
Dark Fermentation in the Dark Biosphere: The Case of Citrobacter sp. T1.2D-12


Abstract
:1. Introduction
2. Materials and Methods
2.1. Methodology for Citrobacter sp. T1.2D-1 Isolation
2.2. DNA Extraction and Sequencing
2.3. Genomic Analysis
2.4. Utilization of Different Electron Donors under Aerobic and Anaerobic Conditions
2.5. Evaluation of the Consumption and Production of Hydrogen
2.6. Nitrate-Dependent Fe2+ Oxidation Assay
2.7. Tetrathionate Reduction Assay
2.8. pH Tolerance Assay
2.9. Heavy Metal-Resistance Assay
3. Results and Discussion
3.1. Genomic Analysis
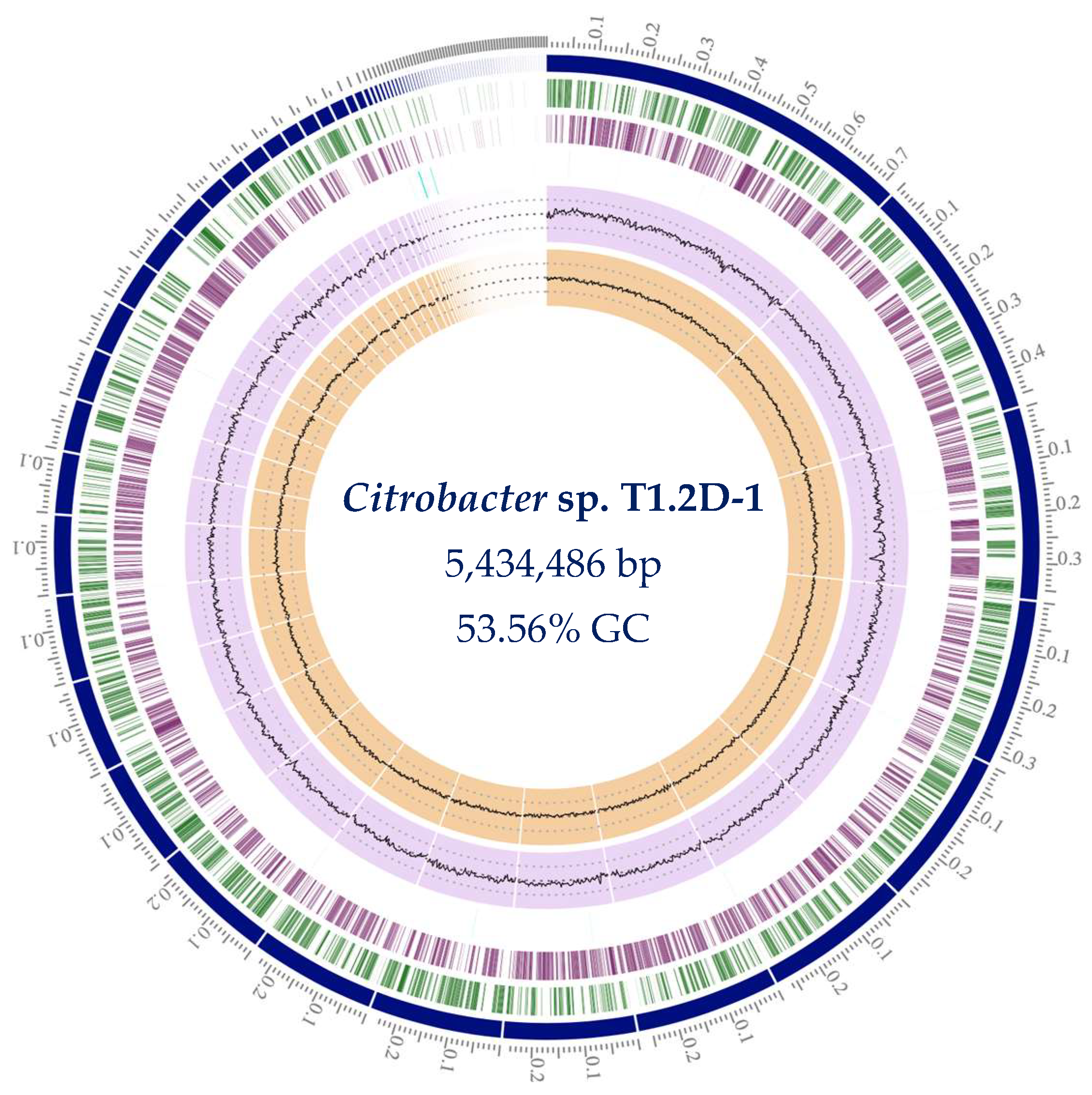
3.1.1. Genome Assembly and Annotation
3.1.2. Phylogenomic Analysis
3.2. Carbon Sources Utilization
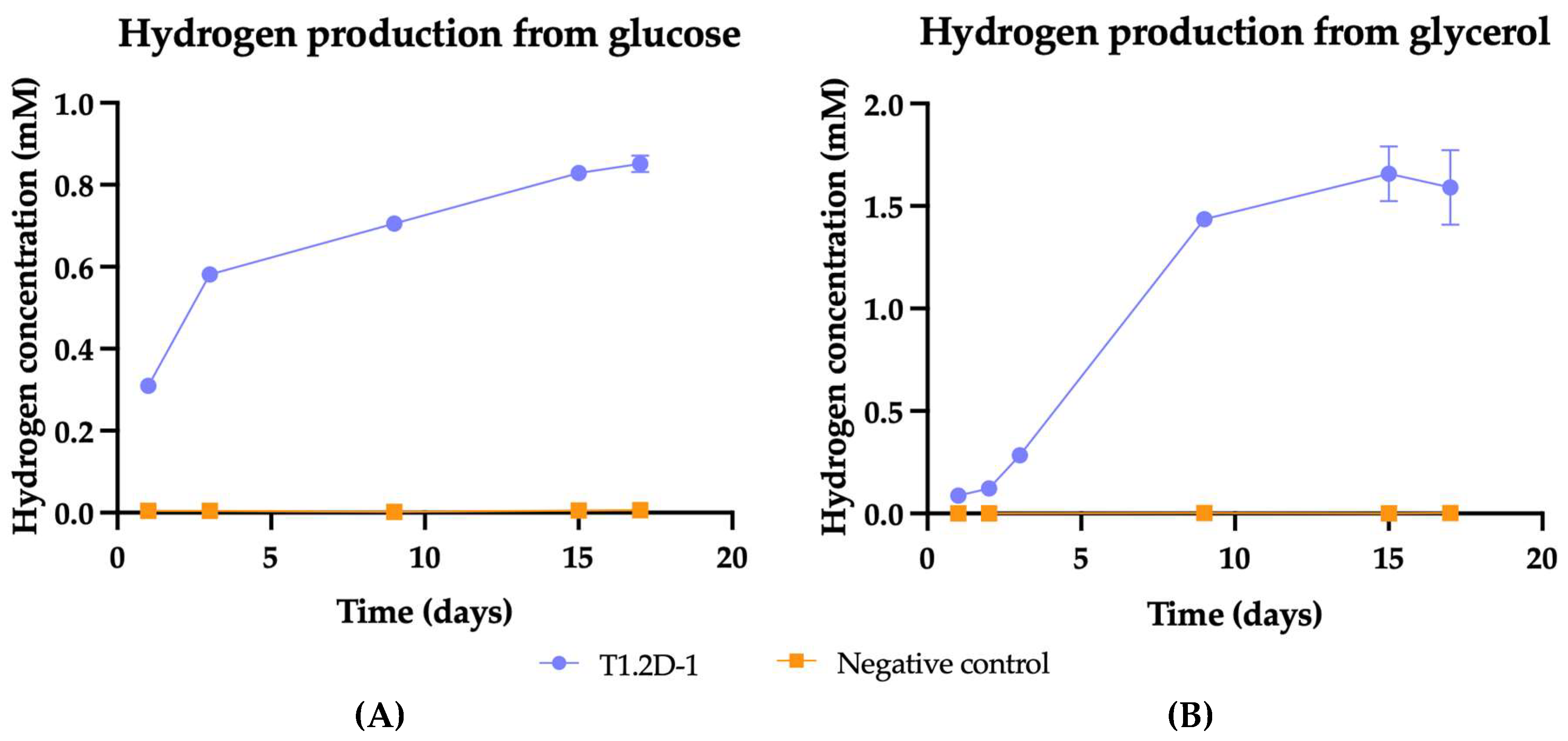
Hydrogen Production
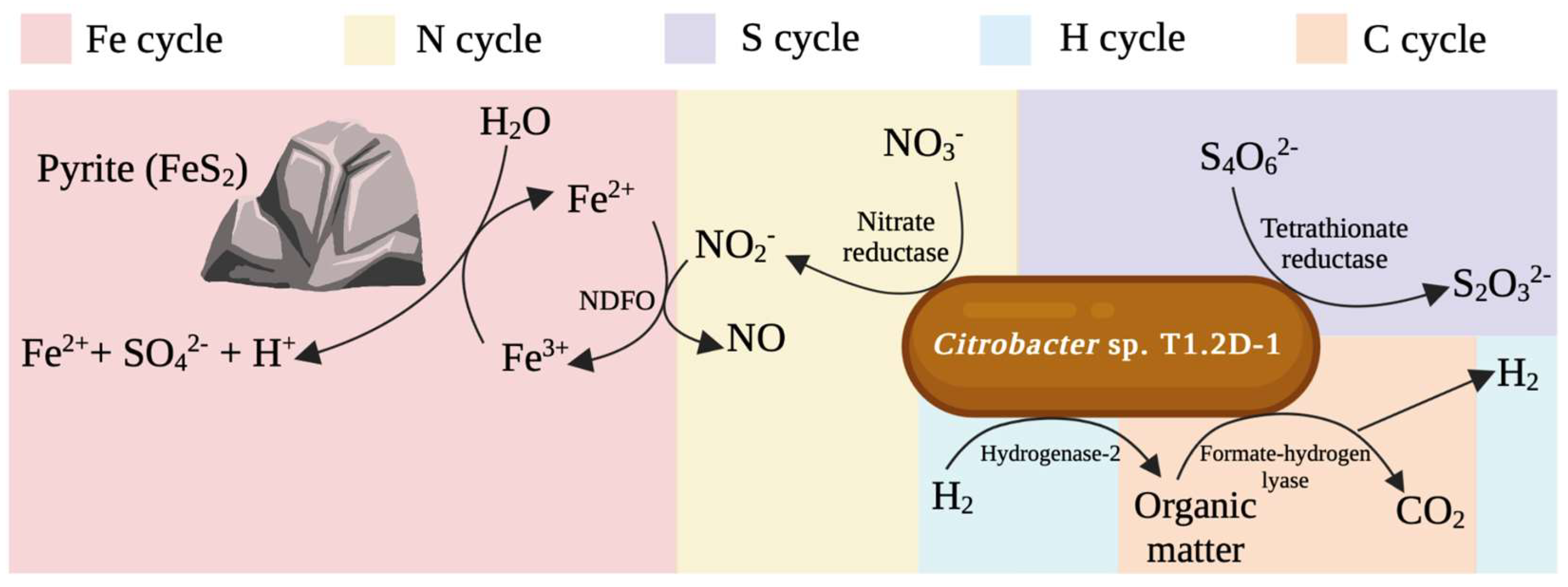
3.3. Other Metabolisms of Interest
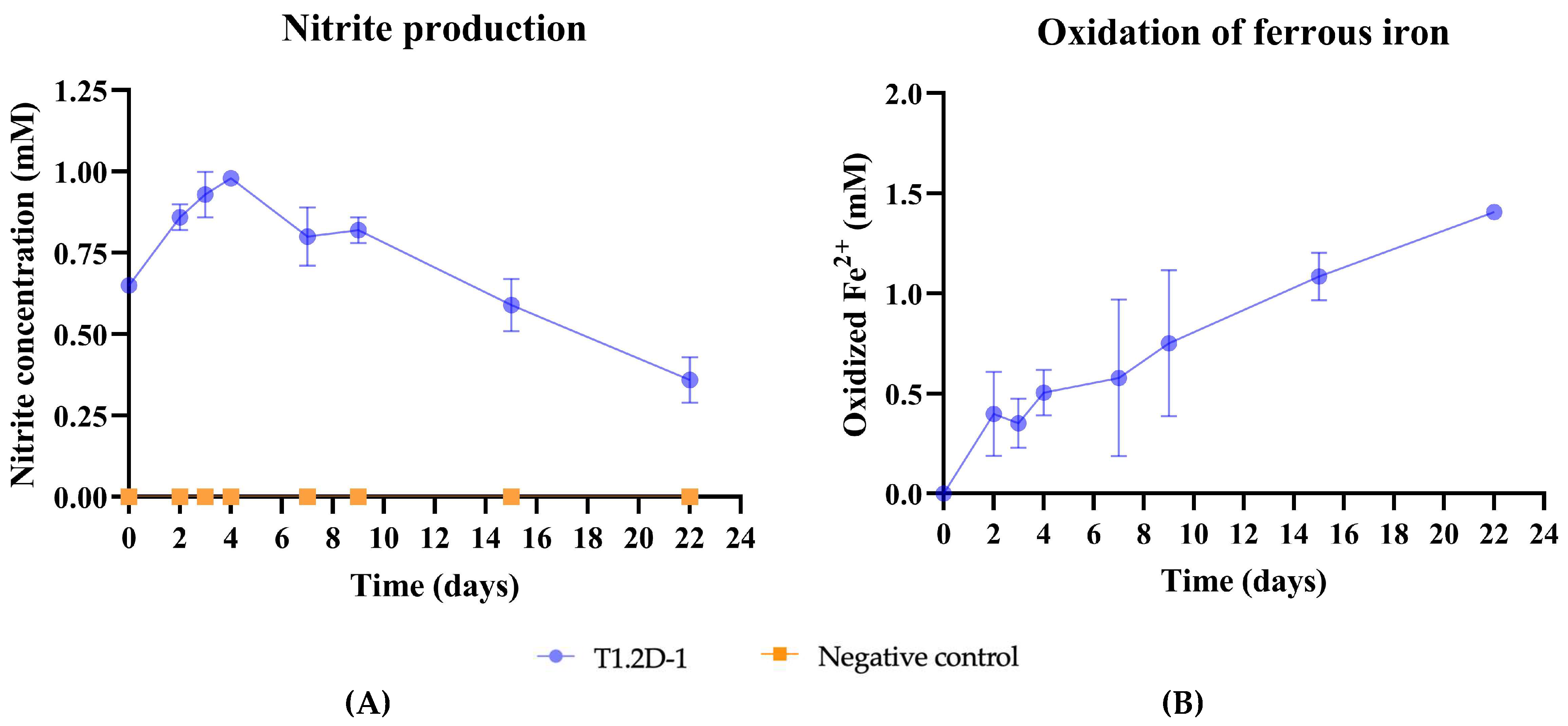
3.3.1. Nitrate-Dependent Fe2+ Oxidation Assay
3.3.2. Tetrathionate Reduction Assay
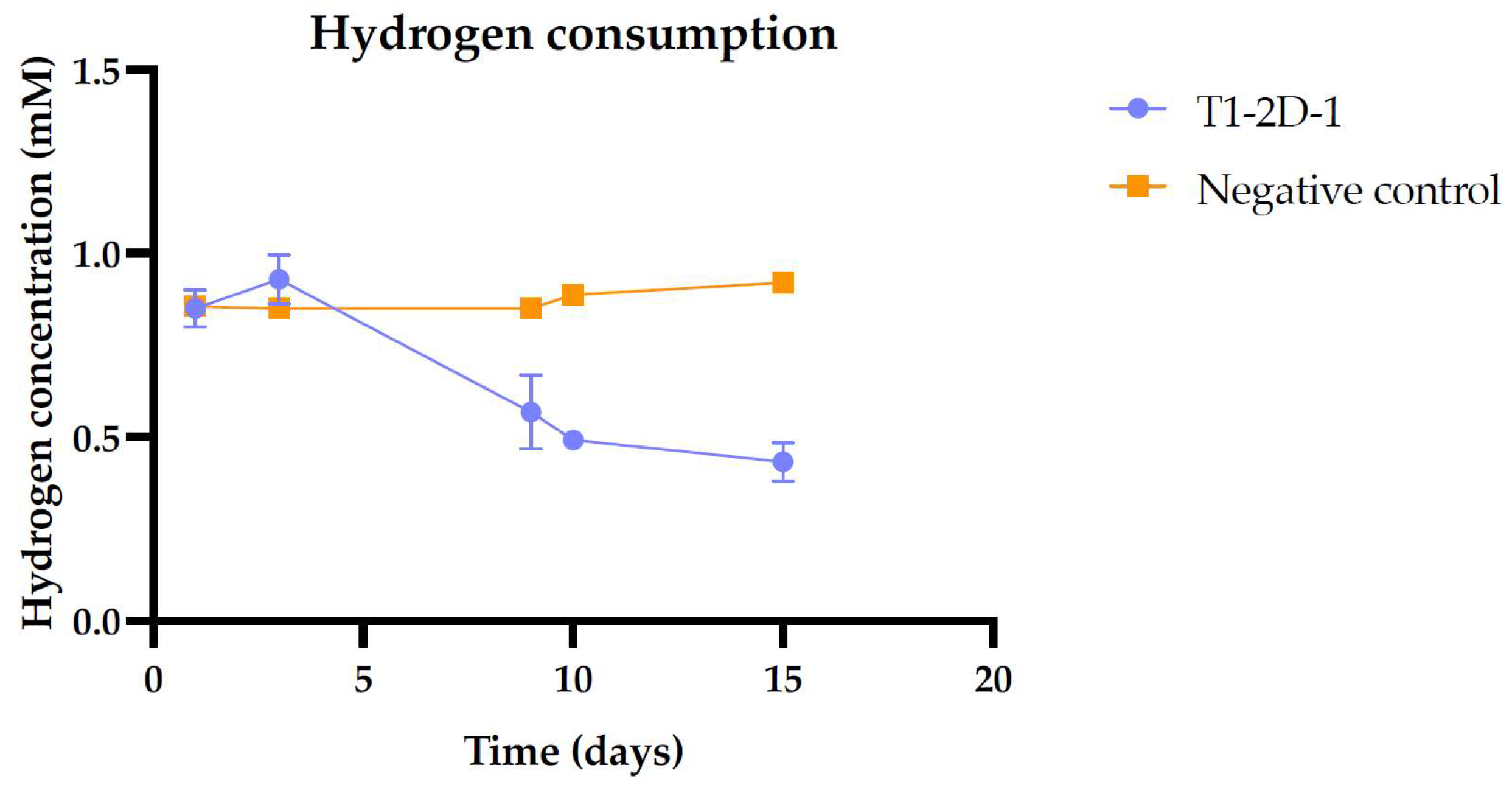
3.3.3. Hydrogen Consumption Assay
3.3.4. pH Tolerance Assay
