C.D.; An, J.Y.; Park, T.H.; Sim, S.J. Astaxanthin biosynthesis from simultaneous N and P uptake by the green alga Haematococcus pluvialis in primary-treated wastewater. Biochem. Eng. J. 2006, 31, 234–238. [Google Scholar] [CrossRef]
- Francezon, N.; Herbaut, M.; Bardeau, J.-F.; Cougnon, C.; Bélanger, W.; Tremblay, R.; Jacquette, B.; Dittmer, J.; Pouvreau, J.-B.; Mouget, J.-L.; et al. Electrochromic properties and electrochemical behavior of marennine, a bioactive blue-green pigment produced by the marine diatom Haslea ostrearia. Mar. Drugs 2021, 19, 231. [Google Scholar] [CrossRef] [PubMed]
- Gabed, N.; Verret, F.; Peticca, A.; Kryvoruchko, I.; Gastineau, R.; Bosson, O.; Séveno, J.; Davidovich, O.; Davidovich, N.; Witkowski, A.; et al. What was old is new again: The pennate diatom Haslea ostrearia (Gaillon) Simonsen in the multi-omic age. Mar. Drugs 2022, 20, 234. [Google Scholar] [CrossRef]
- Lee, A.H.; Shin, H.Y.; Park, J.H.; Koo, S.Y.; Kim, S.M.; Yang, S.H. Fucoxanthin from microalgae Phaeodactylum tricornutum inhibits pro-inflammatory cytokines by regulating both NF-κB and NLRP3 inflammasome activation. Sci. Rep. 2021, 11, 543. [Google Scholar] [CrossRef]
- Udayan, A.; Pandey, A.K.; Sirohi, R.; Sreekumar, N.; Sang, B.-I.; Sim, S.J.; Kim, S.H.; Pandey, A. Production of microalgae with high lipid content and their potential as sources of nutraceuticals. Phytochem. Rev. 2022, 23, 1–28. [Google Scholar] [CrossRef]
- Margalith, P.Z. Pigment Microbiology; Chapman & Hall: London, UK, 1992. [Google Scholar]
- Williamson, N.R.; Fineran, P.C.; Leeper, F.J.; Salmond, G.P.C. The biosynthesis and regulation of bacterial prodiginines. Nat. Rev. Microbiol. 2006, 4, 887–899. [Google Scholar] [CrossRef]
- Barredo, J.L.; García-Estrada, C.; Kosalkova, K.; Barreiro, C. Biosynthesis of astaxanthin as a main carotenoid in the heterobasidiomycetous yeast Xanthophyllomyces dendrorhous. J. Fungi 2017, 3, 44. [Google Scholar] [CrossRef]
- Ren, Y.; Sun, H.; Deng, J.; Huang, J.; Chen, F. Carotenoid production from microalgae: Biosynthesis, salinity responses and novel biotechnologies. Mar. Drugs 2021, 19, 713. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Jing, X.; Liu, X.; Lindblad, P. Biotechnological production of the sunscreen pigment scytonemin in cyanobacteria: Progress and strategy. Mar. Drugs 2021, 19, 129. [Google Scholar] [CrossRef] [PubMed]
- Misawa, N.; Nakagawa, M.; Kobayashi, K.; Yamano, S.; Izawa, Y.; Nakamura, K.; Harashima, K. Elucidation of the Erwinia uredovora carotenoid biosynthetic pathway by functional analysis of gene products expressed in Escherichia coli. J. Bacteriol. 1990, 172, 6704–6712. [Google Scholar] [CrossRef] [Green Version]
- Sankari, M.; Rao, P.R.; Hemachandran, H.; Pullela, P.K.; George, P.D.C.; Tayubi, I.A.; Subramanian, B.; Gothandam, K.M.; Singh, P.; Ramamoorthy, S. Prospects and progress in the production of valuable carotenoids: Insights from metabolic engineering, synthetic biology, and computational approaches. J. Biotechnol. 2018, 266, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Liu, N.; Greisen, P.; Li, J.; Qiao, K.; Huang, S.; Stephanopoulos, G. Removal of lycopene substrate inhibition enables high carotenoid productivity in Yarrowia lipolytica. Nat. Commun. 2022, 13, 572. [Google Scholar] [CrossRef]
- Liu, A.; Chen, A.J.; Liu, B.; Wei, Q.; Bai, J.; Hu, Y. Investigation of citrinin and monacolin K gene clusters variation among pigment producer Monascus species. Fungal Genet. Biol. 2022, 160, 103687. [Google Scholar] [CrossRef]
- Xu, X.; Chu, X.; Du, B.; Huang, C.; Xie, C.; Zhang, Z.; Jiang, L. Functional characterization of a novel violacein biosynthesis operon from Janthinobacterium sp. B9-8. Appl. Microbiol. Biotechnol. 2022, 106, 2903–2916. [Google Scholar] [CrossRef]
- Sunil, L.; Shetty, N.P. Biosynthesis and regulation of anthocyanin pathway genes. Appl. Microbiol. Biotechnol. 2022, 106, 1783–1798. [Google Scholar] [CrossRef]
- Zha, J.; Koffas, M.A.G. Production of anthocyanins in metabolically engineered microorganisms: Current status and perspectives. Synth. Syst. Biotechnol. 2017, 2, 259–266. [Google Scholar] [CrossRef]
- Sánchez-Muñoz, S.; Mariano-Silva, G.; Leite, M.O.; Mura, F.B.; Verma, M.L.; Silva, S.S.d.; Chandel, A.K. Production of fungal and bacterial pigments and their applications. In Biotechnological Production of Bioactive Compounds; Verma, M.L., Chandel, A.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 327–361. [Google Scholar]
- Soccol, C.R.; da Costa, E.S.F.; Letti, L.A.J.; Karp, S.G.; Woiciechowski, A.L.; Vandenberghe, L.P.d.S. Recent developments and innovations in solid state fermentation. Biotechnol. Res. Innov. 2017, 1, 52–71. [Google Scholar] [CrossRef]
- de Araújo, H.W.C.; Fukushima, K.; Takaki, G.M.C. Prodigiosin production by Serratia marcescens UCP 1549 using renewable-resources as a low cost substrate. Molecules 2010, 15, 6931–6940. [Google Scholar] [CrossRef] [PubMed]
