

Advanced Monitoring and Control of Redox Potential in Wine Fermentation across Scales

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
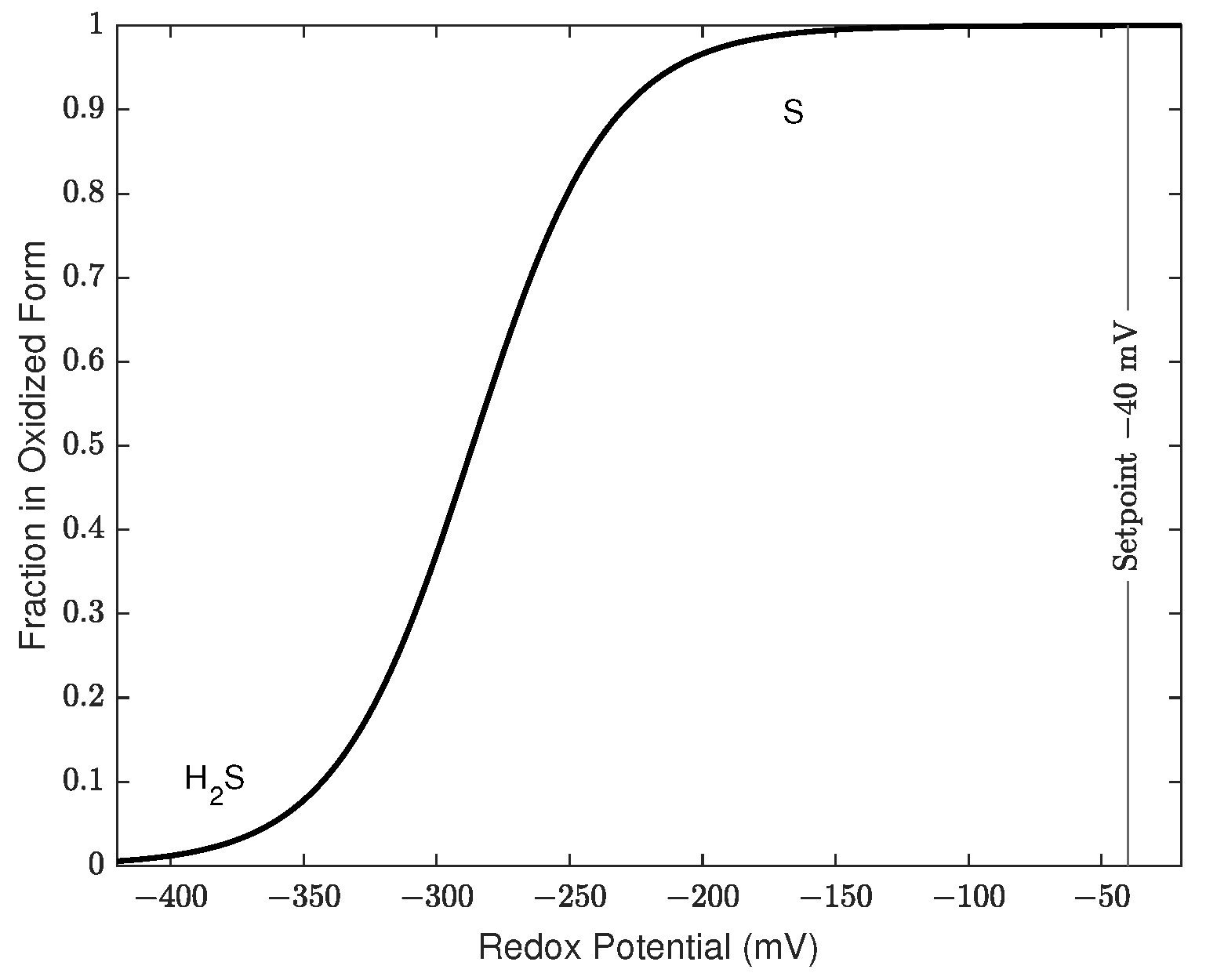
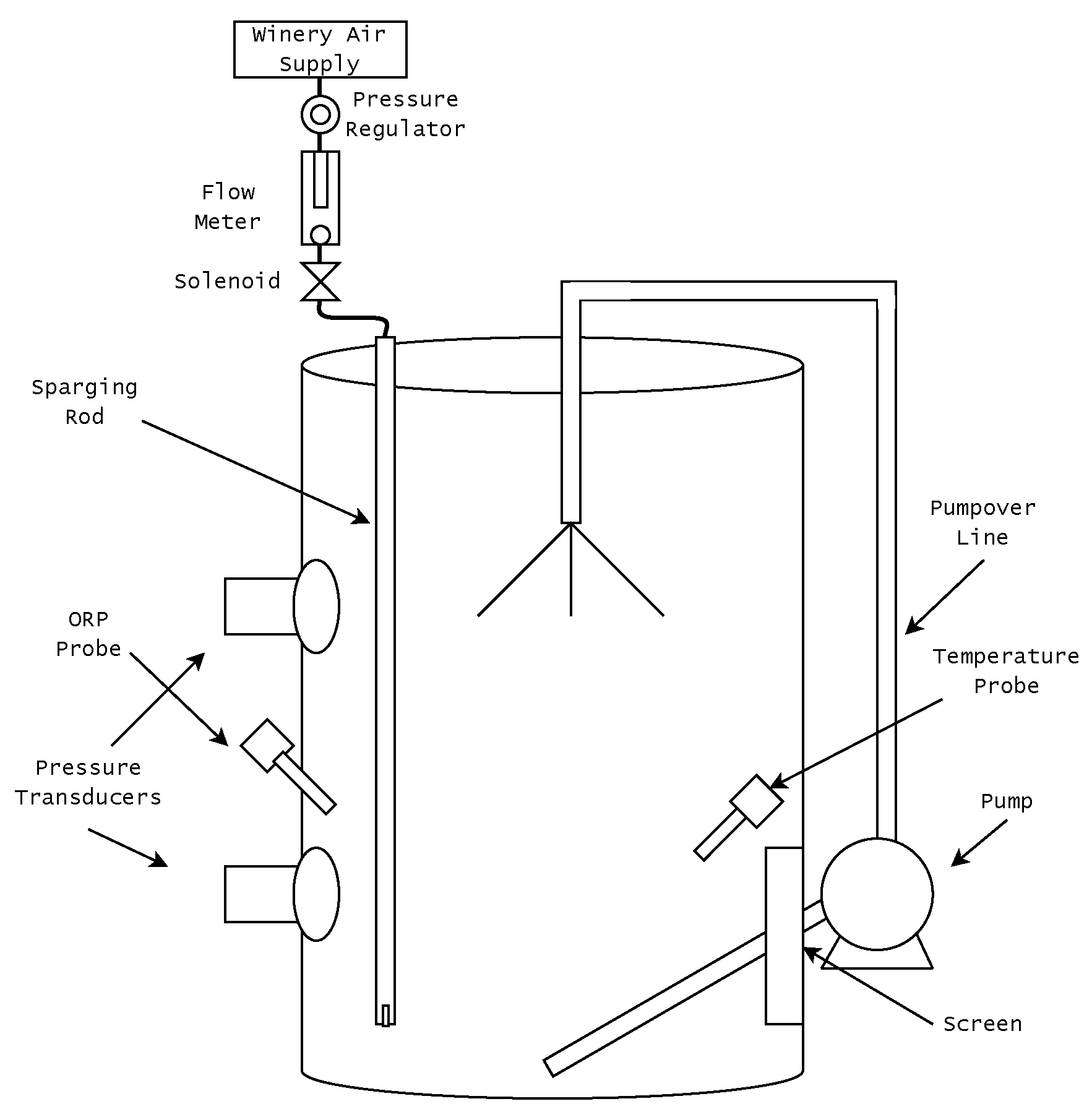
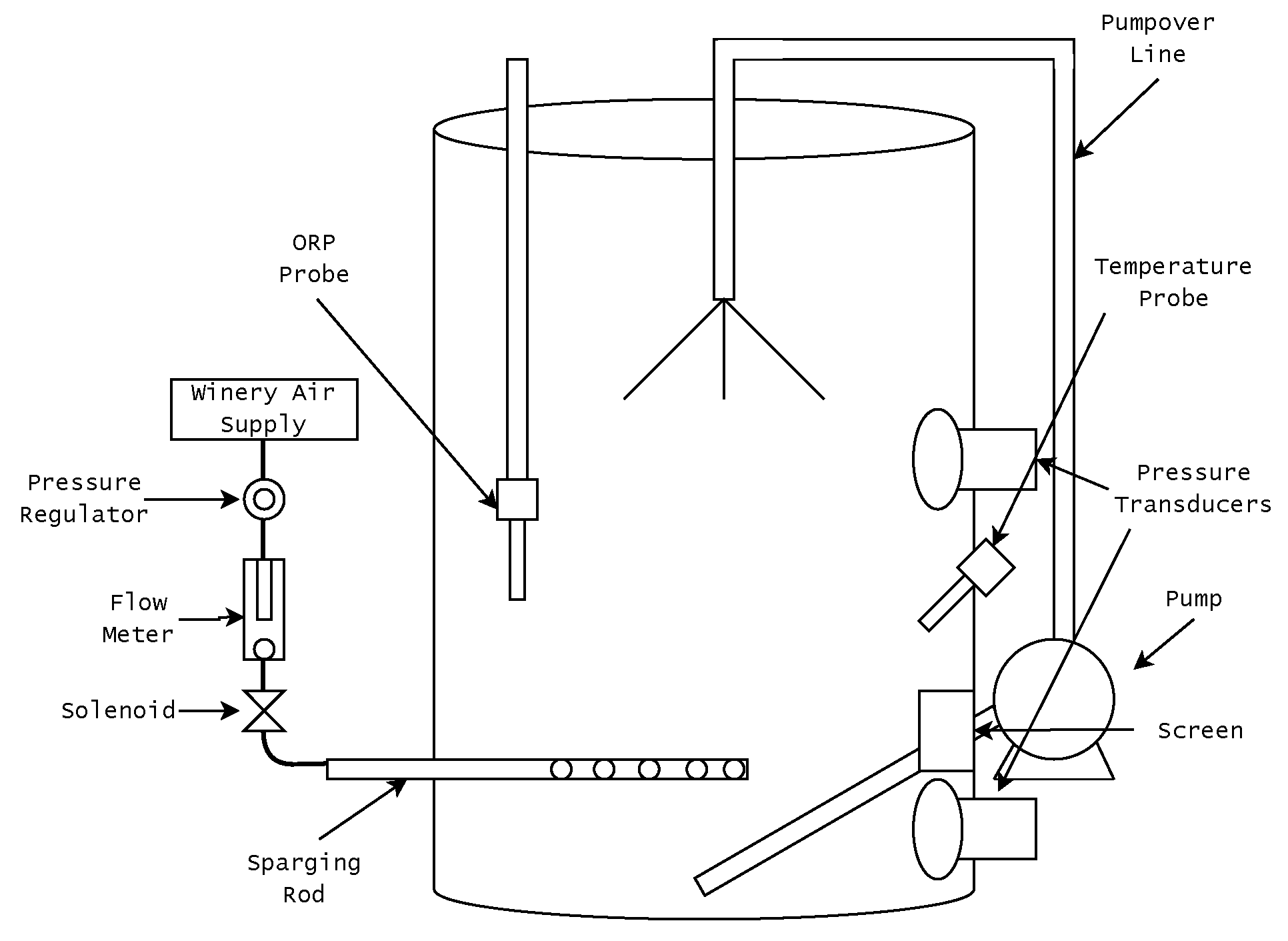
2.1. The Sensing and Control System
2.2. The Wine Fermentations
3. Results
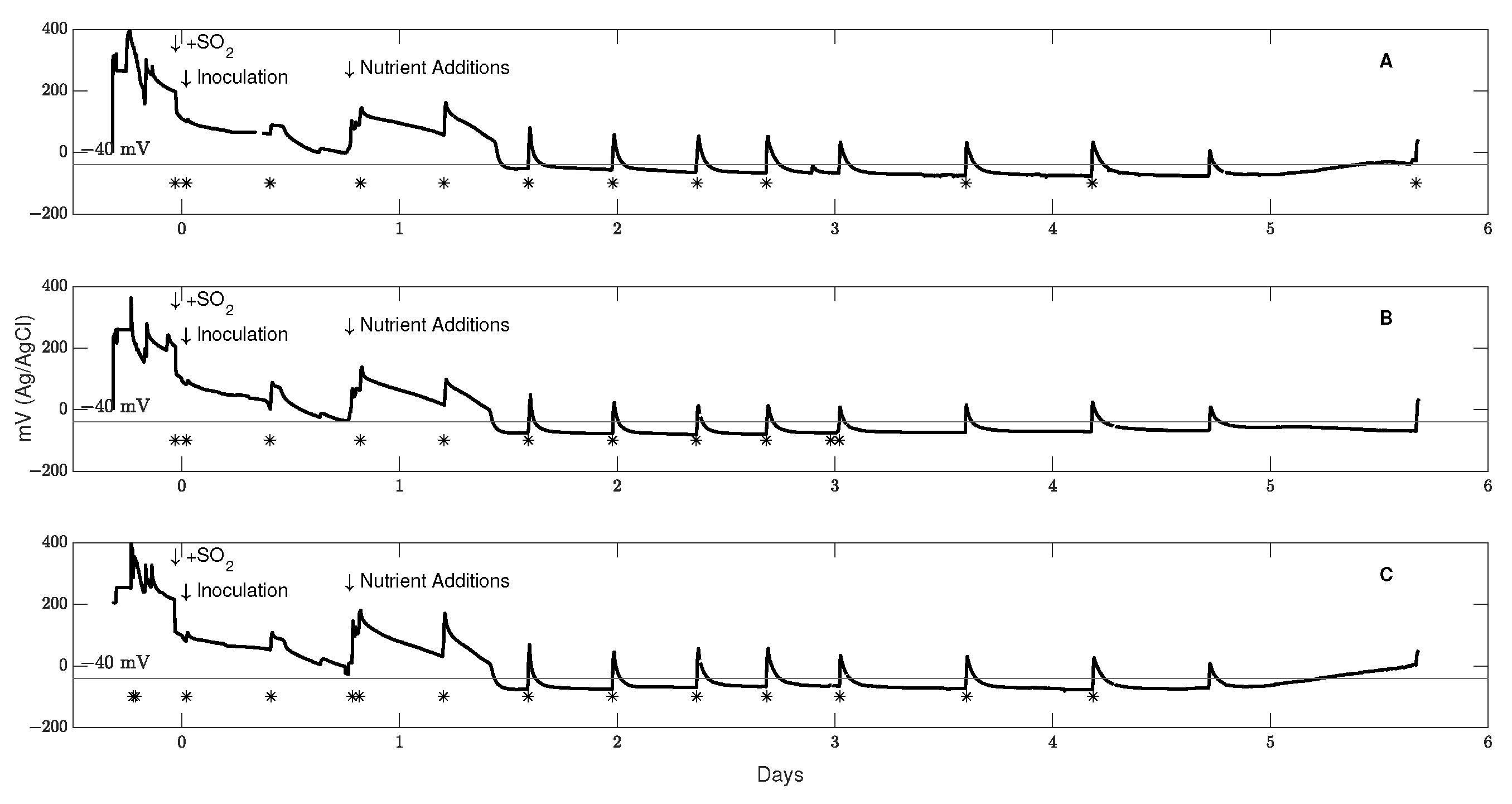
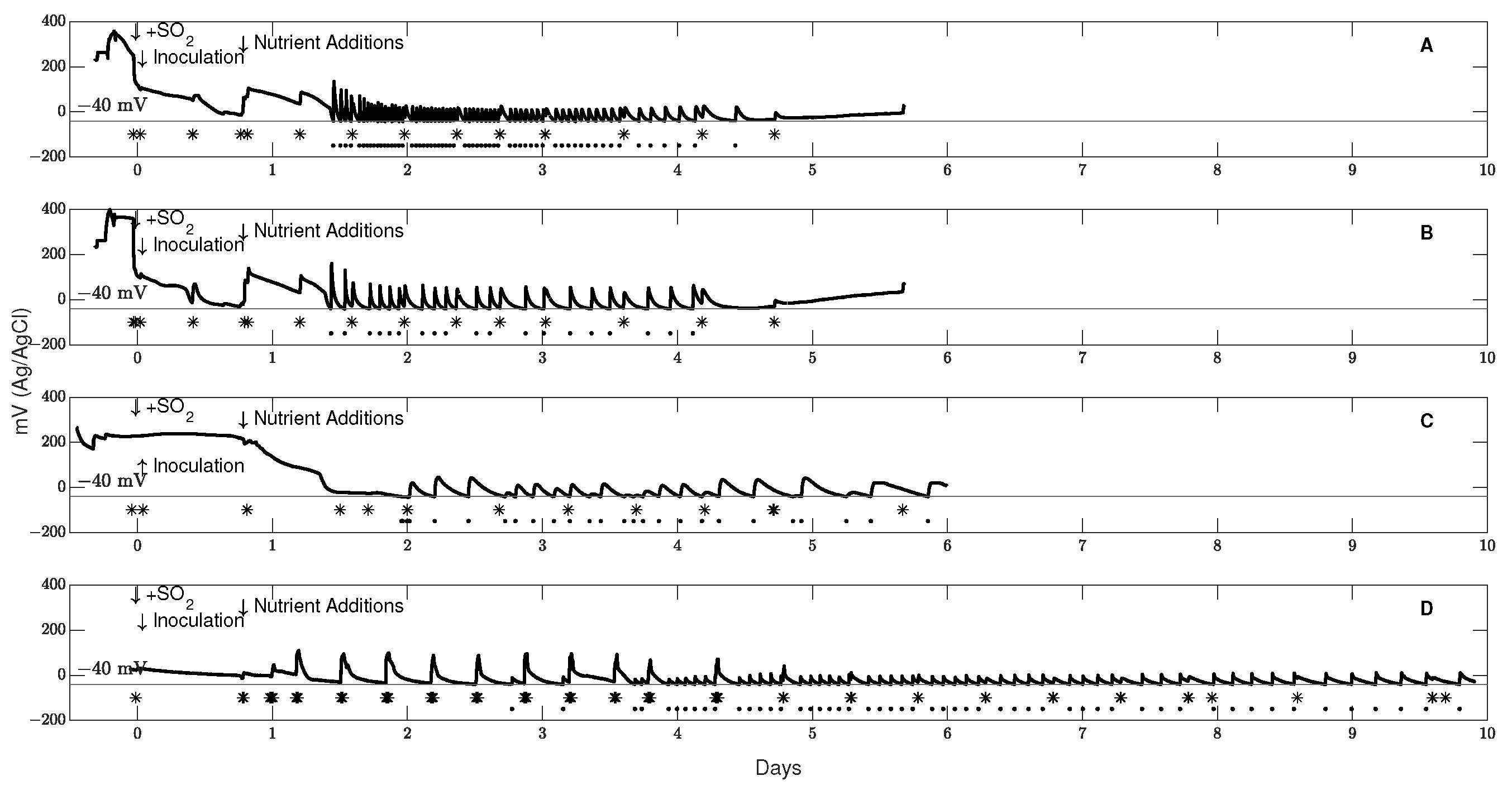
3.1. Control of Redox Potential across Scale
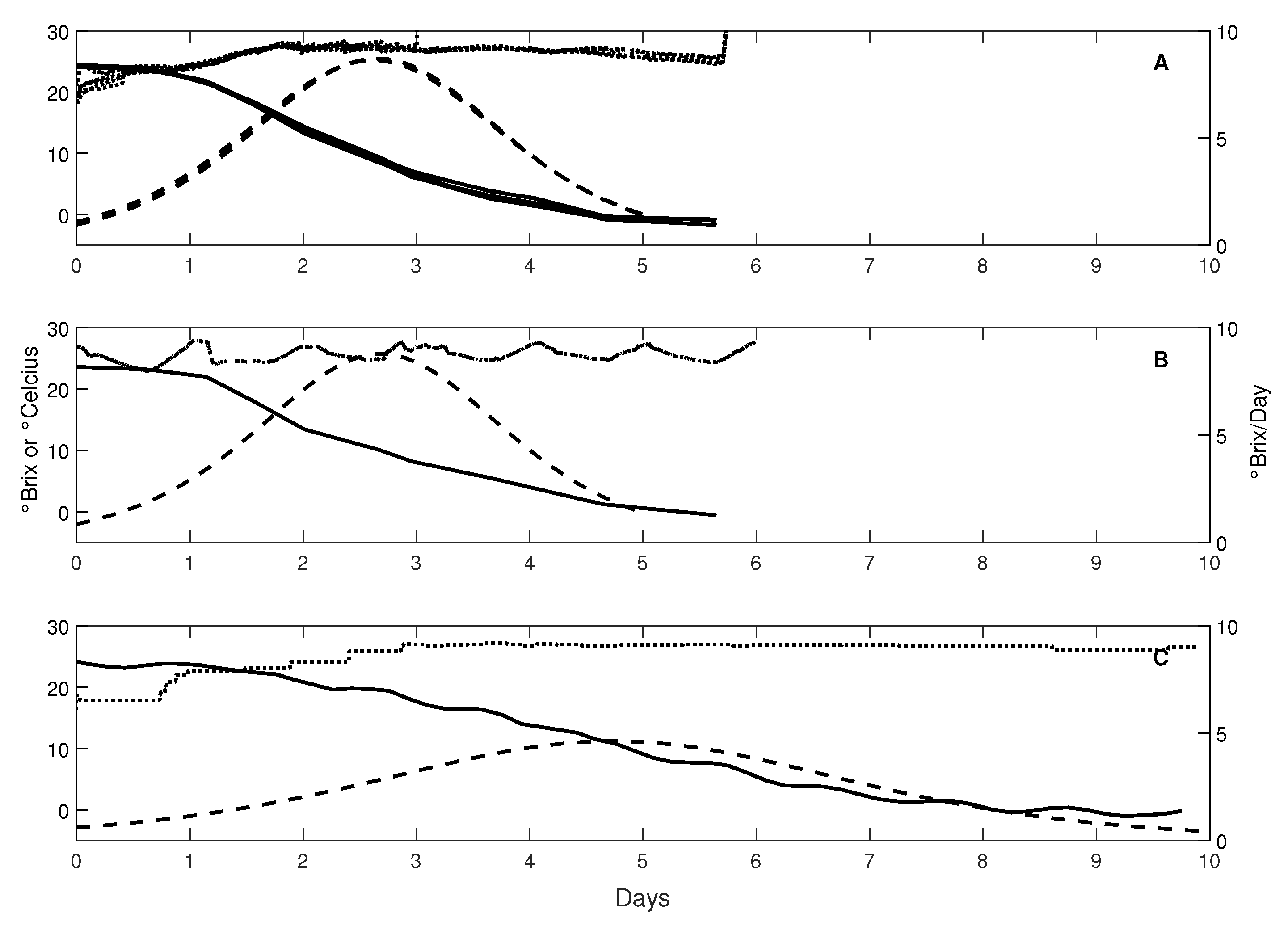
3.2. Fermentation Patterns at Different Scales
3.3. Fermentation Parameter Estimation at Different Scales
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DO | Dissolved Oxygen |
| ORP | Oxidation-reduction Potential |
| PLC | Programmable Logic Controller |
| Ag/AgCl | Silver/Silver Chloride |
| SHE | Standard Hydrogen Electrode |
| PID | Proportional Integral Derivative |
References
- Bisson, L.F. Stuck and Sluggish Fermentations. Am. J. Enol. Vitic. 1999, 50, 107–119. [Google Scholar] [CrossRef]
- Hewitt, L.F. Oxidation-Reduction Potentials in Bacteriology and Biochemistry; London County Council: London, UK, 1931. [Google Scholar]
- Hongo, M.; Ishizaki, A.; Uyeda, M. Studies on oxidation-reduction potentials (ORP) of microbial cultures. Part I. L-glutamic acid fermentation. Agric. Biol. Chem. 1972, 36, 141–145. [Google Scholar]