3.3.5. Heavy Metal Tolerance Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wong, Y.M.; Wu, T.Y.; Juan, J.C. A review of sustainable hydrogen production using seed sludge via dark fermentation. Renew. Sustain. Energy Rev. 2014, 34, 471–482. [Google Scholar] [CrossRef]
- Otaki, H.; Everroad, R.C.; Matsuura, K.; Haruta, S. Production and consumption of hydrogen in hot spring microbial mats dominated by a filamentous anoxygenic photosynthetic bacterium. Microb. Environ. 2012, 27, 293–299. [Google Scholar] [CrossRef]
- Hylemon, P.B.; Harris, S.C.; Ridlon, J.M. Metabolism of hydrogen gases and bile acids in the gut microbiome. FEBS Lett. 2018, 592, 2070–2082. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Zhang, X.; Zhao, G.; Zhang, X.; Dong, L.; Chen, Y. Aerobic hydrogen-oxidizing bacteria in soil: From cells to ecosystems. Rev. Environ. Sci. Bio/Technol. 2022, 21, 877–904. [Google Scholar] [CrossRef]
- Ruff, S.E.; Humez, P.; de Angelis, I.H.; Diao, M.; Nightingale, M.; Cho, S.; Connors, L.; Kuloyo, O.O.; Seltzer, A.; Bowman, S. Hydrogen and dark oxygen drive microbial productivity in diverse groundwater ecosystems. Nat. Commun. 2023, 14, 3194. [Google Scholar] [CrossRef]
- Kieft, T.L. Microbiology of the Deep Continental Biosphere. In Advances in Environmental Microbiology; Springer: Berlin/Heidelberg, Germany, 2016; pp. 225–249. [Google Scholar] [CrossRef]