- Naik, C.; Srisevita, J.M.; Shushma, K.N.; Farah, N.; Shilpa, A.C.; Muttanna, C.D.; Darshan, N.; Sannadurgappa, D. Peanut oil cake: A novel substrate for enhanced cell growth and prodigiosin production from Serratia marcescens CF-53. J. Res. Biol. 2012, 2, 549–557. [Google Scholar]
- Zhou, Z.; Yin, Z. Corncob hydrolysate, an efficient substrate for Monascus pigment production through submerged fermentation. Biotechnol. Appl. Biochem. 2014, 61, 716–723. [Google Scholar] [CrossRef]
- Haque, M.A.; Kachrimanidou, V.; Koutinas, A.; Lin, C.S.K. Valorization of bakery waste for biocolorant and enzyme production by Monascus purpureus. J. Biotechnol. 2016, 231, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Silbir, S.; Goksungur, Y. Natural red pigment production by Monascus purpureus in submerged fermentation systems using a food industry waste: Brewer’s spent grain. Foods 2019, 8, 161. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Luo, Y.; Guo, T.; Tang, C.; Chai, X.; Zhao, W.; Bai, J.; Lin, Q. Cost-effective pigment production by Monascus purpureus using rice straw hydrolysate as substrate in submerged fermentation. J. Biosci. Bioeng. 2020, 129, 229–236. [Google Scholar] [CrossRef]
- Hilares, R.T.; de Souza, R.A.; Marcelino, P.F.; da Silva, S.S.; Dragone, G.; Mussatto, S.I.; Santos, J.C. Sugarcane bagasse hydrolysate as a potential feedstock for red pigment production by Monascus ruber. Food Chem. 2018, 245, 786–791. [Google Scholar] [CrossRef]
- Sinha, S.; Singh, G.; Arora, A.; Paul, D. Carotenoid production by red yeast isolates grown in agricultural and “mandi” waste. Waste Biomass Valorization 2021, 12, 3939–3949. [Google Scholar] [CrossRef]
- An, G.H.; Jang, B.G.; Cho, M.H. Cultivation of the carotenoid- hyperproducing mutant 2A2N of the red yeast Xanthophyllomyces dendrorhous (Phaffia rhodozyma) with molasses. J. Biosci. Bioeng. 2001, 92, 121–125. [Google Scholar] [CrossRef]
- Bonadio, M.d.P.; de Freita, L.A.; Mutton, M.J.R. Carotenoid production in sugarcane juice and synthetic media supplemented with nutrients by Rhodotorula rubra l02. Braz. J. Microbiol. 2018, 49, 872–878. [Google Scholar] [CrossRef]
- Tarangini, K.; Mishra, S. Production of melanin by soil microbial isolate on fruit waste extract: Two step optimization of key paramete. Biotechnol. Rep. 2014, 4, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Park, Y.H.; Han, J.K.; Park, J.H.; Lee, K.H.; Kyung, H.; Choi, H. Microorganism for Producing Rboflavin and Method for Producing Riboflavin using the Same 2004. U.S. Patent 2004O110248A1, 23 January 2007. [Google Scholar]
- Cassarini, M.; Crônier, D.; Besaury, L.; Rémond, C. Protein-rich agro-industrial co-products are key substrates for growth of Chromobacterium vaccinii and its violacein bioproduction. Waste Biomass Valorization 2022, 1–10. [Google Scholar] [CrossRef]
- Aruldass, C.A.; Rubiyatno; Venil, C.K.; Ahmad, W.A. Violet pigment production from liquid pineapple waste by Chromobacterium violaceum UTM5 and evaluation of its bioactivity. RSC Adv. 2015, 5, 51524–51536. [Google Scholar] [CrossRef]
- Ahmad, W.A.; Yusof, N.Z.; Nordin, N.; Zakaria, Z.A.; Rezali, M.F. Production and characterization of violacein by locally isolated Chromobacterium violaceum grown in agricultural wastes. Appl. Biochem. Biotechnol. 2012, 167, 1220–1234. [Google Scholar] [CrossRef]
- Aruldass, C.A.; Dufosse, L.; Ahmad, W.A. Current perspective of yellowish-orange pigments from microorganisms-a review. J. Clean. Prod. 2018, 180, 168–182. [Google Scholar] [CrossRef]
- Tarangini, K.; Mishra, S. Production, characterization and analysis of melanin from isolated marine Pseudomonas sp. using vegetable waste. Res. J. Eng. Sci. 2013, 2, 40–46. [Google Scholar]
- El-Fouly, M.Z.; Sharaf, A.M.; Shahin, A.A.M.; El-Bialy, H.A.; Omara, A.M.A. Biosynthesis of pyocyanin pigment by Pseudomonas aeruginosa. J. Radiat. Res. Appl. Sci. 2015, 8, 36–48. [Google Scholar] [CrossRef]
- Joshi, V.K.; Attri, D.; Rana, M.S. Optimization of apple pomace based medium and fermentation conditions for pigment production by Sarcina sp. Indian J. Nat. Prod. Resour. 2011, 2, 421–427. [Google Scholar]
- Sumathi, C.; Mohanapriya, D.; Swarnalatha, S.; Dinesh, M.G.; Sekaran, G. Production of prodigiosin using tannery fleshing and evaluating its pharmacological effects. Sci. World J. 2014, 2014, 290–327. [Google Scholar] [CrossRef]
- Hernandez-Velasco, P.; Morales-Atilano, I.; Rodríguez-Delgado, M.; Rodríguez-Delgado, J.M.; Luna-Moreno, D.; Avalos-Alanís, F.G.; Villarreal-Chiu, J.F. Photoelectric evaluation of dye-sensitized solar cells based on prodigiosin pigment derived from Serratia marcescens 11E. Dye. Pigment. 2020, 177, 108278. [Google Scholar] [CrossRef]
