

Selection of Yarrowia lipolytica Strains as Possible Solution to Valorize Untreated Cheese Whey
 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Cultivation
2.2. Cheese Whey
2.3. Proteolysis Profile
2.4. Lipolysis Profile
2.5. Cheese Whey Characterization
2.6. Screening Y. lipolytica Growth in Cheese Whey
2.7. Volatile Molecule Profile
2.8. Data Analyses
3. Results
3.1. Enzymatic Properties of the 20 Strains of Y. lipolytica
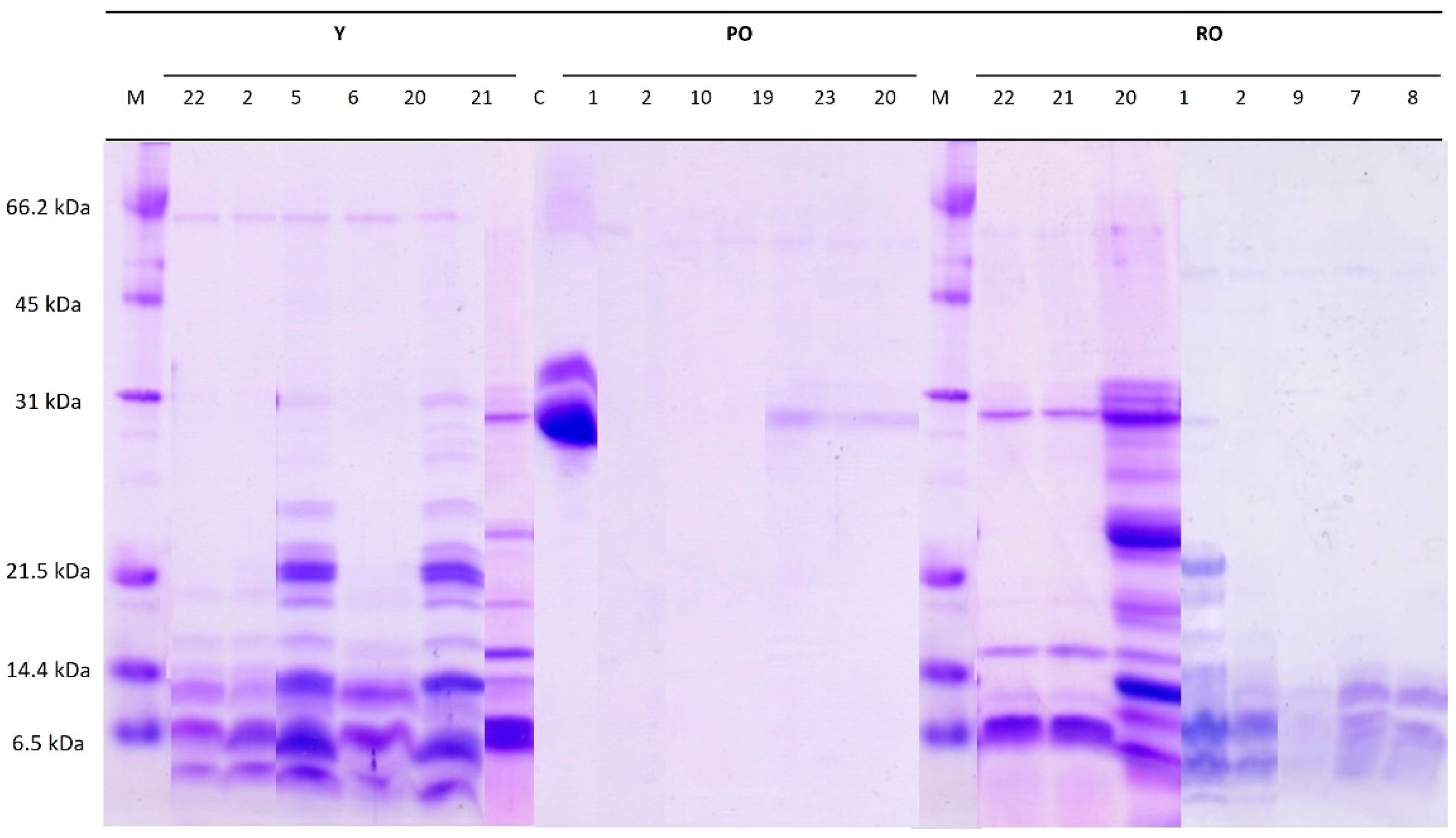
3.1.1. Proteolysis Profile
3.1.2. Lipolysis Profile
3.2. Growth in Cheese Whey
3.2.1. Cheese Whey Characterization
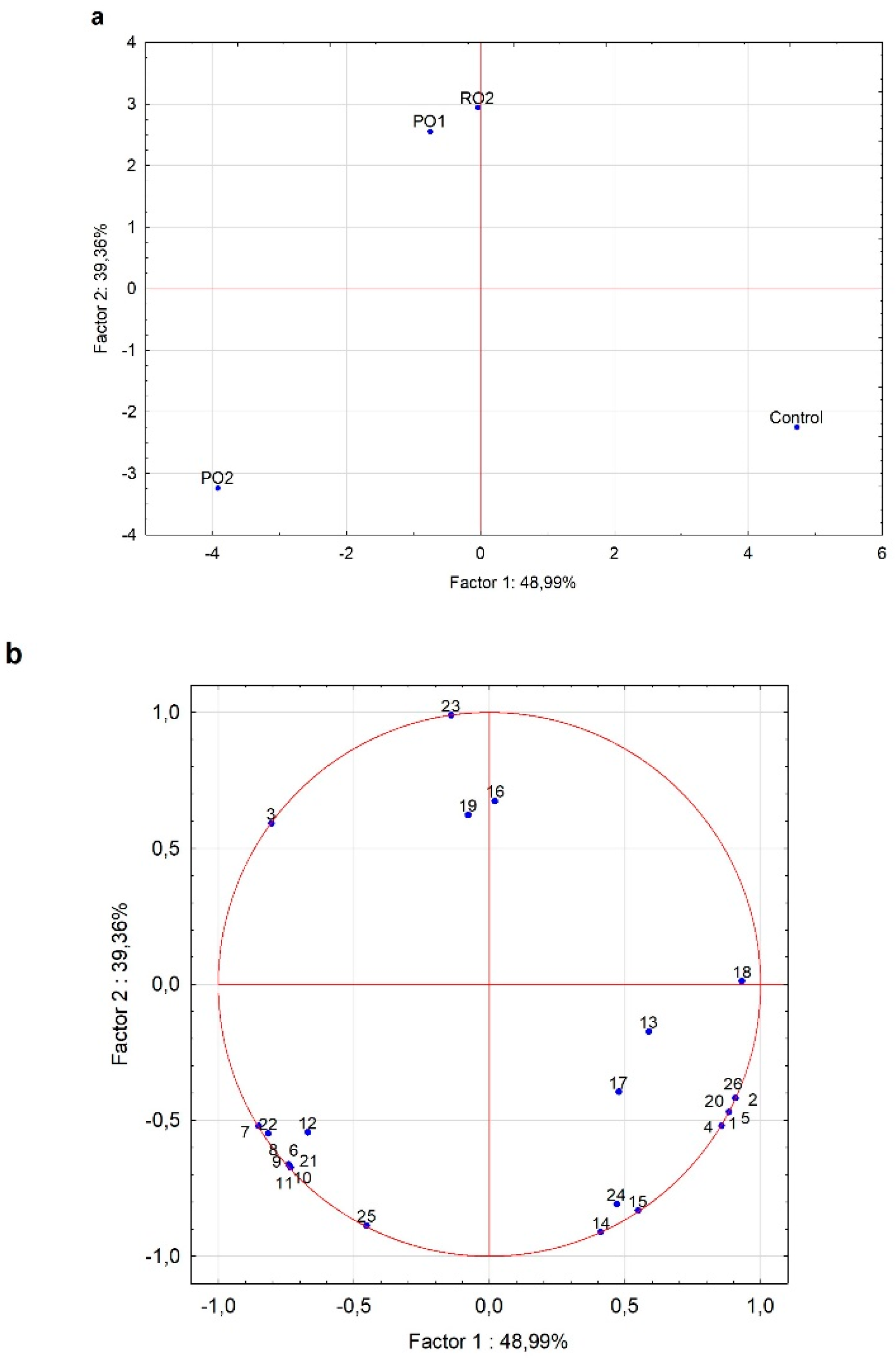
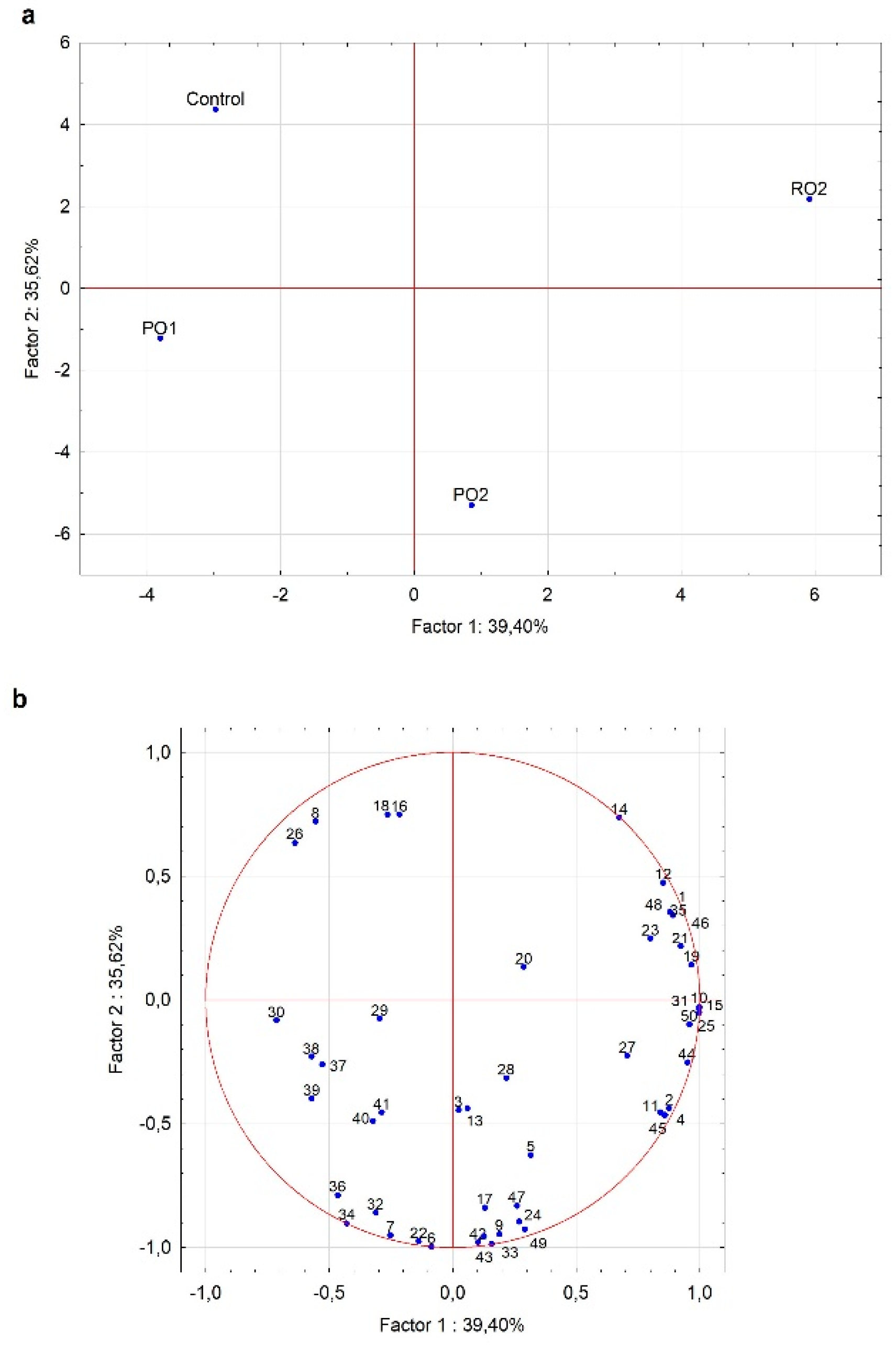
3.2.2. Screening Y. lipolytica Growth in Cheese Whey
3.3. Volatile Molecule Producation in Different Types of Whey by the Three Best Growing Strains
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gottardi, D.; Siroli, L.; Vannini, L.; Patrignani, F.; Lanciotti, R. Recovery and valorization of agri-food wastes and by-products using the non-conventional yeast Yarrowia lipolytica. Trends Food Sci. Technol. 2021, 115, 74–86. [Google Scholar] [CrossRef]
- Fröhlich-Wyder, M.T.; Arias-Roth, E.; Jakob, E. Cheese yeasts. Yeast 2019, 36, 129–141. [Google Scholar] [CrossRef]
- Groenewald, M.; Boekhout, T.; Neuvéglise, C.; Gaillardin, C.; van Dijck, P.W.; Wyss, M. Yarrowia lipolytica: Safety assessment of an oleaginous yeast with a great industrial potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef] [PubMed]
- Zieniuk, B.; Fabiszewska, A. Yarrowia lipolytica: A beneficious yeast in biotechnology as a rare opportunistic fungal pathogen: A minireview. World J. Microbiol. Biotechnol. 2019, 35, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazards, E.P.O.B.; Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernández Escámez, P.S.; Girones, R.; Koutsoumanis, K.; Lindqvist, R. Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 8: Suitability of taxonomic units notified to EFSA until March 2018. EFSA J. 2018, 16, e05315. [Google Scholar]
- EFSA NDA Panel (EFSA Panel on Nutrition, Novel Foods and Food Allergens); Turck, D.; Castenmiller, J.; de Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Safety of Yarrowia lipolytica yeast biomass as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2019, 17, e05594. [Google Scholar]
- Ryan, M.P.; Walsh, G. The biotechnological potential of whey. Rev. Environ. Sci. Bio/Technol. 2016, 15, 479–498. [Google Scholar] [CrossRef] [Green Version]
- Tsermoula, P.; Khakimov, B.; Nielsen, J.H.; Engelsen, S.B. Whey-The waste-stream that became more valuable than the food product. Trends Food Sci. Technol. 2021, 118, 230–241. [Google Scholar] [CrossRef]
- Yadav, J.S.S.; Yan, S.; Pilli, S.; Kumar, L.; Tyagi, R.D.; Surampalli, R.Y. Cheese whey: A potential resource to transform into bioprotein, functional/nutritional proteins and bioactive peptides. Biotechnol. Adv. 2015, 33, 756–774. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, Z.; Zhang, S.; Page, G.; Jaworski, N.W. The role of lactose in weanling pig nutrition: A literature and meta-analysis review. J. Anim. Sci. Biotechnol. 2021, 12, 1–17. [Google Scholar] [CrossRef]
- Pires, A.F.; Marnotes, N.G.; Rubio, O.D.; Garcia, A.C.; Pereira, C.D. Dairy by-products: A review on the valorization of whey and second cheese whey. Foods 2021, 10, 1067. [Google Scholar] [CrossRef]
- Barba, F.J. An integrated approach for the valorization of cheese whey. Foods 2021, 10, 564. [Google Scholar] [CrossRef]
- Zotta, T.; Solieri, L.; Iacumin, L.; Picozzi, C.; Gullo, M. Valorization of cheese whey using microbial fermentations. Appl. Microbiol. Biotechnol. 2020, 104, 2749–2764. [Google Scholar] [CrossRef]
- Kaur, R.; Panwar, D.; Panesar, P.S. Biotechnological approach for valorization of whey for value-added products. In Food Industry Wastes; Elsevier: Amsterdam, The Netherlands, 2020; pp. 275–302. [Google Scholar]