- Escudero, C.; Oggerin, M.; Amils, R. The deep continental subsurface: The dark biosphere. Int. Microbiol. 2018, 21, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Dombard, D.A.R.; Malas, J. Advances in defining ecosystem functions of the terrestrial subsurface biosphere. Front. Microbiol. 2022, 13, 891528. [Google Scholar] [CrossRef] [PubMed]
- Stevens, T.O.; McKinley, J.P. Lithoautotrophic microbial ecosystems in deep basalt aquifers. Science 1995, 270, 450–455. [Google Scholar] [CrossRef]
- Pedersen, K. Exploration of deep intraterrestrial microbial life: Current perspectives. FEMS Microbiol. Lett. 2000, 185, 9–16. [Google Scholar] [CrossRef]
- Amils, R.; Escudero, C.; Oggerin, M.; Puente Sánchez, F.; Arce Rodríguez, A.; Fernández Remolar, D.; Rodríguez, N.; García Villadangos, M.; Sanz, J.L.; Briones, C.; et al. Coupled C, H, N, S and Fe biogeochemical cycles operating in the continental deep subsurface of the Iberian Pyrite Belt. Environ. Microbiol. 2022, 25, 428–453. [Google Scholar] [CrossRef]
- Wu, X.; Pedersen, K.; Edlund, J.; Eriksson, L.; Åström, M.; Andersson, A.F.; Bertilsson, S.; Dopson, M. Potential for hydrogen-oxidizing chemolithoautotrophic and diazotrophic populations to initiate biofilm formation in oligotrophic, deep terrestrial subsurface waters. Microbiome 2017, 5, 37. [Google Scholar] [CrossRef] [PubMed]
- Sanz, J.L.; Rodriguez, N.; Escudero, C.; Carrizo, D.; Amils, R. Biological production of H2, CH4 and CO2 in the deep subsurface of the Iberian Pyrite Belt. Environ. Microbiol. 2021, 23, 3913–3922. [Google Scholar] [CrossRef] [PubMed]
- Tornos, F. Environment of formation and styles of volcanogenic massive sulfides: The Iberian Pyrite Belt. Ore Geol. Rev. 2006, 28, 259–307. [Google Scholar] [CrossRef]
- Amils, R.; González-Toril, E.; Fernández-Remolar, D.; Gómez, F.; Rodríguez, N.; Durán, C. Interaction of the sulfur and iron cycles in the Tinto River ecosystem. Rev. Environ. Sci. Biotechnol. 2002, 1, 299–309. [Google Scholar] [CrossRef]
- Amils, R.; Fernández-Remolar, D. IPBSL Team Río Tinto: A geochemical and mineralogical terrestrial analogue of Mars. Life 2014, 4, 511–534. [Google Scholar] [CrossRef] [PubMed]