- Aruldass, C.A.; Venil, C.K.; Zakaria, Z.A.; Ahmad, W.A. Brown sugar as a low-cost medium for the production of prodigiosin by locally isolated Serratia marcescens UTM1. Int. Biodeterior. Biodegrad. 2014, 95, 19–24. [Google Scholar] [CrossRef]
- Lin, C.; Jia, X.; Chen, L.; Zhang, H.; Lin, R.; Chen, J. Enhanced production of prodigiosin by Serratia marcescens FZSF02 in the form of pigment pellets. Electron J. Biotechnol. 2019, 40, 58–64. [Google Scholar] [CrossRef]
- Picha, P.; Kale, D.; Dave, I.; Pardeshi, S. Comparative studies on prodigiosin production by Serratia marcescens using various crude fatty acid sources its characterization and applications. J. Int. J. Curr. Microbiol. Appl. Sci. 2015, 2, 254–267. [Google Scholar]
- Luti, K.J.K.; Yonis, R.W.; Mahmoud, S.T. An application of solid state fermentation and elicitation with some microbial cells for the enhancement of prodigiosin production by Serratia marcescens. J. Al-Nahrain Univ. 2018, 21, 98–105. [Google Scholar] [CrossRef]
- El-Bondkly, A.M.A.; El-Gendy, M.M.A.; Bassyouni, R.H. Overproduction and biological activity of prodigiosin-like pigments from recombinant fusant of endophytic marine Streptomyces species. Antonie Van Leeuwenhoek 2012, 102, 719–734. [Google Scholar] [CrossRef]
- Arikan, E.B.; Canli, O.; Caro, Y.; Dufossé, L.; Dizge, N. Production of bio-based pigments from food processing industry by- products (apple, pomegranate, black carrot, red beet pulps) using Aspergillus carbonarius. J. Fungi 2020, 6, 240. [Google Scholar] [CrossRef]
- Roukas, T.; Varzakakou, M.; Kotzekidou, P. From cheese whey to carotenes by Blakeslea trispora in a bubble column reactor. Appl. Biochem. Biotechnol. 2015, 175, 182–193. [Google Scholar] [CrossRef]
- Park, E.Y.; Ito, Y.; Nariyama, M.; Sugimoto, T.; Lies, D.; Kato, T. The improvement of riboflavin production in Ashbya gossypii via disparity mutagenesis and DNA microarray analysis. Appl. Microbiol. Biotechnol. 2011, 91, 1315–1326. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, W.; Nkechi, O.; Lu, P.; Bai, J.; Lin, Q.; Liu, J. Utilization of low-cost agricultural by-product rice husk for Monascus pigments production via submerged batch-fermentation. J. Sci. Food Agric. 2022, 102, 2454–2463. [Google Scholar] [CrossRef]
- Mehri, D.; Perendeci, N.A.; Goksungur, Y. Utilization of whey for red pigment production by Monascus purpureus in submerged fermentation. Fermentation 2021, 7, 75. [Google Scholar] [CrossRef]
- Embaby, A.M.; Hussein, M.N.; Hussein, A. Monascus orange and red pigments production by Monascus purpureus ATCC16436 through co-solid state fermentation of corn cob and glycerol: An eco-friendly environmental low cost approach. PLoS ONE 2018, 13, e0207755. [Google Scholar] [CrossRef] [PubMed]
- Silveira, S.T.; Daroit, D.J.; Brandelli, A. Pigment production by Monascus purpureus in grape waste using factorial design. Food Sci. Technol. 2008, 41, 170–174. [Google Scholar] [CrossRef]
- Kantifedaki, A.; Kachrimanidou, V.; Mallouchos, A.; Papanikolaou, S.; Koutinas, A.A. Orange processing waste valorisation for the production of bio-based pigments using the fungal strains Monascus purpureus and Penicillium purpurogenum. J. Clean. Prod. 2018, 185, 882–890. [Google Scholar] [CrossRef]
- Valduga, E.; Ribeiro, A.H.R.; Cence, K.; Colet, R.; Tiggemann, L.; Zeni, J.; Toniazzo, G. Carotenoids production from a newly isolated Sporidiobolus pararoseus strain using agroindustrial substrates. Biocatal. Agric. Biotechnol. 2014, 3, 207–213. [Google Scholar] [CrossRef]
- Morales-Oyervides, L.; Ruiz-Sánchez, J.P.; Oliveira, J.C.; Sousa-Gallagher, M.J.; Morales-Martínez, T.K.; Albergamo, A.; Salvo, A.; Giuffrida, D.; Dufossé, L.; Montañez, J. Medium design from corncob hydrolyzate for pigment production by Talaromyces atroroseus GH2: Kinetics modeling and pigments characterization. Biochem. Eng. J. 2020, 161, 107698. [Google Scholar] [CrossRef]
- Pandit, S.G.; Ramesh, K.P.M.; Puttananjaiah, M.H.; Dhale, M.A. Cicer arietinum (Bengal gram) husk as alternative for Talaromyces purpureogenus CFRM02 pigment production: Bioactivities and identification. LWT—Food Sci. Technol. 2019, 116, 108499. [Google Scholar] [CrossRef]
- Nasrabadi, M.; Razavi, S. Optimization of β-carotene production by a mutant of the lactose-positive yeast Rhodotorula achenorium from whey ultrafiltrate. Food Sci. Biotech. 2011, 20, 445–454. [Google Scholar] [CrossRef]
- Schneider, T.; Graeff-Hönninger, S.; French, W.T.; Hernandez, R.; Merkt, N.; Claupein, W.; Hetrick, M.; Pham, P. Lipid and carotenoid production by oleaginous red yeast Rhodotorula glutinis cultivated on brewery effluents. Energy 2013, 61, 34–43. [Google Scholar] [CrossRef]
- Tinoi, J.; Rakariyatham, N.; Deming, R.L. Simplex optimization of carotenoid production by Rhodotorula glutinis using hydrolyzed mung bean waste flour as substrate. Process Biochem. 2005, 40, 2551–2557. [Google Scholar] [CrossRef]