- Hongo, M.; Uyeda, M. Studies on oxidation-reduction potentials (ORP) of microbial cultures. Part II. L-valine fermentation by Aerobacter aerogenes No 505 and its conversion to lactic acid and alanine formation under anaerobic conditions. Agric. Biol. Chem. 1972, 36, 269–272. [Google Scholar]
- Hongo, M.; Uyeda, M. Studies on oxidation-reduction potentials (ORP) of microbial cultures. Part III. Lactic acid fermentation by Rhizopus G-36 (1). Agric. Biol. Chem. 1972, 36, 273–278. [Google Scholar] [CrossRef]
- Hongo, M.; Uyeda, M. Studies on oxidation-reduction potentials (ORP) of microbial cultures. Part IV. Lactic acid fermentation by Rhizopus G-36 (2). Agric. Biol. Chem. 1972, 36, 279–284. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Chien, W.-S.; Duan, K.-J.; Chang, P.R. Effect of aeration timing and interval during very-high-gravity ethanol fermentation. Process Biochem. 2011, 46, 1025–1028. [Google Scholar] [CrossRef]
- Liu, C.-G.; Wang, N.; Lin, Y.-H.; Bai, F.-W. Very high gravity ethanol fermentation by flocculating yeast under redox potential-controlled conditions. Biotechnol. Biofuels 2012, 5, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Khongsay, N.; Yin, Y.-H.; Laopaiboon, P.; Laopaiboon, L. Improvement of very-high-gravity ethanol fer-mentation from sweet sorghum juice by controlling fermentation redox potential. J. Taiwan Inst. Chem. Eng. 2014, 45, 302–307. [Google Scholar] [CrossRef]
- Goncharuk, V.V.; Bagrii, V.A.; Mel’nik, L.A.; Chebotareva, R.D.; Bashtan, S.Y. The use of redox potential in water treatment processes. J. Water Chem. Technol. 2010, 32, 1–9. [Google Scholar] [CrossRef]