- Panesar, P.S.; Bera, M.B.; Kaur, S. Bioutilization of whey for ethanol production using yeast isolate. Int. J. Food Ferment. Technol. 2014, 4, 107. [Google Scholar]
- Ganju, S.; Gogate, P.R. A review on approaches for efficient recovery of whey proteins from dairy industry effluents. J. Food Eng. 2017, 215, 84–96. [Google Scholar] [CrossRef]
- Zerva, A.; Limnaios, A.; Kritikou, A.S.; Thomaidis, N.S.; Taoukis, P.; Topakas, E. A novel thermophile β-galactosidase from Thermothielavioides terrestris producing galactooligosaccharides from acid whey. New Biotechnol. 2021, 63, 45–53. [Google Scholar] [CrossRef]
- De Giorgi, S.; Raddadi, N.; Fabbri, A.; Toschi, T.G.; Fava, F. Potential use of ricotta cheese whey for the production of lactobionic acid by Pseudomonas taetrolens strains. New Biotechnol. 2018, 42, 71–76. [Google Scholar] [CrossRef]
- Frigon, M.D. Acid whey treatment and conversion to single cell protein via aerobic yeast activated sludge. Water Pract. Technol. 2020, 15, 494–505. [Google Scholar] [CrossRef]
- Rabaioli Rama, G.; Kuhn, D.; Beux, S.; Jachetti Maciel, M.; Volken de Souza, C.F. Cheese whey and ricotta whey for the growth and encapsulation of endogenous lactic acid bacteria. Food Bioprocess Technol. 2020, 13, 308–322. [Google Scholar] [CrossRef]
- Steephen, M.; Geetha, R.; Sathian, C. Activity profile of starter cultures maintained in whey based medium. Pharma Innov. J. 2020, 9, 127–129. [Google Scholar]
- Kurtzman, C.; Fell, J.W.; Boekhout, T. The Yeasts: A Taxonomic Study; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Taskin, M.; Saghafian, A.; Aydogan, M.N.; Arslan, N.P. Microbial lipid production by cold-adapted oleaginous yeast Yarrowia lipolytica B9 in non-sterile whey medium. Biofuels Bioprod. Biorefining 2015, 9, 595–605. [Google Scholar] [CrossRef]
- Yalcin, S.K.; Bozdemir, M.T.; Ozbas, Z.Y. Utilization of whey and grape must for citric acid production by two Yarrowia lipolytica strains. Food Biotechnol. 2009, 23, 266–283. [Google Scholar] [CrossRef]
- Arslan, N.P.; Aydogan, M.N.; Taskin, M. Citric acid production from partly deproteinized whey under non-sterile culture conditions using immobilized cells of lactose—Positive and cold-adapted Yarrowia lipolytica B9. J. Biotechnol. 2016, 231, 32–39. [Google Scholar] [CrossRef]
- Lanciotti, R.; Vannini, L.; Lopez, C.C.; Gobbetti, M.; Guerzoni, M.E. Evaluation of the ability of Yarrowia lipolytica to impart strain-dependent characteristics to cheese when used as a ripening adjunct. Int. J. Dairy Technol. 2005, 58, 89–99. [Google Scholar] [CrossRef]
- Patrignani, F.; Vannini, L.; Gardini, F.; Guerzoni, M.E.; Lanciotti, R. Variability of the lipolytic activity and volatile molecules production by a strain of Yarrowia lipolytica in pork fat and its dependence on environmental conditions. Meat Sci. 2011, 89, 21–26. [Google Scholar] [CrossRef]
- Patrignani, F.; Iucci, L.; Vallicelli, M.; Guerzoni, M.E.; Gardini, F.; Lanciotti, R. Role of surface-inoculated Debaryomyces hansenii and Yarrowia lipolytica strains in dried fermented sausage manufacture. Part 1: Evaluation of their effects on microbial evolution, lipolytic and proteolytic patterns. Meat Sci. 2007, 75, 676–686. [Google Scholar] [CrossRef]
- Lanciotti, R.; Gianotti, A.; Baldi, D.; Angrisani, R.; Suzzi, G.; Mastrocola, D.; Guerzoni, M. Use of Yarrowia lipolytica strains for the treatment of olive mill wastewater. Bioresour. Technol. 2005, 96, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Guerzoni, M.; Lanciotti, R.; Vannini, L.; Galgano, F.; Favati, F.; Gardini, F.; Suzzi, G. Variability of the lipolytic activity in Yarrowia lipolytica and its dependence on environmental conditions. Int. J. Food Microbiol. 2001, 69, 79–89. [Google Scholar] [CrossRef]
- Sinigaglia, M.; Lanciotti, R.; Guerzonil, M.E. Biochemical and physiological characteristics of Yarrowia lipolytica strains in relation to isolation source. Can. J. Microbiol. 1994, 40, 54–59. [Google Scholar] [CrossRef]