- Ribeiro, J.E.S.; Sant’Ana, A.M.d.S.; Martini, M.; Sorce, C.; Andreucci, A.; de Melo, D.J.N.; da Silva, F.L.H. Rhodotorula glutinis cultivation on cassava wastewater for carotenoids and fatty acids generation. Biocatal. Agric. Biotechnol. 2019, 22, 101419. [Google Scholar] [CrossRef]
- Saenge, C.; Cheirsilp, B.; Suksaroge, T.T.; Bourtoom, T. Potential use of oleaginous red yeast Rhodotorula glutinis for the bioconversion of crude glycerol from biodiesel plant to lipids and carotenoids. Process Biochem. 2011, 46, 210–218. [Google Scholar] [CrossRef]
- Taskin, M.; Sisman, T.; Erdal, S.; Kurbanoglu, E.B. Use of waste chicken feathers as peptone for production of carotenoids in submerged culture of Rhodotorula glutinis MT-5. Eur. Food Res. Technol. 2011, 233, 657–665. [Google Scholar] [CrossRef]
- Marova, I.; Carnecka, M.; Halienova, A.; Certik, M.; Dvorakova, T.; Haronikova, A. Use of several waste substrates for carotenoid-rich yeast biomass production. J. Environ. Manag. 2011, 95, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Frengova, G.I.; Emilina, S.D.; Beshkova, D.M. Carotenoid production by lactoso-negative yeasts co-cultivated with lactic acid bacteria in whey ultrafiltrate. Z. Naturforsch. C. J. Biosci. 2003, 58, 562–567. [Google Scholar] [CrossRef]
- Kaur, B.; Chakraborty, D.; Kaur, H. Production and stability analysis of yellowish pink pigments from Rhodotorula rubra MTCC 1446. Int. J. Microbiol. 2012, 7, 1–7. [Google Scholar] [CrossRef]
- Moreira, M.D.; Melo, M.M.; Coimbra, J.M.; Dos Reis, K.C.; Schwan, R.F.; Silva, C.F. Solid coffee waste as alternative to produce carotenoids with antioxidant and antimicrobial activities. Waste Manag. 2018, 82, 93–99. [Google Scholar] [CrossRef]
- Manimala, M.R.A.; Murugesan, R. Studies on carotenoid pigment production by yeast Rhodotorula mucilaginosa using cheap materials of agro-industrial origin. Pharma Innov. 2017, 6, 80. [Google Scholar]
- Sharma, R.; Ghoshal, G. Optimization of carotenoids production by Rhodotorula mucilaginosa (MTCC-1403) using agro-industrial waste in bioreactor: A statistical approach. Biotechnol. Rep. 2020, 25, e00407. [Google Scholar] [CrossRef]
- Liu, Z.; Feist, A.M.; Dragone, G.; Mussatto, S.I. Lipid and carotenoid production from wheat straw hydrolysates by different oleaginous yeasts. J. Clean. Prod. 2020, 249, 119308. [Google Scholar] [CrossRef]
- Freitas, C.; Parreira, T.M.; Roseiro, J.; Reis, A.; da Silva, T.L. Selecting low-cost carbon sources for carotenoid and lipid production by the pink yeast Rhodosporidium toruloides NCYC 921 using flow cytometry. Bioresour. Technol. 2014, 158, 355–359. [Google Scholar] [CrossRef]
- Machado, W.R.C.; de Medeiros Burkert, J.F. Optimization of agroindustrial medium for the production of carotenoids by wild yeast Sporidiobolus pararoseus. Afr. J. Microbiol. Res. 2015, 9, 209–219. [Google Scholar]
- Colet, R.; Urnau, L.; Bampi, J.; Zeni, J.; Dias, B.B.; Rodrigues, E.; Jacques, R.A.; Di Luccio, M.; Valduga, E. Use of low-cost agro products as substrate in semi-continuous process to obtain carotenoids by Sporidiobolus salmonicolor. Biocatal. Agric. Biotechnol. 2017, 11, 268–274. [Google Scholar] [CrossRef]
- Valduga, E.; Tatsch, P.; Vanzo, L.; Rauber, F.; Di Luccio, M.; Treichel, H. Assessment of hydrolysis of cheese whey and use of hydrolysate for bioproduction of carotenoids by Sporidiobolus salmonicolor CBS 2636. J. Sci. Food Agric. 2009, 89, 1060–1065. [Google Scholar] [CrossRef]
- Vázquez, M.; Santos, V.; Parajó, J.C. Fed-batch cultures of Phaffia rhodozyma in xylose-containing media made from wood hydrolysates. Food Biotechnol. 1998, 12, 43–55. [Google Scholar] [CrossRef]
- Tinoi, J.; Rakariyatham, N.; Deming, R.L. Utilization of mustard waste isolates for improved production of astaxanthin by Xanthophyllomyces dendrorhous. J. Ind. Microbiol. Biotechnol. 2006, 33, 309–314. [Google Scholar] [CrossRef]
- Ramírez, J.; Obledo, N.; Arellano, M.; Herrera, E. Astaxanthin production by Phaffia rhodozyma in a fed-batch culture using a low cost medium feedin. e-Gnosis 2006, 4, 1–9. [Google Scholar]
- Haard, N.F. Astaxanthin formation by the yeast Phaffia rhodozyma on molasses. Biotechnol. Lett. 1988, 10, 609–614. [Google Scholar] [CrossRef]
- Meyer, P.S.; du Preez, J.C. Astaxanthin production by a Phaffia rhodozyma mutant on grape juice. World J. Microbiol. Biotechnol. 1994, 10, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Villegas-Méndez, M.Á.; Aguilar-Machado, D.E.; Balagurusamy, N.; Montañez, J.; Morales-Oyervides, L. Agro-industrial wastes for the synthesis of carotenoids by Xanthophyllomyces dendrorhous: Mesquite pods- based medium design and optimization. Biochem. Eng. J. 2019, 150, 107260. [Google Scholar] [CrossRef]
- Rodrigues, D.B.; Flores, É.M.M.; Barin, J.S.; Mercadante, A.Z.; Jacob-Lopes, E.; Zepka, L.Q. Production of carotenoids from microalgae cultivated using agroindustrial wastes. Food Res. Int. 2014, 65, 144–148. [Google Scholar] [CrossRef]