- Bryce, C.; Blackwell, N.; Schmidt, C.; Otte, J.; Huang, Y.; Kleindienst, S.; Tomaszewski, E.; Schad, M.; Warter, V.; Peng, C. Microbial anaerobic Fe(II) oxidation–ecology, mechanisms and environmental implications. Environ. Microbiol. 2018, 20, 3462–3483. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.J.; Goebel, B.M.; Rodgers, T.M.; Schrenk, M.O.; Gihring, T.M.; Cardona, M.M.; Mcguire, M.M.; Hamers, R.J.; Pace, N.R.; Banfield, J.F. Geomicrobiology of pyrite (FeS2) dissolution: Case study at Iron Mountain, California. Geomicrobiol. J. 1999, 16, 155–179. [Google Scholar]
- Schippers, A.; Jørgensen, B.B. Biogeochemistry of pyrite and iron sulfide oxidation in marine sediments. Geochim. Cosmochim. Acta 2002, 66, 85–92. [Google Scholar] [CrossRef]
- Juncher Jørgensen, C.; Jacobsen, O.S.; Elberling, B.; Aamand, J. Microbial oxidation of pyrite coupled to nitrate reduction in anoxic groundwater sediment. Environ. Sci. Technol. 2009, 43, 4851–4857. [Google Scholar] [CrossRef]
- Leandro, T. Microbial Diversity and Anaerobic Metabolisms in the Subsurface of The Iberian Pyritic Belt. Ph.D. Dissertation; Universidad de Coimbra: Coimbra, Portugal, 2018. Available online: https://cab.inta-csic.es/en/tesis/microbial-diversity-and-anaerobic-metabolisms-in-the-subsurface-of-the-iberian-pyritic-belt/ (accessed on 1 January 2018).