- Burns, P.; Patrignani, F.; Serrazanetti, D.; Vinderola, G.; Reinheimer, J.A.; Lanciotti, R.; Guerzoni, M.E. Probiotic Crescenza cheese containing Lactobacillus casei and Lactobacillus acidophilus manufactured with high-pressure homogenized milk. J. Dairy Sci. 2008, 91, 500–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braz, C.A.; Carvalho, A.K.; Bento, H.; Reis, C.E.; De Castro, H.F. Production of value-added microbial metabolites: Oleaginous fungus as a tool for valorization of dairy by-products. BioEnergy Res. 2020, 13, 963–973. [Google Scholar] [CrossRef]
- Costa, S.; Summa, D.; Semeraro, B.; Zappaterra, F.; Rugiero, I.; Tamburini, E. Fermentation as a strategy for bio-transforming waste into resources: Lactic acid production from agri-food residues. Fermentation 2020, 7, 3. [Google Scholar] [CrossRef]
- Brizuela, N.S.; Arnez-Arancibia, M.; Semorile, L.; Bravo-Ferrada, B.M.; Tymczyszyn, E.E. Whey permeate as a substrate for the production of freeze-dried Lactiplantibacillus plantarum to be used as a malolactic starter culture. World J. Microbiol. Biotechnol. 2021, 37, 1–12. [Google Scholar] [CrossRef]
- Gardini, F.; Tofalo, R.; Belletti, N.; Iucci, L.; Suzzi, G.; Torriani, S.; Guerzoni, M.; Lanciotti, R. Characterization of yeasts involved in the ripening of Pecorino Crotonese cheese. Food Microbiol. 2006, 23, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Tofalo, R.; Schirone, M.; Fasoli, G.; Perpetuini, G.; Patrignani, F.; Manetta, A.C.; Lanciotti, R.; Corsetti, A.; Martino, G.; Suzzi, G. Influence of pig rennet on proteolysis, organic acids content and microbiota of Pecorino di Farindola, a traditional Italian ewe’s raw milk cheese. Food Chem. 2015, 175, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Vannini, L.; Patrignani, F.; Iucci, L.; Ndagijimana, M.; Vallicelli, M.; Lanciotti, R.; Guerzoni, M.E. Effect of a pre-treatment of milk with high pressure homogenization on yield as well as on microbiological, lipolytic and proteolytic patterns of “Pecorino” cheese. Int. J. Food Microbiol. 2008, 128, 329–335. [Google Scholar] [CrossRef]
- Lanciotti, R.; Vannini, L.; Patrignani, F.; Iucci, L.; Vallicelli, M.; Ndagijimana, M.; Guerzoni, M.E. Effect of high pressure homogenisation of milk on cheese yield and microbiology, lipolysis and proteolysis during ripening of Caciotta cheese. J. Dairy Res. 2006, 73, 216–226. [Google Scholar] [CrossRef]
- Ganesan, B.; Weimer, B.C. Amino acid catabolism and its relationship to cheese flavor outcomes. In Cheese; Elsevier: Amsterdam, The Netherlands, 2017; pp. 483–516. [Google Scholar]
- Ardö, Y. Enzymes in Cheese Ripening. In Agents Change; Springer Nature: Berlin, Germany, 2021; pp. 363–395. [Google Scholar]
- Vannini, L.; Baldi, D.; Lanciotti, R. Use of Fourier transform infrared spectroscopy to evaluate the proteolytic activity of Yarrowia lipolytica and its contribution to cheese ripening. Int. J. Food Microbiol. 2001, 69, 113–123. [Google Scholar]
- Suzzi, G.; Lanorte, M.; Galgano, F.; Andrighetto, C.; Lombardi, A.; Lanciotti, R.; Guerzoni, M. Proteolytic, lipolytic and molecular characterisation of Yarrowia lipolytica isolated from cheese. Int. J. Food Microbiol. 2001, 69, 69–77. [Google Scholar] [CrossRef]
- Rossi, S.; Parrotta, L.; Del Duca, S.; Dalla Rosa, M.; Patrignani, F.; Schluter, O.; Lanciotti, R. Effect of Yarrowia lipolytica RO25 cricket-based hydrolysates on sourdough quality parameters. LWT-Food Sci. Technol. 2021, 148, 111760. [Google Scholar] [CrossRef]
- Szoltysik, M.; Dabrowska, A.; Babij, K.; Pokora, M.; Zambrowicz, A.; Polomska, X.; Wojtatowicz, M.; Chrzanowska, J. Biochemical and microbiological changes in cheese inoculated with Yarrowia lipolytica yeast. Żywność Nauka Technol. Jakość 2013, 4, 49–64. [Google Scholar] [CrossRef]
- Guerzoni, M.; Vannini, L.; Chaves Lopez, C.; Gobbetti, M.; Lanciotti, R. Yarrowia lipolytica as potential ripening agent in milk products. In Proceedings of the Yeasts in the Dairy Industry: Positive and Negative Aspects, Copenhagen, Denmark, 2–3 September 1996. [Google Scholar]