- Korumilli, T.; Mishra, S. Carotenoid production by Bacillus clausii using rice powder as the sole substrate pigment analyses and optimization of key pro- duction parameters. J. Biochem. Technol. 2014, 5, 788–794. [Google Scholar]
- Lian, X.; Wang, C.; Guo, K. Identification of new red pigments produced by Monascus ruber. Dye. Pigment. 2007, 73, 121–125. [Google Scholar] [CrossRef]
- Venkatachalam, M.; Shum-Chéong-Sing, A.; Dufossé, L.; Fouillaud, M. Statistical optimization of the physico-chemical parameters for pigment production in submerged fermentation of Talaromyces albobiverticillius 30548. Microorganisms 2020, 8, 711. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, C.; Vinithkumar, N.V.; Kirubagaran, R.; Venil, C.K.; Dufosse, L. Applications of prodigiosin extracted from marine red pigmented bacteria Zooshikella sp. and actinomycete Streptomyces sp. Microorganisms 2020, 8, 556. [Google Scholar] [CrossRef] [PubMed]
- Orlandi, V.T.; Martegani, E.; Giaroni, C.; Baj, A.; Bolognese, F. Bacterial pigments: A colorful palette reservoir for biotechnological applications. Biotechnol. Appl. Biochem. 2021, 69, 981–1001. [Google Scholar] [CrossRef]
- Prasastha, V.R.; Yasur, J.; Abishad, P.; Unni, V.; Gourkhede, D.P.; Nishanth, M.A.D.; Niveditha, P.; Vergis, J.; Malik, S.V.S.; Byrappa, K.; et al. Antimicrobial efficacy of green synthesized nanosilver with entrapped cinnamaldehyde against multi-drug-resistant enteroaggregative Escherichia coli in Galleria mellonella. Pharmaceutics 2022, 14, 1924. [Google Scholar] [CrossRef]
- Pereira, M.F.; Rossi, C.C. Overview of rearing and testing conditions and a guide for optimizing Galleria mellonella breeding and use in the laboratory for scientific purposes. J. Pathol. Microbiol. Immunol. 2020, 128, 607–620. [Google Scholar] [CrossRef]
- Pereira, M.F.; Rossi, C.C.; da Silva, G.C.; Rosa, J.N.; Bazzolli, D.M.S. Galleria mellonella as an infection model: An in-depth look at why it works and practical considerations for successful application. Pathog. Dis. 2020, 78, ftaa056. [Google Scholar] [CrossRef]
- Tsai, C.J.-Y.; Loh, J.M.S.; Proft, T. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef]
- Andrea, A.; Krogfelt, K.A.; Jenssen, H. Methods and challenges of using the greater wax moth (Galleria mellonella) as a model organism in antimicrobial compound discovery. Microorganisms 2019, 7, 85. [Google Scholar] [CrossRef]
- Cutuli, M.A.; Petronio Petronio, G.; Vergalito, F.; Magnifico, I.; Pietrangelo, L.; Venditti, N.; Di Marco, R. Galleria mellonella as a consolidated in vivo model hosts: New developments in antibacterial strategies and novel drug testing. Virulence 2019, 10, 527–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojda, I. Immunity of the greater wax moth Galleria mellonella. Insect Sci. 2017, 24, 342–357. [Google Scholar] [CrossRef]
- Abas, N.; Kalair, A.R.; Khan, N.; Haider, A.; Saleem, Z.; Saleem, M.S. Natural and synthetic refrigerants, global warming: A review. Renew. Sustain. Energy Rev. 2018, 90, 557–569. [Google Scholar] [CrossRef]
- Khan, S.; Malik, A. Toxicity evaluation of textile effluents and role of native soil bacterium in biodegradation of a textile dye. Environ. Sci. Pollut. Res. 2018, 25, 4446–4458. [Google Scholar] [CrossRef]
- Six, J.; Frey, S.D.; Thiet, R.K.; Batten, K.M. Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Sci. Soc. Am. J. 2006, 70, 555–569. [Google Scholar] [CrossRef]
- Tanet, L.; Martini, S.; Casalot, L.; Tamburini, C. Reviews and syntheses: Bacterial bioluminescence -ecology and impact in the biological carbon pump. Biogeosciences Discuss. 2020, 17, 3757–3778. [Google Scholar] [CrossRef]
- Jiao, N.; Robinson, C.; Azam, F.; Thomas, H.; Baltar, F.; Dang, H.; Hardman-Mountford, N.J.; Johnson, M.; Kirchman, D.L.; Koch, B.P.; et al. Mechanisms of microbial carbon sequestration in the ocean—Future research directions. Biogeosciences 2014, 11, 5285–5306. [Google Scholar] [CrossRef]
- Zhang, C.; Dang, H.; Azam, F.; Benner, R.; Legendre, L.; Passow, U.; Polimene, L.; Robinson, C.; Suttle, C.A.; Jiao, N. Evolving paradigms in biological carbon cycling in the ocean. Natl. Sci. Rev. 2018, 5, 481–499. [Google Scholar] [CrossRef]
- Matteoli, F.P.; Passarelli-Araujo, H.; Reis, R.J.A.; da Rocha, L.O.; de Souza, E.M.; Aravind, L.; Olivares, F.L.; Venancio, T.M. Genome sequencing and assessment of plant growth-promoting properties of a Serratia marcescens strain isolated from vermicompost. BMC Genom. 2018, 19, 750. [Google Scholar] [CrossRef]
- Habash, S.S.; Brass, H.U.C.; Klein, A.S.; Klebl, D.P.; Weber, T.M.; Classen, T.; Pietruszka, J.; Grundler, F.M.W.; Schleker, A.S.S. Novel prodiginine derivatives demonstrate bioactivities on plants, nematodes, and fungi. Front. Plant Sci. 2020, 11, 579807. [Google Scholar] [CrossRef]