- Borenshtein, D.; Schauer, D. The genus Citrobacter. Prokaryotes 2006, 6, 90–98. [Google Scholar]
- Furlan, J.P.R.; Ribeiro, T.G.; Novais, Â.; Cabral, H.; Peixe, L.; Stehling, E.G. Citrobacter meridianamericanus sp. nov., isolated from a soil sample. Int. J. Syst. Evol. Microbiol. 2023, 73, 005720. [Google Scholar] [CrossRef]
- Wang, H.; Hou, H.; Huang, J. Citrobacter arsenatis sp. nov., an arsenate-reducing bacterium isolated from freshwater sediment. Antonie Van Leeuwenhoek 2021, 114, 1285–1292. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, T.G.; Goncalves, B.R.; da Silva, M.S.; Novais, A.; Machado, E.; Carrico, J.A.; Peixe, L. Citrobacter portucalensis sp. nov., isolated from an aquatic sample. Int. J. Syst. Evol. Microbiol. 2017, 67, 3513–3517. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Cui, X.; Liu, W.; Xu, H.; Zheng, B. Genetic characterization of a novel sequence type of multidrug-resistant Citrobacter freundii strain recovered from wastewater treatment plant. Infect. Drug Res. 2019, 12, 2775–2779. [Google Scholar] [CrossRef] [PubMed]
- Metsoviti, M.; Zeng, A.; Koutinas, A.A.; Papanikolaou, S. Enhanced 1, 3-propanediol production by a newly isolated Citrobacter freundii strain cultivated on biodiesel-derived waste glycerol through sterile and non-sterile bioprocesses. J. Biotechnol. 2013, 163, 408–418. [Google Scholar] [CrossRef]
- Unuofin, J.O.; Moubasher, H.A.; Okoh, A.I.; Nwodo, U.U. Production of polyextremotolerant laccase by Achromobacter xylosoxidans HWN16 and Citrobacter freundii LLJ16. Biotechnol. Rep. 2019, 22, e00337. [Google Scholar] [CrossRef]
- Amils, R.; Fernández-Remolar, D.; Parro, V.; Rodríguez-Manfredi, J.A.; Timmis, K.; Oggerin, M.; Sánchez-Román, M.; López, F.J.; Fernández, J.P.; Puente, F.; et al. Iberian Pyrite Belt Subsurface Life (IPBSL), a drilling project of biohydrometallurgical interest. Adv. Mater. Res. 2013, 825, 15–18. [Google Scholar]
- Sanz, J.L.; Rodríguez, N.; Amils, R. Effect of chlorinated aliphatic hydrocarbons on the acetoclastic methanogenic activity of granular sludge. Appl. Microbiol. Biotechnol. 1997, 47, 324–328. [Google Scholar] [CrossRef]
- Leandro, T.; Rodriguez, N.; Rojas, P.; Sanz, J.L.; Da Costa, M.S.; Amils, R. Study of methanogenic enrichment cultures of rock cores from the deep subsurface of the Iberian Pyritic Belt. Heliyon 2018, 4, e00605. [Google Scholar] [CrossRef]
- Kate, W. Preparation of genomic DNA from bacteria. Curr. Protoc. Mol. Biol. 2001, 56, 2–4. [Google Scholar] [CrossRef]
- Olson, R.D.; Assaf, R.; Brettin, T.; Conrad, N.; Cucinell, C.; Davis, J.J.; Dempsey, D.M.; Dickerman, A.; Dietrich, E.M.; Kenyon, R.W. Introducing the Bacterial and Viral Bioinformatics Resource Center (BV-BRC): A resource combining PATRIC, IRD and ViPR. Nucleic Acids Res. 2023, 51, 678–689. [Google Scholar] [CrossRef]
- The Galaxy Community. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2022 update. Nucleic Acids Res. 2022, 50, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Antipov, D.; Hartwick, N.; Shen, M.; Raiko, M.; Lapidus, A.; Pevzner, P.A. plasmidSPAdes: Assembling plasmids from whole genome sequencing data. Bioinformatics 2016, 32, 3380–3387. [Google Scholar] [CrossRef] [PubMed]
- Boetzer, M.; Henkel, C.V.; Jansen, H.J.; Butler, D.; Pirovano, W. Scaffolding pre-assembled contigs using SSPACE. Bioinformatics 2011, 27, 578–579. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2022, 50, 801–807. [Google Scholar] [CrossRef]
- Rodriguez-R, L.M.; Gunturu, S.; Harvey, W.T.; Rosselló-Mora, R.; Tiedje, J.M.; Cole, J.R.; Konstantinidis, K.T. The Microbial Genomes Atlas (MiGA) webserver: Taxonomic and gene diversity analysis of Archaea and Bacteria at the whole genome level. Nucleic Acids Res. 2018, 46, 282–288. [Google Scholar] [CrossRef]
- Han, M.V.; Zmasek, C.M. phyloXML: XML for evolutionary biology and comparative genomics. BMC Bioinform. 2009, 10, 356. [Google Scholar] [CrossRef]
- Zhou, G.; Li, L.; Shi, Q.; Ouyang, Y.; Chen, Y.; Hu, W. Effects of nutritional and environmental conditions on planktonic growth and biofilm formation of Citrobacter werkmanii BF-6. J. Microbiol. Biotechnol. 2013, 23, 1673–1682. [Google Scholar] [CrossRef]
- Mateos, G.; Bonilla, A.M.; de Francisco de Polanco, S.; Martínez, J.M.; Escudero, C.; Rodríguez, N.; Sánchez-Andrea, I.; Amils, R. Shewanella sp. T2. 3D-1.1 a Novel Microorganism Sustaining the Iron Cycle in the Deep Subsurface of the Iberian Pyrite Belt. Microorganisms 2022, 10, 1585. [Google Scholar] [CrossRef] [PubMed]
- Bricio, C. Producción de óxidos de Nitrógeno Gaseosos en Thermus Thermophilus. Ph.D. Dissertation; Universidad Autónoma de Madrid: Madrid, Spain, 2012. Available online: http://hdl.handle.net/10486/11954 (accessed on 26 October 2012).