- Fickers, P.; Benetti, P.-H.; Waché, Y.; Marty, A.; Mauersberger, S.; Smit, M.; Nicaud, J.-M. Hydrophobic substrate utilisation by the yeast Yarrowia lipolytica, and its potential applications. FEMS Yeast Res. 2005, 5, 527–543. [Google Scholar] [CrossRef] [Green Version]
- Brígida, A.I.; Amaral, P.F.; Coelho, M.A.; Goncalves, L.R. Lipase from Yarrowia lipolytica: Production, characterization and application as an industrial biocatalyst. J. Mol. Catal. B Enzym. 2014, 101, 148–158. [Google Scholar] [CrossRef]
- Kumari, A.; Gupta, R. Extracellular expression and characterization of thermostable lipases, LIP8, LIP14 and LIP18, from Yarrowia lipolytica. Biotechnol. Lett. 2012, 34, 1733–1739. [Google Scholar] [CrossRef]
- Kumari, A.; Verma, V.V.; Gupta, R. Comparative biochemical characterization and in silico analysis of novel lipases Lip11 and Lip12 with Lip2 from Yarrowia lipolytica. World J. Microbiol. Biotechnol. 2012, 28, 3103–3111. [Google Scholar] [CrossRef] [PubMed]
- Thierry, A.; Collins, Y.F.; Mukdsi, M.A.; McSweeney, P.L.; Wilkinson, M.G.; Spinnler, H.E. Lipolysis and metabolism of fatty acids in cheese. In Cheese; Elsevier: Amsterdam, The Netherlands, 2017; pp. 423–444. [Google Scholar]
- Rivière, A.; Moens, F.; Selak, M.; Maes, D.; Weckx, S.; De Vuyst, L. The ability of bifidobacteria to degrade arabinoxylan oligosaccharide constituents and derived oligosaccharides is strain dependent. Appl. Environ. Microbiol. 2014, 80, 204–217. [Google Scholar] [CrossRef] [Green Version]
- Alonso García, E.; de la Fuente Ordoñez, J.J.; Lavilla Lerma, L.; Estudillo-Martínez, M.D.; Castillo-Gutiérrez, S.; Benomar, N.; Knapp, C.W.; Abriouel, H. Transcriptomic profile and probiotic properties of Lactiplantibacillus pentosus pre-adapted to edible oils. Front. Microbiol. 2021, 12, 747043. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, S.J.; Pannella, G.; Iorizzo, M.; Testa, B.; Succi, M.; Tremonte, P.; Sorrentino, E.; Di Renzo, M.; Strollo, D.; Coppola, R. Inoculum strategies and performances of malolactic starter Lactobacillus plantarum M10: Impact on chemical and sensorial characteristics of Fiano Wine. Microorganisms 2020, 8, 516. [Google Scholar] [CrossRef] [Green Version]
- Martins, D.; English, A.M. Catalase activity is stimulated by H2O2 in rich culture medium and is required for H2O2 resistance and adaptation in yeast. Redox Biol. 2014, 2, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Chin, Y.-W.; Lee, S.; Yu, H.H.; Yang, S.J.; Kim, T.-W. Combinatorial effects of protective agents on survival rate of the yeast starter, Saccharomyces cerevisiae 88-4, after Freeze-Drying. Microorganisms 2021, 9, 613. [Google Scholar] [CrossRef]
- Guowei, S.; Yang, X.; Li, C.; Huang, D.; Lei, Z.; He, C. Comprehensive optimization of composite cryoprotectant for Saccharomyces boulardii during freeze-drying and evaluation of its storage stability. Prep. Biochem. Biotechnol. 2019, 49, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Abadias, M.; Benabarre, A.; Teixidó, N.; Usall, J.; Vinas, I. Effect of freeze drying and protectants on viability of the biocontrol yeast Candida sake. Int. J. Food Microbiol. 2001, 65, 173–182. [Google Scholar] [CrossRef]
- Gul, L.B.; Con, A.H.; Gul, O. Storage stability and sourdough acidification kinetic of freeze-dried Lactobacillus curvatus N19 under optimized cryoprotectant formulation. Cryobiology 2020, 96, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Mazorra-Manzano, M.A.; Robles-Porchas, G.R.; González-Velázquez, D.A.; Torres-Llanez, M.J.; Martínez-Porchas, M.; García-Sifuentes, C.O.; González-Córdova, A.F.; Vallejo-Córdoba, B. Cheese whey fermentation by its native microbiota: Proteolysis and bioactive peptides release with ACE-inhibitory activity. Fermentation 2020, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Mansour, S.; Beckerich, J.; Bonnarme, P. Lactate and amino acid catabolism in the cheese-ripening yeast Yarrowia lipolytica. Appl. Environ. Microbiol. 2008, 74, 6505–6512. [Google Scholar] [CrossRef] [PubMed]