- Gutiérrez-Román, M.I.; Holguín-Meléndez, F.; Bello-Mendoza, R.; Guillén-Navarro, K.; Dunn, M.F.; Huerta-Palacios, G. Production of prodigiosin and chitinases by tropical Serratia marcescens strains with potential to control plant pathogens. World J. Microbiol. Biotechnol. 2012, 28, 145–153. [Google Scholar] [CrossRef]
- Suryawanshi, R.K.; Patil, C.D.; Borase, H.P.; Narkhede, C.P.; Salunke, B.K.; Patil, S.V. Mosquito larvicidal and pupaecidal potential of prodigiosin from Serratia marcescens and understanding its mechanism of action. Pestic. Biochem. Physiol. 2015, 123, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Banik, A.; Pandya, P.; Patel, B.; Rathod, C.; Dangar, M. Characterization of halotolerant, pigmented, plant growth promoting bacteria of groundnut rhizosphere and its in-vitro evaluation of plant-microbe protocooperation to withstand salinity and metal stress. Sci. Total Environ. 2018, 630, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Jimtha, C.J.; Jishma, P.; Sreelekha, S.; Chithra, S.; Radhakrishnan, E.K. Antifungal properties of prodigiosin producing rhizospheric Serratia sp. Rhizosphere 2017, 3, 105–108. [Google Scholar] [CrossRef]
- Meena, B.; Anburajan, L.; Sathish, T.; Das, A.K.; Vinithkumar, N.V.; Kirubagaran, R.; Dharani, G. Studies on diversity of Vibrio sp. and the prevalence of hapA, tcpI, st, rtxA&C, acfB, hlyA, ctxA, ompU and toxR genes in environmental strains of Vibrio cholerae from Port Blair bays of South Andaman, India. Mar. Pollut. Bull. 2019, 144, 105–116. [Google Scholar]
- Sivalingam, K.M. Isolation, identification and evaluation of Spirulina platensis for its effect on seed germination of groundnut (Arachis hypogaea L.), Wolaita Sodo, Southern Ethiopia. J. Algal Biomass Utln. 2020, 11, 34–42. [Google Scholar]
- Oyanedel, R.; Hinsley, A.; Dentinger, B.T.M.; Milner-Gulland, E.J.; Furci, G. A way forward for wild fungi in international sustainability policy. Conserv. Lett. 2022, 15, e12882. [Google Scholar] [CrossRef]
- Rillig, M.C.; Mummey, D.L. Mycorrhizas and soil structure. New Phytol. 2006, 171, 41–53. [Google Scholar] [CrossRef]
- Chahal, K.; Gupta, V.; Verma, N.K.; Chaurasia, A.; Rana, B. Arbuscular mycorrhizal (AM) fungi as a tool for sustainable agricultural system. In Mycorrhizal Fungi-Utilization in Agriculture and Forestry; Radhakrishnan, R., Ed.; IntechOpen: London, UK, 2020; pp. 1–12. [Google Scholar]
- Kiers, E.T.; Duhamel, M.; Beesetty, Y.; Mensah, J.A.; Franken, O.; Verbruggen, E.; Fellbaum, C.R.; Kowalchuk, G.A.; Hart, M.M.; Bago, A.; et al. Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis. Science 2011, 333, 880–882. [Google Scholar] [CrossRef]
- Liang, T.; Shi, X.; Guo, T.; Peng, S. Arbuscular mycorrhizal fungus mediate changes in mycorrhizosphere soil aggregates. Agric. Sci. 2015, 6, 1455–1463. [Google Scholar] [CrossRef]
- Mohammadi, K.; Khalesro, S.; Sohrabi, Y.; Heidari, G. A review: Beneficial effects of the mycorrhizal fungi for plant growth. J. Appl. Environ. Biol. Sci. 2011, 1, 310–319. [Google Scholar]
- French, K.E. Engineering mycorrhizal symbioses to alter plant metabolism and improve crop health. Front Microbiol. 2017, 8, 1403. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.; Khare, A.; Rawat, P. Glomalin arbuscular mycorrhizal fungal reproduction, lifestyle and dynamic role in global sustainable agriculture for future generation. In Fungal Reproduction and Growth; Sultan, S., Singh, G.K.S., Eds.; IntechOpen: London, UK, 2022; pp. 1–22. [Google Scholar]
- Hause, B.; Fester, T. Molecular and cell biology of arbuscular mycorrhizal symbiosis. Planta 2005, 221, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, B.D.; Kyaschenko, J.; Varenius, K.; Clemmensen, K.E.; Dahlberg, A.; Karltun, E.; Stendahl, J. A group of ectomycorrhizal fungi restricts organic matter accumulation in boreal forest. Ecol. Lett. 2021, 24, 1341–1351. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z.-L.; Santalahti, M.; Köster, K.; Berninger, F.; Pumpanen, J.; Heinonsalo, J.; Sun, H. Soil fungal community structure in boreal pine forests: From southern to subarctic areas of Finland. Front. Microbiol. 2021, 12, 653896. [Google Scholar] [CrossRef] [PubMed]
- Clemmensen, K.E.; Finlay, R.D.; Dahlberg, A.; Stenlid, J.; Wardle, D.A.; Lindahl, B.D. Carbon sequestration is related to mycorrhizal fungal community shifts during long-term succession in boreal forests. New Phytol. 2015, 205, 1525–1536. [Google Scholar] [CrossRef]
- Lombardi, N.; Caira, S.; Troise, A.D.; Scaloni, A.; Vitaglione, P.; Vinale, F.; Marra, R.; Salzano, A.M.; Lorito, M.; Woo, S.L. Trichoderma applications on strawberry plants modulate the physiological processes positively affecting fruit production and quality. Front. Microbiol. 2020, 11, 1364. [Google Scholar] [CrossRef]