- Brasca, M.; Morandi, S.; Lodi, R.; Tamburini, A. Redox potential to discriminate among species of lactic acid bacteria. J. Appl. Microbiol. 2007, 103, 1516–1624. [Google Scholar] [CrossRef]
- Reichart, O.; Szakmar, K.; Jozwiak, A.; Felfoldi, J.; Baranyai, L. Redox potential measurement as a rapid method for microbiological testing and its validation for coliform determination. Int. J. Food Microbiol. 2007, 114, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Dahod, S.K. Redox potential as a better substitute for dissolved oxygen in fermentation process control. Biotechnol. Bioeng. 1982, 24, 2123–2125. [Google Scholar] [CrossRef]
- Lund, B.M.; Knox, M.R.; Sims, A.P. The effect of oxygen and redox potential on growth of Clostridium botulinum type E from a spore inoculum. Food Microbiol. 1984, 1, 277–287. [Google Scholar] [CrossRef]
- Coleman, R.E.; Boulton, R.B.; Stuchebrukhov, A.A. Kinetics of autoxidation in wine. In Recent Advances in Chemical Kinetics; IntechOpen: London, UK, 2022. [Google Scholar]
- Park, S.K.; Boulton, R.B.; Noble, A.C. Formation of hydrogen sulfide and glutathione during fermentation of white grape musts. Am. J. Enol. Vitic. 2000, 51, 91–97. [Google Scholar] [CrossRef]
- Kukec, A.; Berovic, M.; Celan, S.; Wondra, M. The role of on-line redox potential measurement in Sauvignon blanc fermentation. Food Technol. Biotechnol. 2002, 40, 49–55. [Google Scholar]