- Ariana, M.; Hamedi, J. Enhanced production of nisin by co-culture of Lactococcus lactis sub sp. lactis and Yarrowia lipolytica in molasses based medium. J. Biotechnol. 2017, 256, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Louhasakul, Y.; Cheirsilp, B.; Treu, L.; Kougias, P.G.; Angelidaki, I. Metagenomic insights into bioaugmentation and biovalorization of oily industrial wastes by lipolytic oleaginous yeast Yarrowia lipolytica during successive batch fermentation. Biotechnol. Appl. Biochem. 2020, 67, 1020–1029. [Google Scholar] [CrossRef] [PubMed]
- López del Castillo-Lozano, M.; Delile, A.; Spinnler, H.; Bonnarme, P.; Landaud, S. Comparison of volatile sulphur compound production by cheese-ripening yeasts from methionine and methionine–cysteine mixtures. Appl. Microbiol. Biotechnol. 2007, 75, 1447–1454. [Google Scholar] [CrossRef]
- Martin, N.; Berger, C.; Le Du, C.; Spinnler, H. Aroma compound production in cheese curd by coculturing with selected yeast and bacteria. J. Dairy Sci. 2001, 84, 2125–2135. [Google Scholar] [CrossRef]
- Spinnler, H.; Berger, C.; Lapadatescu, C.; Bonnarme, P. Production of sulfur compounds by several yeasts of technological interest for cheese ripening. Int. Dairy J. 2001, 11, 245–252. [Google Scholar] [CrossRef]
- Fox, P.F.; McSweeney, P.L.; Cogan, T.M.; Guinee, T.P. Cheese: Chemistry, Physics and Microbiology, Volume 1: General Aspects; Elsevier: Amsterdam, The Netherlands, 2004. [Google Scholar]
- Cozzolino, R.; Martignetti, A.; De Giulio, B.; Malorni, L.; Addeo, F.; Picariello, G. SPME GC-MS monitoring of volatile organic compounds to assess typicity of Pecorino di Carmasciano ewe-milk cheese. Int. J. Dairy Technol. 2021, 74, 383–392. [Google Scholar] [CrossRef]
- Marilley, L.; Casey, M. Flavours of cheese products: Metabolic pathways, analytical tools and identification of producing strains. Int. J. Food Microbiol. 2004, 90, 139–159. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
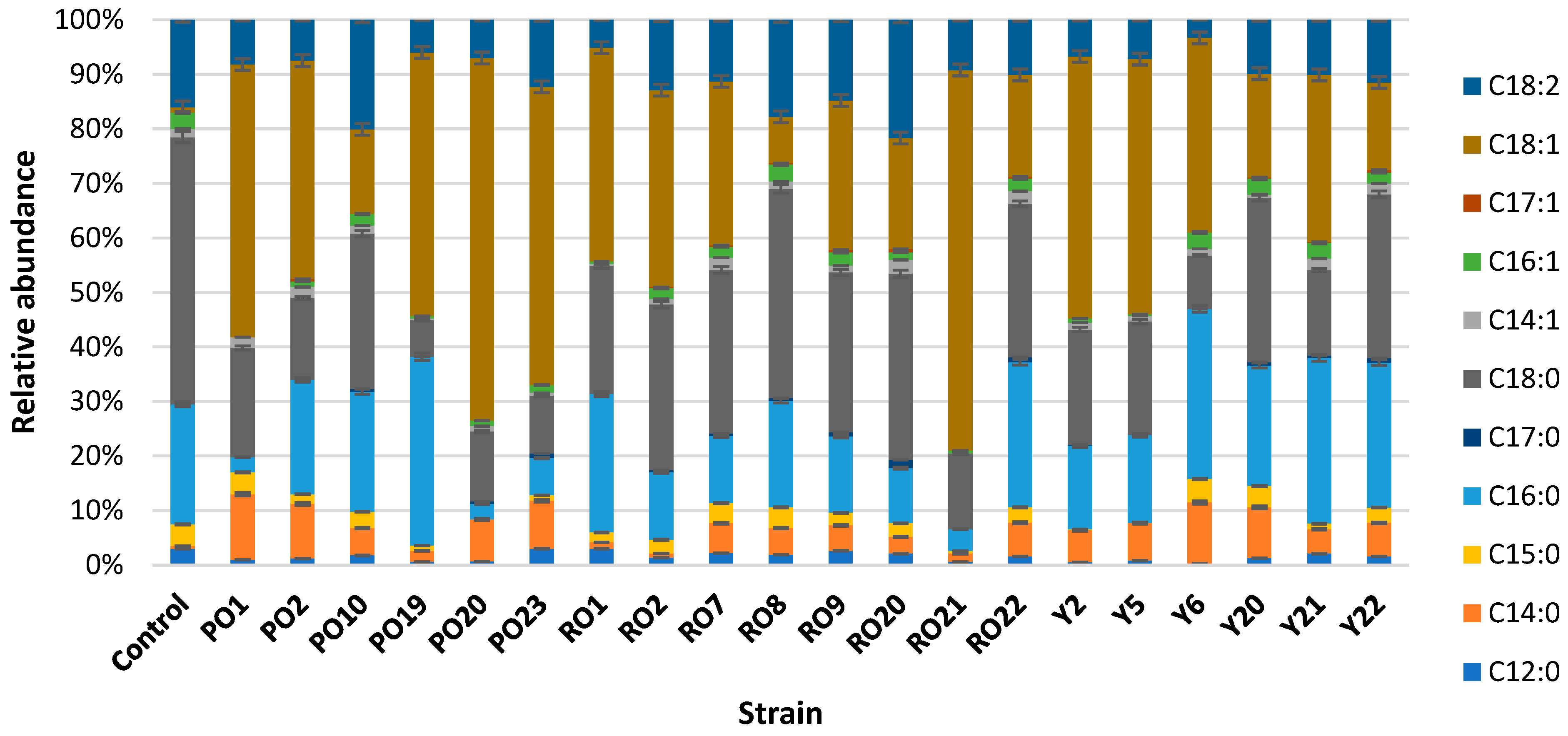{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Measuring Unit | Caciotta Whey | Ricotta Whey | Squacquerone Whey | Mixed Whey |
|---|---|---|---|---|---|
| Fats | g/L | 4.7 ± 1.5 a | 3.4 ± 0.3 a | 2.9 ± 0.1 b | 3.5 ± 2.1 a |
| Proteins | g/L | 10.7 ± 1.5 a | 6.0 ± 0.1 b | 9.9 ± 1.9 a | 11.0 ± 0.9 a |
| Lactose | g/L | 46.9 ± 0.8 a | 45.8 ± 1.5 a | 48.8 ± 0.9 b | 40.5 ± 0.2 c |
| Glucose | g/L | 1.3 ± 0.5 a | <0.1 b | 2.0 ± 0.8 a | <0.1 b |
| Lactic acid | g/L | 1.5 ± 0.8 a | 1.2 ± 1.1 a | 1.3 ± 0.5 a | 7.7 ± 0.1 b |
| Chemical Oxygen Demand (COD) | g/L O2 | 90.9 ± 4.2 | 78.6 ± 14.9 | 84.7 ± 3.8 | 81.1 ± 14.0 |
| Total nitrogen | % | 0.17 ± 0.03 | 0.10 ± 0.02 | 0.16 ± 0.03 | 0.18 ± 0.01 |
| pH | 6.5 ± 0.06 a | 6.0 ± 0.07 a | 6.5 ± 0.11 a | 3.6 ± 0.11 b | |
| Phosphorous | mg/kg | 450 ± 52 | 351 ± 82 | 448 ± 46 | 421 ± 18 |
| Phosphates as PO4 | mg/kg | 1379 ± 158 | 1075 ± 249 | 1373 ± 141 | 1291 ± 56 |
| Calcium | mg/kg | 380 ± 64 | 354 ± 72 | 361 ± 129 | 328 ± 18 |
| Potassium | mg/kg | 1450 ± 145 | 1428 ± 92 | 1451 ± 151 | 1449 ± 62 |
| Sodium | mg/kg | 3386 ± 332 | 2588 ± 356 | 2672 ± 350 | 2913 ± 164 |
| Iodine | mg/kg | 0.48 ± 0.10 | 3.02 ± 0.36 | 0.47 ± 0.14 | - |
| Caciotta Whey | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Concentration (log CFU/g) | pH | |||||||||||||||||||||||||||||
| Strain | 0 h | 24 h | 48 h | 72 h | 0 h | 24 h | 48 h | 72 h | ||||||||||||||||||||||
| PO1 | 4.9 | ± | 0.2 | 6.5 | m | ± | 0.06 | 7.1 | m,n | ± | 0.01 | 8.8 | i | ± | 0.15 | 6.5 | ± | 0.1 | 6.4 | i,l | ± | 0.02 | 5.2 | g | ± | 0.01 | 4.2 | a,b,c | ± | 0.02 |
| PO10 | 4.9 | ± | 0.2 | 6.0 | f,g,h | ± | 0.13 | 6.1 | e,f | ± | 0.02 | 6.5 | c,d,e | ± | 0.12 | 6.5 | ± | 0.1 | 6.5 | l | ± | 0.03 | 5.1 | f | ± | 0.02 | 4.3 | a,b,c | ± | 0.01 |