- Gill, S.S.; Gill, R.; Trivedi, D.K.; Anjum, N.A.; Sharma, K.K.; Ansari, M.W.; Ansari, A.A.; Johri, A.K.; Prasad, R.; Pereira, E.; et al. Piriformospora indica: Potential and significance in plant stress tolerance. Front. Microbiol. 2016, 7, 332. [Google Scholar] [CrossRef]
- Fernandez, C.W.; Koide, R.T. The function of melanin in the ectomycorrhizal fungus Cenococcum geophilum under water stress. Fungal Ecol. 2013, 6, 479–486. [Google Scholar] [CrossRef]
- Kapoor, D.; Karnwal, A. Yeast as plant growth promoter and biocontrol agent. In Fungi Bio-Prospects in Sustainable Agriculture, Environment and Nano-Technology Volume 1: Fungal Diversity of Sustainable Agriculture; Sharma, V.K., Shah, M., Parmar, S., Kumar, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 429–457. [Google Scholar]
- Sondak, C.F.; Ang, P.O.; Beardall, J.; Bellgrove, A.; Boo, S.M.; Gerung, G.S.; Hepburn, C.D.; Hong, D.D.; Hu, Z.; Kawai, H.; et al. Carbon dioxide mitigation potential of seaweed aquaculturebeds (SABs). J. Appl. Phycol. 2017, 29, 2363–2373. [Google Scholar] [CrossRef]
- Zhao, B.; Su, A. Macro assessment of microalgae-based CO2 sequestration: Environmental and energy effects. Algal Res. 2020, 51, 102066. [Google Scholar] [CrossRef]
- Oncel, S.S.; Kose, A.; Oncel, D.S. Carbon sequestration in microalgae photobioreactors building integrated. In Start-Up Creation The Smart Eco-Efficient Built Environment; Pacheco-Torgal, F., Rasmussen, E., Granqvist, C.-G., Ivanov, V., Kaklauskas, A., Makonin, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 161–200. [Google Scholar]
- Vale, M.A.; Ferreira, A.; Pires, J.C.M.; Gonçalves, A.L. CO2 capture using microalgae. In Advances in Carbon Capture Methods, Technologies and Applications; Rahimpour, M.R., Farsi, M., Makarem, M.A., Eds.; Elsevier: Duxford, UK, 2020; pp. 381–405. [Google Scholar]
- Pavlik, D.; Zhong, Y.; Daiek, C.; Liao, W.; Morgan, R.; Clary, W.; Liu, Y. Microalgae cultivation for carbon dioxide sequestration and protein production using a high-efficiency photobioreactor system. Algal Res. 2017, 25, 413–420. [Google Scholar] [CrossRef]
- Xu, X.; Gu, X.; Wang, Z.; Shatner, W.; Wang, Z. Progress, challenges and solutions of research on photosynthetic carbon sequestration efficiency of microalgae. Renew. Sustain. Energy Rev. 2019, 110, 65–82. [Google Scholar] [CrossRef]
- Narala, R.R.; Garg, S.; Sharma, K.K.; Thomas-Hall, S.R.; Deme, M.; Li, Y.; Schenk, P.M. Comparison of microalgae cultivation in photobioreactor, open raceway pond, and a two-stage hybrid system. Front. Energy Res. 2016, 4, 29. [Google Scholar] [CrossRef] [Green Version]
- Onyeaka, H.; Miri, T.; Obileke, K.; Hart, A.; Anumudu, C.; Al-Sharify, Z.T. Minimizing carbon footprint via microalgae as a biological capture. Carbon Capture Sci. Technol. 2021, 1, 100007. [Google Scholar] [CrossRef]
- Jerney, J.; Spilling, K. Large Scale Cultivation of Microalgae: Open and Closed Systems. Methods Mol. Biol. 2020, 1980, 1–8. [Google Scholar]
- Viswanaathan, S.; Perumal, P.K.; Sundaram, S. Integrated approach for carbon sequestration and wastewater treatment using algal–bacterial consortia: Opportunities and challenges. Sustainability 2022, 14, 1075. [Google Scholar] [CrossRef]
- Vishnupriya, S.; Bhavaniramya, S.; Baskaran, D.; Karthiayani, A. Microbial pigments and their application. In Microbial Polymers; Vaishnav, A., Choudhary, D.K., Eds.; Springer Nature: Singapore, 2021; pp. 197–214. [Google Scholar]
- Numan, M.; Bashir, S.; Mumtaz, R.; Tayyab, S.; Rehman, N.U.; Khan, A.L.; Shinwari, Z.K.; Al-Harrasi, A. Therapeutic applications of bacterial pigments: A review of current status and future opportunities. 3 Biotech 2018, 8, 207. [Google Scholar] [CrossRef]
- Mohammadi, M.A.; Ahangari, H.; Mousazadeh, S.; Hosseini, S.M.; Dufossé, L. Microbial pigments as an alternative to synthetic dyes and food additives: A brief review of recent studies. Bioprocess Biosyst. Eng. 2022, 45, 1–12. [Google Scholar] [CrossRef]
- Kaur, P.; Singh, S.; Ghoshal, G.; Ramamurthy, P.C.; Parihar, P.; Singh, J.; Singh, A. Valorization of agri-food industry waste for the production of microbial pigments: An eco-friendly approach. In Advances in Agricultural and Industrial Microbiology; Nayak, S.K., Baliyarsingh, B., Mannazzu, I., Singh, A., Mishra, B.B., Eds.; Springer: Singapore, 2022; pp. 137–166. [Google Scholar]
- Ramesh, C.H.; Anwesh, M.; Vinithkumar, N.V.; Kirubagaran, R.; Dufossé, L. Complete genome analysis of undecylprodigiosin pigment biosynthesizing marine Streptomyces species displaying potential bioactive applications. Microorganisms 2021, 9, 2249. [Google Scholar] [CrossRef]
- Amorim, L.F.A.; Mouro, C.; Riool, M.; Gouveia, I.C. Antimicrobial food packaging based on prodigiosin-incorporated double-layered bacterial cellulose and chitosan composites. Polymers 2022, 14, 315. [Google Scholar] [CrossRef] [PubMed]