- Novotný, Č Instability of the tetrathionate respiratory chain of Citrobacter freundii. Folia Microbiol. 1978, 23, 428–432. [CrossRef] [PubMed]
- Kelly, D.P.; Chambers, L.A.; Trudinger, P.A. Cyanolysis and spectrophotometric estimation of trithionate in mixture with thiosulfate and tetrathionate. Anal. Chem. 1969, 41, 898–901. [Google Scholar] [CrossRef]
- Yuan, C.; Yin, Z.; Wang, J.; Qian, C.; Wei, Y.; Zhang, S.; Jiang, L.; Liu, B. Comparative genomic analysis of Citrobacter and key genes essential for the pathogenicity of Citrobacter koseri. Front. Microbiol. 2019, 10, 2774. [Google Scholar] [CrossRef] [PubMed]
- Sinha, P.; Roy, S.; Das, D. Role of formate hydrogen lyase complex in hydrogen production in facultative anaerobes. Int. J. Hydrogen Energy 2015, 40, 8806–8815. [Google Scholar] [CrossRef]
- Kim, S.; Seol, E.; Raj, S.M.; Park, S.; Oh, Y.; Ryu, D.D. Various hydrogenases and formate-dependent hydrogen production in Citrobacter amalonaticus Y19. Int. J. Hydrogen Energy 2008, 33, 1509–1515. [Google Scholar] [CrossRef]
- Evans, R.M.; Beaton, S.E. A novel overproduction system for the structural determination of a proton-pumping hydrogen-producing [NiFe]-hydrogenase. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2018; Volume 613, pp. 91–116. [Google Scholar]
- Poblete-Castro, I.; Wittmann, C.; Nikel, P.I. Biochemistry, genetics and biotechnology of glycerol utilization in Pseudomonas species. Microb. Biotechnol. 2020, 13, 32–53. [Google Scholar] [CrossRef]
- Clomburg, J.M.; Gonzalez, R. Anaerobic fermentation of glycerol: A platform for renewable fuels and chemicals. Trends Biotechnol. 2013, 31, 20–28. [Google Scholar] [CrossRef]
- Ju, J.; Wang, D.; Heo, S.; Kim, M.; Seo, J.; Kim, Y.; Kim, D.; Kang, S.; Kim, C.; Oh, B. Enhancement of 1, 3-propanediol production from industrial by-product by Lactobacillus reuteri CH53. Microb. Cell Fact. 2020, 19, 6. [Google Scholar] [CrossRef]
- Lee, J.H.; Jung, M.; Oh, M. High-yield production of 1, 3-propanediol from glycerol by metabolically engineered Klebsiella pneumoniae. Biotechnol. Biofuels 2018, 11, 104. [Google Scholar] [CrossRef]
- Tao, Y.; Bu, C.; Zou, L.; Hu, Y.; Zheng, Z.; Ouyang, J. A comprehensive review on microbial production of 1, 2-propanediol: Micro-organisms, metabolic pathways, and metabolic engineering. Biotechnol. Biofuels 2021, 14, 216. [Google Scholar] [CrossRef] [PubMed]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [PubMed]
- Konstantinidis, K.T.; Tiedje, J.M. Towards a genome-based taxonomy for prokaryotes. J. Bacteriol. 2005, 187, 6258–6264. [Google Scholar] [CrossRef] [PubMed]
- Yazdani, S.S.; Gonzalez, R. Anaerobic fermentation of glycerol: A path to economic viability for the biofuels industry. Curr. Opin. Biotechnol. 2007, 18, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.; Wu, J.; Huang, R.; Zhang, W.; He, W.; Deng, Z.; Han, Y.; Xiao, B.; Luo, H.; Qu, W. Effects of Temperature and Total Solid Content on Biohydrogen Production from Dark Fermentation of Rice Straw: Performance and Microbial Community Characteristics. Chemosphere 2022, 286, 131655. [Google Scholar] [CrossRef]
- Corsino, S.F.; Di Trapani, D.; Capodici, M.; Torregrossa, M.; Viviani, G. Optimization of acetate production from citrus wastewater fermentation. Water Resour. Ind. 2021, 25, 100140. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, C.; Lu, Y.; Wu, X.; Wang, L.; Wang, L.; Wang, L.; Han, B.; Xing, X.H. States and challenges for high-value biohythane production from waste biomass by dark fermentation technology. Bioresour. Technol. 2013, 135, 292–303. [Google Scholar] [CrossRef]
- Abedi, E.; Hashemi, S.M.B. Lactic Acid Production—Producing Microorganisms and Substrates Sources-State of Art. Heliyon 2020, 6, 04974. [Google Scholar] [CrossRef]
- Tse, T.J.; Wiens, D.J.; Reaney, M.J. Production of bioethanol—A review of factors affecting ethanol yield. Fermentation 2021, 7, 268. [Google Scholar] [CrossRef]
- Patel, S.K.; Kumar, P.; Mehariya, S.; Purohit, H.J.; Lee, J.; Kalia, V.C. Enhancement in hydrogen production by co-cultures of Bacillus and Enterobacter. Int. J. Hydrogen Energy 2014, 39, 14663–14668. [Google Scholar] [CrossRef]
- Cabrol, L.; Marone, A.; Tapia-Venegas, E.; Steyer, J.; Ruiz-Filippi, G.; Trably, E. Microbial ecology of fermentative hydrogen producing bioprocesses: Useful insights for driving the ecosystem function. FEMS Microbiol. Rev. 2017, 41, 158–181. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Pan, X.; Zhang, D.; Lee, D.; Al-Misned, F.A.; Mortuza, M.G. Anaerobic nitrate reduction with oxidation of Fe(II) by Citrobacter freundii strain PXL1—A potential candidate for simultaneous removal of As and nitrate from groundwater. Ecol. Eng. 2015, 77, 196–201. [Google Scholar] [CrossRef]