- Berovic, M.; Mavri, J.; Wondra, M.; Kosmerl, T.; Bavcar, D. Influence of temperature and carbon dioxide on fermentation of Cabernet Sauvignon must. Food Technol. Biotechnol. 2003, 41, 353–359. [Google Scholar]
- Coleman, R.E. Kinetics of Oxygen Consumption in Solutions of Iron and Tartaric Acid. Ph.D. Thesis, University of California, Davis, CA, USA, 2019. [Google Scholar]
- Peters, R. Oxidation and reduction chains, and the influence of complex ions on the electromotive force. Z. Phys. Chem. Stöchiometrie Verwandtschaftslehre 1898, 26, 193. [Google Scholar] [CrossRef]
- Boulton, R.B.; Singleton, V.L.; Bisson, L.F.; Kunkee, R.E. Principles and Practices of Winemaking; Springer: New York, NY, USA, 1996. [Google Scholar]
- Joslyn, M.A. California wines. Oxidation-reduction potentials at various stages of production and aging. Ind. Eng. Chem. 1949, 41, 587–592. [Google Scholar] [CrossRef]
- Ribereau-Gayon, J. Traite d’Oenologie, Transformations et Traitments des Vins; Libraire Polytechnique Ch. Beranger: Paris, France, 1950. [Google Scholar]
- Schanderl, H. Mikrobiologie des Mostes und Weines; Verlag Eugen Ulmer: Stuttgart, Germany, 1959. [Google Scholar]
- Killeen, D.J.; Boulton, R.B.; Knoesen, A. Advanced monitoring and control of redox potential in wine fermentation. Am. J. Enol. Vitic. 2018, 69, 394–399. [Google Scholar] [CrossRef]
- Murray, D.B.; Haynes, K.; Tomita, M. Redox regulation in respiring Saccharomyces cerevisiae. Biochim. Biophys. 2011, 1810, 945–958. [Google Scholar] [CrossRef]