- Metwally, R.A.; El Sikaily, A.; El-Sersy, N.A.; A.Ghozlan, H.; Sabry, S.A. Antimicrobial activity of textile fabrics dyed with prodigiosin pigment extracted from marine Serratia rubidaea RAM_Alex bacteria. Egypt. J. Aquat. Res. 2021, 47, 301–305. [Google Scholar] [CrossRef]
- Lee, J.; Kim, J.; Song, J.E.; Song, W.-S.; Kim, E.-J.; Kim, Y.-G.; Jeong, H.-J.; Kim, H.R.; Choi, K.-Y.; Kim, B.-G. Production of tyrian purple indigoid dye from tryptophan in Escherichia coli. Nat. Chem. Biol. 2021, 17, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.-Y. A review of recent progress in the synthesis of bio-indigoids and their biologically assisted end-use applications. Dye. Pigment. 2020, 181, 108570. [Google Scholar] [CrossRef]
- Hou, Y.; Yan, M.; Wang, Q.; Wang, Y.; Xu, Y.; Wang, Y.; Li, H.; Wang, H. C-phycocyanin from Spirulina maxima as a green fluorescent probe for the highly selective detection of Mercury(II) in seafood. Food Anal. Methods 2017, 10, 1931–1939. [Google Scholar] [CrossRef]
- Patel, A.K.; Albarico, F.P.J.B.; KrishnaPerumal, P.; Vadrale, A.P.; Nian, C.T.; Chau, H.T.B.; Anwar, C.; Wani, H.M.; Pal, A.; Saini, R.; et al. Algae as an emerging source of bioactive pigments. Bioresour. Technol. 2022, 351, 126910. [Google Scholar] [CrossRef]
- Srivastava, A.; Kalwani, M.; Chakdar, H.; Pabbi, S.; Shukla, P. Biosynthesis and biotechnological interventions for commercial production of microalgal pigments: A review. Bioresour. Technol. 2022, 352, 127071. [Google Scholar] [CrossRef]



| Pigmented Microbe | Pigment | Source | Application | Reference |
|---|---|---|---|---|
| Bacteria | ||||
| Bacillus | Carotenoid | Different sources | Colorant | [59] |
| Chromobacterium violaceum | Violacein | River water and agricultural waste | Antimicrobial | [22] |
| Janthinobacterium sp. | Violacein | River water | Antimicrobial | [16] |
| Paracoccus carotinifaciens | Astaxanthin | Soil | Coloring agent | [60] |
| Planococcus sp. | Carotenoid | Wastewater | Food additive | [57] |
| Serratia marcescens | Prodigiosin | Soil | Dye, antimicrobial | [61] |
| Streptomyces cavourensis | Melanin | Sea cucumber | Antimicrobial | [62] |
| Streptomyces sp. | 4,8,13-trihydroxy-6,11-dione-trihydrogranaticins A (TDTA) | Soil | Feed additive | [63] |
| Fungi | ||||
| Monascus purpureus | Monascus red pigment | Red mold rice | Food colorant | [64] |
| Monascus ruber | Monascus red pigment | Soil | Food colorant | [65] |
| Talaromyces albobiverticillius | Monascus-like | Marine | Industrial | [56] |
| Yeast | ||||
| Rhodotorula glutinis | Carotenoid | Soil | Food colorant | [66] |
| Rhodotorula paludigena | Carotenoid | Mangrove | Fish feed | [67] |
| Xanthophyllomyces dendrorhous (=Phaffia rhodozyma) | Astaxanthin | Trees | Food additive | [68,69] |
| Cyanobacteria | ||||
| Arthrospira maxima (=Spirulina maxima) | Phycocyanin | Freshwater | Food and drug | [70] |
| Arthrospira platensis (=Spirulina platensis) | Phycocyanin | Freshwater | Dye and food additive, fluorescent probe | [71,72] |
| Arthrospira platensis | Phycocyanin | Seawater | Food and drug | [73] |
| Microalgae | ||||
| Chlorella vulgaris | Carotenoids | Freshwater | Food and drug | [74] |
| Cyanidioschyzon merolae | Phycocyanin | Hot sulfuric springs and geysers | Food colorant | [75] |
| Cyanidium caldarium | Phycocyanin | Thermal area | Food colorant | [76] |
| Cyanophora paradoxa, Dunaliella salina | Zeaxanthin and β-cryptoxanthin | Freshwater | Anticancer | [77] |
| Galdieria sulphuraria | Phycocyanin | Hot and acidic springs | Food and drug | [78] |
| Haematococcus pluvialis | Astaxanthin | Freshwater | Feed additive | [79] |
| Haslea ostrearia | Marennine | Seawater | Food colorant and drug | [80,81] |
| Phaeodactylum tricornutum | Fucoxanthin | Marine | Anti-inflammatory | [82] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramesh, C.; Prasastha, V.R.; Venkatachalam, M.; Dufossé, L. Natural Substrates and Culture Conditions to Produce Pigments from Potential Microbes in Submerged Fermentation. Fermentation 2022, 8, 460. https://doi.org/10.3390/fermentation8090460
Ramesh C, Prasastha VR, Venkatachalam M, Dufossé L. Natural Substrates and Culture Conditions to Produce Pigments from Potential Microbes in Submerged Fermentation. Fermentation. 2022; 8(9):460. https://doi.org/10.3390/fermentation8090460
Chicago/Turabian StyleRamesh, Chatragadda, V. R. Prasastha, Mekala Venkatachalam, and Laurent Dufossé. 2022. "Natural Substrates and Culture Conditions to Produce Pigments from Potential Microbes in Submerged Fermentation" Fermentation 8, no. 9: 460. https://doi.org/10.3390/fermentation8090460
APA StyleRamesh, C., Prasastha, V. R., Venkatachalam, M., & Dufossé, L. (2022). Natural Substrates and Culture Conditions to Produce Pigments from Potential Microbes in Submerged Fermentation. Fermentation, 8(9), 460. https://doi.org/10.3390/fermentation8090460