- Escudero, C.; Del Campo, A.; Ares, J.R.; Sánchez, C.; Martínez, J.M.; Gómez, F.; Amils, R. Visualizing microorganism-mineral interaction in the Iberian Pyrite Belt subsurface: The Acidovorax case. Front. Microbiol. 2020, 11, 572104. [Google Scholar] [CrossRef] [PubMed]
- Amulya, K.; Mohan, S.V. Fixation of CO2, electron donor and redox microenvironment regulate succinic acid production in Citrobacter amalonaticus. Sci. Total Environ. 2019, 695, 133838. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Wang, Y.; Yao, S.; Hu, C.; Ma, T.; Xia, W. Bioconversion of carbon dioxide to succinate by Citrobacter. Chem. Eng. J. 2023, 452, 139668. [Google Scholar] [CrossRef]
- Ajmal, A.W.; Saroosh, S.; Mulk, S.; Hassan, M.N.; Yasmin, H.; Jabeen, Z.; Nosheen, A.; Shah, S.M.U.; Naz, R.; Hasnain, Z. Bacteria isolated from wastewater irrigated agricultural soils adapt to heavy metal toxicity while maintaining their plant growth promoting traits. Sustainability 2021, 13, 7792. [Google Scholar] [CrossRef]
- Hassan, S.; Abskharon, R.; Gad El-Rab, S.; Shoreit, A. Isolation, characterization of heavy metal resistant strain of Pseudomonas aeruginosa isolated from polluted sites in Assiut city, Egypt. J. Basic Microbiol. 2008, 48, 168–176. [Google Scholar] [CrossRef]
- Hassen, A.; Saidi, N.; Cherif, M.; Boudabous, A. Resistance of environmental bacteria to heavy metals. Bioresour. Technol. 1998, 64, 7–15. [Google Scholar] [CrossRef]
- Keramati, P.; Hoodaji, M.; Tahmourespour, A. Multi-metal resistance study of bacteria highly resistant to mercury isolated from dental clinic effluent. Afr. J. Microbiol. Res. 2011, 5, 831–837. [Google Scholar]
- Hussain, S.; Khan, M.; Sheikh, T.M.M.; Mumtaz, M.Z.; Chohan, T.A.; Shamim, S.; Liu, Y. Zinc essentiality, toxicity, and its bacterial bioremediation: A comprehensive insight. Front. Microbiol. 2022, 13, 900740. [Google Scholar] [CrossRef]
- Boyd, S.M.; Rhinehardt, K.L.; Ewunkem, A.J.; Harrison, S.H.; Thomas, M.D.; Graves, J.L., Jr. Experimental Evolution of Copper Resistance in Escherichia coli Produces Evolutionary Trade-Offs in the Antibiotics Chloramphenicol, Bacitracin, and Sulfonamide. Antibiotics 2022, 11, 711. [Google Scholar] [CrossRef] [PubMed]
- Diep, P.; Leo Shen, H.; Wiesner, J.A.; Mykytczuk, N.; Papangelakis, V.; Yakunin, A.F.; Mahadevan, R. Engineered nickel bioaccumulation in Escherichia coli by NikABCDE transporter and metallothionein overexpression. Eng. Life Sci. 2023, 23, 2200133. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, R.; Srivastava, S. Zinc resistance mechanisms in bacteria. Curr. Sci. 2001, 81, 768. Available online: https://www.jstor.org/stable/24106396 (accessed on 21 July 2023).
- Roberts, C.S.; Ni, F.; Mitra, B. The zinc and iron binuclear transport center of ZupT, a ZIP transporter from Escherichia coli. Biochemistry 2021, 60, 3738–3752. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbon Source | Growth | |
|---|---|---|
| Aerobic Conditions | Anaerobic Conditions | |
| Acetate | + | + |
| Arabinose | + | ND |
| Citrate | + | − |
| Fructose | + | ND |
| Fumarate | ND | + |
| Galactose | + | + (G) |
| Glucose | + | + (G) |
| Glycerol | + | + (G) |
| Lactate | + | ND |
| Lactose | ND | + |
| Malic acid | ND | + |
| Mannitol | + | ND |
| Pyruvate | + | + |
| Starch | + | ND |
| Sucrose | + | + (G) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallego-Rodríguez, V.; Martínez-Bonilla, A.; Rodríguez, N.; Amils, R. Dark Fermentation in the Dark Biosphere: The Case of Citrobacter sp. T1.2D-12. Fermentation 2023, 9, 887. https://doi.org/10.3390/fermentation9100887
Gallego-Rodríguez V, Martínez-Bonilla A, Rodríguez N, Amils R. Dark Fermentation in the Dark Biosphere: The Case of Citrobacter sp. T1.2D-12. Fermentation. 2023; 9(10):887. https://doi.org/10.3390/fermentation9100887
Chicago/Turabian StyleGallego-Rodríguez, Violeta, Adrián Martínez-Bonilla, Nuria Rodríguez, and Ricardo Amils. 2023. "Dark Fermentation in the Dark Biosphere: The Case of Citrobacter sp. T1.2D-12" Fermentation 9, no. 10: 887. https://doi.org/10.3390/fermentation9100887
APA StyleGallego-Rodríguez, V., Martínez-Bonilla, A., Rodríguez, N., & Amils, R. (2023). Dark Fermentation in the Dark Biosphere: The Case of Citrobacter sp. T1.2D-12. Fermentation, 9(10), 887. https://doi.org/10.3390/fermentation9100887