- Liu, C.-G.; Lin, Y.-H.; Bai, F.-W. Global gene expression analysis of Saccharomyces cerevisiae grown under redox potential-controlled very-high-gravity conditions. Biotechnol. J. 2013, 8, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.-J.; Zhang, J.-R.; Zhang, R.-X.; Qin, Y.; Yang, X.-B.; Jin, G.-J. Oxidation-reduction potential affects medium-chain fatty acid ethyl ester production during wine alcohol fermentation. Food Res. Int. 2022, 157, 11369. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-H.; Chien, W.-S.; Duan, K.-J. Correlations between reduction-oxidation potential profiles and growth patterns of Saccharomyces cerevisiae during very-high-gravity fermentation. Process Biochem. 2010, 45, 765–770. [Google Scholar] [CrossRef]
- Rankine, B.C. Nature, origin and prevention of hydrogen sulphide aroma in wines. J. Sci. Food Agric. 1963, 14, 79–91. [Google Scholar] [CrossRef]
- Schutz, M.; Kunkee, R.E. Formation of hydrogen sulfide from elemental sulfur during fermentation by wine yeast. Am. J. Enol. Vitic. 1977, 28, 137–144. [Google Scholar]
- Siebert, T.E.; Bramley, B.; Solomon, M.R. Hydrogen sulfide: Aroma detection threshold study in white and red wine. AWRI Tech. Rev. 2009, 183, 14–16. [Google Scholar]
- Coleman, R.E.; Boulton, R.B.; Stuchebrukhov, A.A. Kinetics of autoxidation of tartaric acid in the presence of iron. J. Chem. Phys. 2020, 153, 064503. [Google Scholar] [CrossRef]
- Walker, G.A.; Nelson, J.; Halligan, T.; Lima, M.M.M.; Knoesen, A.; Runnebaum, R.C. Monitoring site-specific fermentation outcomes via oxidation reduction potential and uv-vis spectroscopy to characterize “hidden” parameters of pinot noir wine fermentations. Molecules 2021, 26, 4748. [Google Scholar] [CrossRef]