| PO19 | 4.9 | ± | 0.2 | 6.1 | f,g,h,i | ± | 0.08 | 6.6 | g,h,i,l | ± | 0.09 | 6.3 | b,c,d | ± | 0.04 | 6.5 | ± | 0.1 | 6.4 | h,i | ± | 0.01 | 5.1 | e,f | ± | 0.05 | 4.8 | f | ± | 0.01 |
| PO2 | 4.9 | ± | 0.2 | 6.3 | i,l | ± | 0.18 | 6.8 | h,i,l,m | ± | 0.06 | 8.6 | i | ± | 0.03 | 6.5 | ± | 0.1 | 6.4 | i,l | ± | 0.05 | 5.5 | h | ± | 0.01 | 4.5 | d,e | ± | 0.01 |
| PO20 | 4.9 | ± | 0.2 | 6.1 | g,h,i | ± | 0.11 | 6.4 | f,g,h | ± | 0.10 | 6.5 | c,d | ± | 0.01 | 6.5 | ± | 0.1 | 6.0 | c | ± | 0.01 | 4.5 | a | ± | 0.03 | 4.5 | d | ± | 0.02 |
| PO23 | 4.9 | ± | 0.2 | 5.0 | c | ± | 0.01 | 5.1 | b | ± | 0.13 | 5.0 | a | ± | 0.01 | 6.5 | ± | 0.1 | 6.3 | f,g | ± | 0.01 | 4.5 | a | ± | 0.01 | 4.2 | a | ± | 0.02 |
| RO1 | 4.9 | ± | 0.2 | 6.2 | h,i | ± | 0.06 | 6.9 | i,l,m | ± | 0.12 | 6.7 | d,e,f | ± | 0.05 | 6.5 | ± | 0.1 | 5.9 | b | ± | 0.02 | 5.0 | e | ± | 0.03 | 5.3 | i,l | ± | 0.23 |
| RO2 | 4.9 | ± | 0.2 | 6.9 | a | ± | 0.01 | 7.3 | n | ± | 0.25 | 7.5 | f | ± | 0.10 | 6.5 | ± | 0.1 | 5.6 | a | ± | 0.03 | 5.2 | g | ± | 0.01 | 5.4 | l | ± | 0.12 |
| RO20 | 4.9 | ± | 0.2 | 5.0 | c | ± | 0.01 | 5.3 | b,c | ± | 0.04 | 5.0 | a | ± | 0.01 | 6.5 | ± | 0.1 | 6.1 | c,d | ± | 0.04 | 4.7 | b | ± | 0.02 | 4.6 | d,e | ± | 0.03 |
| RO21 | 4.9 | ± | 0.2 | 5.8 | e | ± | 0.02 | 6.3 | e,f,g | ± | 0.03 | 6.2 | b,c | ± | 0.01 | 6.5 | ± | 0.1 | 6.0 | c | ± | 0.06 | 4.7 | b,c | ± | 0.02 | 4.4 | b,c,d | ± | 0.01 |
| RO22 | 4.9 | ± | 0.2 | 4.6 | b | ± | 0.01 | 4.0 | a | ± | 0.26 | 5.0 | a | ± | 0.01 | 6.5 | ± | 0.1 | 6.2 | d,e | ± | 0.04 | 4.8 | d | ± | 0.01 | 4.3 | a,b,c | ± | 0.01 |
| RO7 | 4.9 | ± | 0.2 | 5.9 | e,f,g | ± | 0.11 | 6.4 | f,g,h | ± | 0.16 | 8.3 | h | ± | 0.12 | 6.5 | ± | 0.1 | 6.5 | l | ± | 0.01 | 5.0 | e | ± | 0.04 | 4.4 | c,d | ± | 0.01 |
| RO8 | 4.9 | ± | 0.2 | 6.0 | e,f,g,h | ± | 0.01 | 6.5 | f,g,h,i | ± | 0.18 | 7.7 | g | ± | 0.19 | 6.5 | ± | 0.1 | 6.4 | i,l | ± | 0.01 | 5.3 | g | ± | 0.01 | 4.2 | a,b | ± | 0.05 |
| RO9 | 4.9 | ± | 0.2 | 6.1 | g,h,i | ± | 0.04 | 6.6 | g,h,i,l | ± | 0.03 | 7.1 | g | ± | 0.02 | 6.5 | ± | 0.1 | 6.4 | i,l | ± | 0.05 | 4.7 | b,c | ± | 0.03 | 4.1 | a | ± | 0.01 |
| Y2 | 4.9 | ± | 0.2 | 6.4 | l,m | ± | 0.07 | 6.9 | l,m | ± | 0.08 | 6.8 | f | ± | 0.01 | 6.5 | ± | 0.1 | 6.4 | i,l | ± | 0.02 | 5.0 | e | ± | 0.01 | 4.8 | f,g | ± | 0.01 |
| Y20 | 4.9 | ± | 0.2 | 4.6 | b | ± | 0.05 | 5.0 | b | ± | 0.02 | 5.0 | a | ± | 0.01 | 6.5 | ± | 0.1 | 6.3 | g,h | ± | 0.02 | 4.8 | d | ± | 0.01 | 5.0 | g,h | ± | 0.02 |
| Y21 | 4.9 | ± | 0.2 | 5.4 | d | ± | 0.01 | 5.7 | c,d | ± | 0.18 | 5.0 | a | ± | 0.02 | 6.5 | ± | 0.1 | 6.2 | e,f | ± | 0.01 | 4.8 | c,d | ± | 0.04 | 4.8 | f,g | ± | 0.02 |
| Y22 | 4.9 | ± | 0.2 | 5.9 | e,f | ± | 0.02 | 6.0 | d,e | ± | 0.07 | 5.8 | b | ± | 0.01 | 6.5 | ± | 0.1 | 6.1 | d | ± | 0.01 | 4.6 | b | ± | 0.05 | 4.8 | f,g | ± | 0.01 |
| Y5 | 4.9 | ± | 0.2 | 6.4 | l,m | ± | 0.03 | 6.8 | h,i,l,m | ± | 0.18 | 7.0 | g | ± | 0.02 | 6.5 | ± | 0.1 | 6.4 | i,l | ± | 0.03 | 5.3 | g | ± | 0.01 | 4.7 | e,f | ± | 0.01 |
| Y6 | 4.9 | ± | 0.2 | 6.4 | l,m | ± | 0.01 | 6.6 | g,h,i,l | ± | 0.13 | 7.5 | f,g | ± | 0.07 | 6.5 | ± | 0.1 | 6.5 | l | ± | 0.04 | 5.6 | i | ± | 0.01 | 5.1 | h,i | ± | 0.01 |
| Ricotta Whey | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Concentration (log CFU/g) | pH | |||||||||||||||||||||||||||||
| 0 h | 24 h | 48 h | 72 h | 0 h | 24 h | 48 h | 72 h | |||||||||||||||||||||||
| PO1 | 4.9 | ± | 0.2 | 6.2 | e,f,g,h | ± | 0.01 | 6.9 | f,g | ± | 0.16 | 7.3 | f,g,h | ± | 0.03 | 6.0 | ± | 0.07 | 6.0 | f,g | ± | 0.02 | 5.9 | g,h | ± | 0.02 | 7.0 | l | ± | 0.01 |
| PO10 | 4.9 | ± | 0.2 | 5.8 | d,e | ± | 0.01 | 6.3 | c,d | ± | 0.02 | 6.0 | b | ± | 0.01 | 6.0 | ± | 0.07 | 6.0 | f,g | ± | 0.01 | 6.2 | m,n | ± | 0.06 | 6.7 | i | ± | 0.01 |
| PO19 | 4.9 | ± | 0.2 | 7.0 | i | ± | 0.02 | 7.4 | h | ± | 0.01 | 7.7 | i | ± | 0.01 | 6.0 | ± | 0.07 | 6.1 | i | ± | 0.04 | 6.2 | l,m | ± | 0.01 | 5.9 | f,g | ± | 0.05 |
| PO2 | 4.9 | ± | 0.2 | 6.4 | g,h | ± | 0.10 | 7.0 | f,g | ± | 0.19 | 7.9 | i | ± | 0.08 | 6.0 | ± | 0.07 | 6.1 | i | ± | 0.01 | 6.2 | l,m | ± | 0.01 | 7.0 | l | ± | 0.11 |