- Lerno, L.; Reichwage, M.; Ponangi, R.; Hearne, L.; Block, D.E.; Oberholster, A. Effects of cap and overall fermentation temperature on phenolic extraction in cabernet sauvignon fermentations. Am. J. Enol. Vitic. 2015, 66, 444–453. [Google Scholar] [CrossRef]
- Nelson, J.; Boulton, R.; Knoesen, A. Automated density measurement with real-time predictive modeling of wine fermentations. IEEE Trans. Instrum. Meas. 2022, 71, 1–7. [Google Scholar] [CrossRef]
- Boulton, R.B. The prediction of fermentation behavior by a kinetic model. Am. J. Enol. Vitic. 1980, 31, 40–44. [Google Scholar]
- Liu, C.-G.; Lin, Y.-H.; Bai, F.-W. Redox potential control and applications in microaerobic and anaerobic fermentations. Biotechnol. Adv. 2013, 31, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yuan, W.; Li, Y.; Bai, F.; Zhong, S.; Jiang, Y. Application of redox potential control to improve ethanol productivity from inulin by consolidated bioprocessing. Process Biochem. 2016, 51, 1544–1551. [Google Scholar] [CrossRef]
- Torija, M.J.; Rozes, N.; Poblet, M.; Guillamon, J.M.; Mas, A. Effects of fermentation temperature on the strain population of Saccharomyces cerevisiae. Int. J. Food Microbiol. 2003, 80, 47–53. [Google Scholar] [CrossRef]
- Ivorra, C.; Perez-Ortin, J.E.; del Olmo, M. An inverse correlation between stress resistance and stuck fer-mentations in wine yeasts. A molecular study. Biotechnol. Bioeng. 1999, 64, 698–708. [Google Scholar] [CrossRef]
- Farina, L.; Medina, K.; Urruty, M.; Biodo, E.; Dellacassa, E.; Carrau, F. Redox effect on volatile compound formation in wine during fermentation by Saccharomyces cerevisiae. Food Chem. 2012, 134, 933–939. [Google Scholar] [CrossRef]
- Bloem, A.; Sanches, I.; Denquin, A.; Camarasa, C. Metabolic impact of redox cofactor perturbations on the formation of aroma compounds in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2016, 82, 174–183. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Values | 100 L | 1,500 L | 10,000 L |
|---|---|---|---|
| Air Pressure (kPa) | 206.8 | 206.8 | 206.8 |
| Air Flow Rate (L/s) | 0.30 | 0.60 | 7.5 |
| Solenoid Open Interval (s) | 20 | 20 | 120 |
| Volume Ratio per Pulse (L air/L wine) | 0.06 | 0.008 | 0.09 |
| Sparger Type | 5-m sparger, 5000 mm2 | 1 hole, 0.79 mm | 24 holes, 0.079 mm |
| Volume | Controlled Redox | Quantity |
|---|---|---|
| 100 L | No | 3 |
| 100 L | Yes | 2 |
| 1500 L | Yes | 1 |
| 10,000 L | Yes | 1 |
| Parameter | 100 L (n = 2) | 1500 L | 10,000 L |
|---|---|---|---|
| Lag Time (h) | 3.78 | 3.33 | 25.1 |
| Initial Nitrogen (mg N/L) | 215 | 198 | 162 |
| Maintenance Rate (1/h) | 0.184 | 0.109 | 0.118 |
| Viability Constant (l/g·h) | 13.8 | 30.8 | 49.2 |
| Ethanol Inhibition (L/g) | 0.035 | 0.044 | 0.037 |
| Sum of Squares Residual per Point (°Brix) | 0.074 | 0.806 | 0.029 |