| PO20 | 4.9 | ± | 0.2 | 6.9 | i | ± | 0.01 | 7.2 | g,h | ± | 0.01 | 7.9 | i | ± | 0.01 | 6.0 | ± | 0.07 | 6.2 | l | ± | 0.02 | 6.3 | n | ± | 0.01 | 6.6 | i | ± | 0.06 |
| PO23 | 4.9 | ± | 0.2 | 5.3 | b | ± | 0.01 | 5.8 | b | ± | 0.01 | 6.4 | c | ± | 0.09 | 6.0 | ± | 0.07 | 6.1 | h,i | ± | 0.01 | 6.2 | m,n | ± | 0.01 | 6.4 | h,i | ± | 0.02 |
| RO1 | 4.9 | ± | 0.2 | 5.7 | c,d | ± | 0.12 | 5.9 | b | ± | 0.11 | 5.0 | a | ± | 0.01 | 6.0 | ± | 0.07 | 5.4 | a | ± | 0.01 | 4.4 | a | ± | 0.10 | 4.2 | a | ± | 0.01 |
| RO2 | 4.9 | ± | 0.2 | 6.9 | i | ± | 0.13 | 7.2 | g,h | ± | 0.07 | 7.3 | g,h | ± | 0.19 | 6.0 | ± | 0.07 | 5.4 | a | ± | 0.01 | 4.8 | b | ± | 0.08 | 4.3 | a | ± | 0.03 |
| RO20 | 4.9 | ± | 0.2 | 5.4 | b,c | ± | 0.01 | 5.8 | b | ± | 0.09 | 6.8 | d,e | ± | 0.07 | 6.0 | ± | 0.07 | 5.7 | c | ± | 0.02 | 5.4 | c,d | ± | 0.01 | 5.6 | d,e,f | ± | 0.11 |
| RO21 | 4.9 | ± | 0.2 | 6.3 | f,g,h | ± | 0.11 | 6.4 | c,d | ± | 0.02 | 7.3 | f,g,h | ± | 0.08 | 6.0 | ± | 0.07 | 5.9 | d | ± | 0.02 | 5.6 | e | ± | 0.10 | 4.8 | b | ± | 0.04 |
| RO22 | 4.9 | ± | 0.2 | 4.6 | a | ± | 0.08 | 5.0 | a | ± | 0.02 | 5.0 | a | ± | 0.01 | 6.0 | ± | 0.07 | 6.0 | e,f | ± | 0.03 | 5.6 | e | ± | 0.01 | 4.9 | b | ± | 0.01 |
| RO7 | 4.9 | ± | 0.2 | 5.4 | b,c | ± | 0.40 | 6.4 | c,d | ± | 0.05 | 6.3 | b,c | ± | 0.01 | 6.0 | ± | 0.07 | 6.0 | f,g | ± | 0.01 | 6.0 | h,i | ± | 0.01 | 5.8 | e,f | ± | 0.15 |
| RO8 | 4.9 | ± | 0.2 | 5.9 | d,e,f | ± | 0.15 | 6.5 | c,d | ± | 0.11 | 7.1 | e,f,g | ± | 0.23 | 6.0 | ± | 0.07 | 6.0 | g,h | ± | 0.02 | 6.1 | i,l | ± | 0.01 | 6.6 | i | ± | 0.21 |
| RO9 | 4.9 | ± | 0.2 | 6.4 | g,h | ± | 0.02 | 6.9 | f,g | ± | 0.1 | 7.6 | h,i | ± | 0.13 | 6.0 | ± | 0.07 | 6.0 | f,g | ± | 0.01 | 5.8 | f,g | ± | 0.06 | 5.5 | c,d,e | ± | 0.23 |
| Y2 | 4.9 | ± | 0.2 | 6.5 | h | ± | 0.05 | 7.0 | f,g | ± | 0.18 | 6.6 | c,d | ± | 0.01 | 6.0 | ± | 0.07 | 5.9 | d,e | ± | 0.01 | 5.4 | d | ± | 0.04 | 5.3 | c | ± | 0.18 |
| Y20 | 4.9 | ± | 0.2 | 6.1 | e,f,g | ± | 0.01 | 6.2 | c | ± | 0.03 | 6.3 | b,c | ± | 0.02 | 6.0 | ± | 0.07 | 6.1 | i | ± | 0.01 | 5.7 | e,f | ± | 0.01 | 4.4 | a | ± | 0.04 |
| Y21 | 4.9 | ± | 0.2 | 6.2 | e,f,g,h | ± | 0.12 | 6.8 | e,f | ± | 0.1 | 7.0 | e,f | ± | 0.20 | 6.0 | ± | 0.07 | 6.1 | i | ± | 0.03 | 6.3 | m,n | ± | 0.03 | 6.1 | g,h | ± | 0.06 |
| Y22 | 4.9 | ± | 0.2 | 6.3 | g,h | ± | 0.08 | 6.9 | f,g | ± | 0.04 | 7.1 | e,f,g | ± | 0.02 | 6.0 | ± | 0.07 | 5.9 | d,e | ± | 0.03 | 5.3 | c | ± | 0.04 | 4.9 | b | ± | 0.01 |
| Y5 | 4.9 | ± | 0.2 | 6.5 | h | ± | 0.07 | 6.6 | d,e | ± | 0.16 | 6.9 | d,e | ± | 0.02 | 6.0 | ± | 0.07 | 5.9 | d | ± | 0.04 | 5.5 | d | ± | 0.04 | 5.3 | c,d | ± | 0.05 |
| Y6 | 4.9 | ± | 0.2 | 6.2 | f,g,h | ± | 0.09 | 6.5 | c,d | ± | 0.04 | 7.0 | e,f,g | ± | 0.30 | 6.0 | ± | 0.07 | 5.7 | b | ± | 0.02 | 5.7 | e,f | ± | 0.01 | 5.8 | e,f | ± | 0.02 |
| Squacquerone Whey | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Concentration (log CFU/g) | pH | |||||||||||||||||||||||||||||
| 24 h | 48 h | 72 h | 0 h | 24 h | 48 h | 72 h | ||||||||||||||||||||||||
| PO1 | 4.9 | ± | 0.2 | 6.5 | h,i,l | ± | 0.10 | 7.2 | i | ± | 0.26 | 7.7 | h | ± | 0.02 | 6.5 | ± | 0.11 | 6.8 | b | ± | 0.05 | 5.8 | l | ± | 0.04 | 5.1 | g,h | ± | 0.21 |
| PO10 | 4.9 | ± | 0.2 | 5.9 | e | ± | 0.08 | 6.3 | d,e | ± | 0.02 | 6.5 | c,d | ± | 0.01 | 6.5 | ± | 0.11 | 6.6 | f,g | ± | 0.01 | 5.4 | g,h,i | ± | 0.01 | 4.9 | d,e,f | ± | 0.12 |
| PO19 | 4.9 | ± | 0.2 | 6.2 | f,g,h | ± | 0.04 | 6.8 | f,g,h,i | ± | 0.06 | 7.6 | g,h | ± | 0.12 | 6.5 | ± | 0.11 | 6.6 | f,g | ± | 0.02 | 4.9 | b | ± | 0.09 | 4.4 | a,b | ± | 0.03 |
| PO2 | 4.9 | ± | 0.2 | 6.4 | g,h,i,l | ± | 0.06 | 6.9 | g,h,i | ± | 0.19 | 7.3 | f,g | ± | 0.05 | 6.5 | ± | 0.11 | 6.6 | e,f,g | ± | 0.02 | 5.2 | d,e,f,g | ± | 0.01 | 4.9 | d,e,f | ± | 0.09 |
| PO20 | 4.9 | ± | 0.2 | 6.0 | e,f | ± | 0.11 | 6.4 | e | ± | 0.10 | 6.9 | e,f | ± | 0.14 | 6.5 | ± | 0.11 | 6.5 | d,e,f,g | ± | 0.01 | 5.1 | b,c,d,e | ± | 0.01 | 4.8 | c,d | ± | 0.01 |
| PO23 | 4.9 | ± | 0.2 | 5.0 | b | ± | 0.01 | 5.1 | b | ± | 0.20 | 5.7 | b | ± | 0.26 | 6.5 | ± | 0.11 | 6.4 | c,d | ± | 0.03 | 4.5 | a | ± | 0.12 | 4.2 | a | ± | 0.02 |
| RO1 | 4.9 | ± | 0.2 | 6.5 | i,l | ± | 0.06 | 6.6 | e,f,g | ± | 0.26 | 6.5 | c,d | ± | 0.19 | 6.5 | ± | 0.11 | 6.0 | a | ± | 0.01 | 4.5 | a | ± | 0.21 | 4.4 | b | ± | 0.01 |
| RO2 | 4.9 | ± | 0.2 | 7.6 | m | ± | 0.04 | 8.4 | l | ± | 0.05 | 8.3 | i | ± | 0.08 | 6.5 | ± | 0.11 | 6.4 | c,d | ± | 0.11 | 6.5 | m | ± | 0.10 | 5.8 | i | ± | 0.03 |
| RO20 | 4.9 | ± | 0.2 | 5.1 | b,c | ± | 0.01 | 5.7 | c | ± | 0.09 | 6.2 | c | ± | 0.29 | 6.5 | ± | 0.11 | 6.4 | c | ± | 0.01 | 5.3 | e,f,g,h | ± | 0.03 | 5.1 | f,g,h | ± | 0.03 |
| RO21 | 4.9 | ± | 0.2 | 6.2 | f,g | ± | 0.08 | 6.4 | d,e | ± | 0.04 | 7.2 | f,g | ± | 0.11 | 6.5 | ± | 0.11 | 6.5 | c,d,e,f | ± | 0.09 | 4.9 | b | ± | 0.05 | 4.9 | d,e | ± | 0.01 |
| RO22 | 4.9 | ± | 0.2 | 4.1 | a | ± | 0.02 | 4.4 | a | ± | 0.07 | 5.0 | a | ± | 0.01 | 6.5 | ± | 0.11 | 6.6 | e,f,g | ± | 0.01 | 5.6 | i,l | ± | 0.01 | 5.2 | h | ± | 0.01 |
| RO7 | 4.9 | ± | 0.2 | 6.0 | e,f | ± | 0.21 | 6.4 | d,e | ± | 0.05 | 6.3 | c | ± | 0.01 | 6.5 | ± | 0.11 | 6.7 | b,g | ± | 0.04 | 5.5 | h,i | ± | 0.04 | 5.1 | e,f,g,h | ± | 0.05 |
| RO8 | 4.9 | ± | 0.2 | 6.1 | e,f | ± | 0.19 | 5.9 | c,d | ± | 0.17 | 6.7 | d,e | ± | 0.08 | 6.5 | ± | 0.11 | 6.6 | e,f,g | ± | 0.05 | 5.3 | f,g,h,i | ± | 0.01 | 4.8 | c,d | ± | 0.01 |
| RO9 | 4.9 | ± | 0.2 | 6.4 | h,i,l | ± | 0.02 | 6.9 | f,g,h,i | ± | 0.05 | 7.0 | e,f | ± | 0.09 | 6.5 | ± | 0.11 | 6.6 | f,g | ± | 0.12 | 5.3 | e,f,g | ± | 0.01 | 4.9 | d,e,f | ± | 0.04 |
| Y2 | 4.9 | ± | 0.2 | 6.6 | l | ± | 0.01 | 7.1 | h,i | ± | 0.01 | 7.2 | f | ± | 0.16 | 6.5 | ± | 0.11 | 6.5 | c,d,e,f | ± | 0.03 | 4.9 | b | ± | 0.07 | 4.6 | c | ± | 0.01 |
| Y20 | 4.9 | ± | 0.2 | 5.5 | d | ± | 0.12 | 5.7 | c | ± | 0.20 | 6.5 | c,d | ± | 0.10 | 6.5 | ± | 0.11 | 6.5 | c,d,e,f,g | ± | 0.01 | 5.1 | b,c,d,e,f | ± | 0.01 | 4.8 | c,d | ± | 0.05 |
| Y21 | 4.9 | ± | 0.2 | 5.3 | c,d | ± | 0.11 | 6.5 | e,f | ± | 0.02 | 7.0 | e,f | ± | 0.04 | 6.5 | ± | 0.11 | 6.6 | d,e,f,g | ± | 0.01 | 5.1 | c,d,e,f | ± | 0.01 | 5.0 | d,e,f,g | ± | 0.01 |
| Y22 | 4.9 | ± | 0.2 | 6.1 | e,f | ± | 0.01 | 6.7 | e,f,g | ± | 0.15 | 7.2 | f | ± | 0.01 | 6.5 | ± | 0.11 | 6.5 | c,d,e | ± | 0.05 | 4.9 | b,c | ± | 0.09 | 4.8 | c,d | ± | 0.01 |
| Y5 | 4.9 | ± | 0.2 | 6.1 | e,f | ± | 0.02 | 6.8 | f,g,h,i | ± | 0.24 | 6.9 | e,f | ± | 0.03 | 6.5 | ± | 0.11 | 6.5 | d,e,f,g | ± | 0.04 | 5.3 | e,f,g | ± | 0.01 | 4.9 | d | ± | 0.03 |
| Y6 | 4.9 | ± | 0.2 | 6.4 | g,h,i | ± | 0.01 | 6.8 | e,f,g,h | ± | 0.15 | 7.1 | f | ± | 0.01 | 6.5 | ± | 0.11 | 6.6 | d,e,f,g | ± | 0.03 | 5.0 | b,c,d | ± | 0.21 | 4.8 | c,d | ± | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gottardi, D.; Siroli, L.; Braschi, G.; Rossi, S.; Bains, N.; Vannini, L.; Patrignani, F.; Lanciotti, R. Selection of Yarrowia lipolytica Strains as Possible Solution to Valorize Untreated Cheese Whey. Fermentation 2023, 9, 51. https://doi.org/10.3390/fermentation9010051
Gottardi D, Siroli L, Braschi G, Rossi S, Bains N, Vannini L, Patrignani F, Lanciotti R. Selection of Yarrowia lipolytica Strains as Possible Solution to Valorize Untreated Cheese Whey. Fermentation. 2023; 9(1):51. https://doi.org/10.3390/fermentation9010051
Chicago/Turabian StyleGottardi, Davide, Lorenzo Siroli, Giacomo Braschi, Samantha Rossi, Narinder Bains, Lucia Vannini, Francesca Patrignani, and Rosalba Lanciotti. 2023. "Selection of Yarrowia lipolytica Strains as Possible Solution to Valorize Untreated Cheese Whey" Fermentation 9, no. 1: 51. https://doi.org/10.3390/fermentation9010051
APA StyleGottardi, D., Siroli, L., Braschi, G., Rossi, S., Bains, N., Vannini, L., Patrignani, F., & Lanciotti, R. (2023). Selection of Yarrowia lipolytica Strains as Possible Solution to Valorize Untreated Cheese Whey. Fermentation, 9(1), 51. https://doi.org/10.3390/fermentation9010051