| Parameter | Uncontrolled Redox Potential (n = 3) | Controlled Redox Potential (n = 2) |
|---|---|---|
| Lag Time (h) | 2.86 ± 1.85 | 3.78 ± 0.48 |
| Initial Nitrogen (mg N/L) | 213 ± 14.9 | 215 ± 28.9 |
| Maintenance Rate (1/h) | 0.144 ± 0.01 | 0.184 ± 0.01 |
| Viability Constant (l/g·h) | 18.7 ± 4.29 | 13.8 ± 0.48 |
| Ethanol Inhibition (L/g) | 0.033 ± 0.009 | 0.035 ± 0.01 |
| Sum of Squares Residual per Point (°Brix) | 0.082 ± 0.02 | 0.074 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nelson, J.; Coleman, R.; Chacón-Rodríguez, L.; Runnebaum, R.; Boulton, R.; Knoesen, A. Advanced Monitoring and Control of Redox Potential in Wine Fermentation across Scales. Fermentation 2023, 9, 7. https://doi.org/10.3390/fermentation9010007
Nelson J, Coleman R, Chacón-Rodríguez L, Runnebaum R, Boulton R, Knoesen A. Advanced Monitoring and Control of Redox Potential in Wine Fermentation across Scales. Fermentation. 2023; 9(1):7. https://doi.org/10.3390/fermentation9010007
Chicago/Turabian StyleNelson, James, Robert Coleman, Leticia Chacón-Rodríguez, Ron Runnebaum, Roger Boulton, and André Knoesen. 2023. "Advanced Monitoring and Control of Redox Potential in Wine Fermentation across Scales" Fermentation 9, no. 1: 7. https://doi.org/10.3390/fermentation9010007
APA StyleNelson, J., Coleman, R., Chacón-Rodríguez, L., Runnebaum, R., Boulton, R., & Knoesen, A. (2023). Advanced Monitoring and Control of Redox Potential in Wine Fermentation across Scales. Fermentation, 9(1), 7. https://doi.org/10.3390/fermentation9010007