
High-Titer Bioethanol Production from Steam-Exploded Corn Stover Using an Engineering Saccharomyces cerevisiae Strain with High Inhibitor Tolerance

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Raw Materials
2.2. Enzymatic Hydrolysis of the Steam Exploded Pulp
2.3. Strains Construction
2.4. Adaptive Laboratory Evolution of the Engineered Strains
2.5. Batch Ethanol Fermentation
2.6. Assay
2.7. Statistical Analysis
3. Results and Discussions
3.1. Composition of the SECSH and the Fermentation Performance of the Background Strain M3013
3.2. Construction of Xylose-Fermenting S. cerevisiae Strains and Their Adaptive Evolution
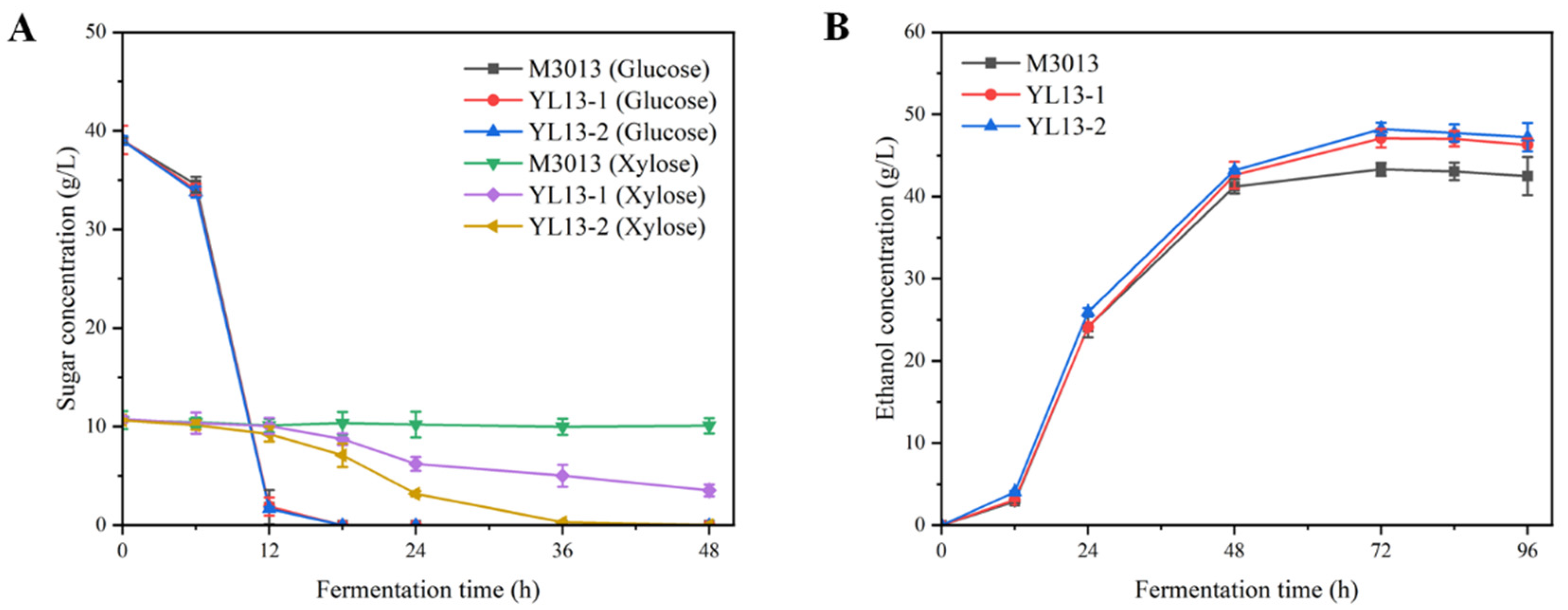
3.3. Fermentation Performance of the Evolved Strain YL13-2 Using SECSH
3.4. Bioethanol Fermentation Using High-Concentration SECSH and Strain YL13-2
3.5. Mass Balance
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ayodele, B.V.; Alsaffar, M.A.; Mustapa, S.I. An overview of integration opportunities for sustainable bioethanol production from first- and second-generation sugar-based feedstocks. J. Clean. Prod. 2020, 245, 118857. [Google Scholar] [CrossRef]
- Kumari, D.; Singh, R. Pretreatment of lignocellulosic wastes for biofuel production: A critical review. Renew. Sustain. Energy Rev. 2018, 90, 877–891. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, H.; Pang, S.; Su, C.; Lv, M.; An, N.; Wang, K.; Cai, D.; Qin, P. Importance of redefinition of corn stover harvest time to enhancing non-food bio-ethanol production. Renew. Energy 2020, 146, 1444–1450. [Google Scholar] [CrossRef]
- Sharma, B.; Larroche, C.; Dussap, C.-G. Comprehensive assessment of 2G bioethanol production. Bioresour. Technol. 2020, 313, 123630. [Google Scholar] [CrossRef] [PubMed]
- Devi, A.; Singh, A.; Bajar, S.; Pant, D.; Din, Z.U. Ethanol from lignocellulosic biomass: An in-depth analysis of pre-treatment methods, fermentation approaches and detoxification processes. J. Environ. Chem. Eng. 2021, 9, 105798. [Google Scholar] [CrossRef]
- Rosales-Calderon, O.; Arantes, V. A review on commercial-scale high-value products that can be produced alongside cellulosic ethanol. Biotechnol. Biofuels 2019, 12, 1–58. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Sankaran, R.; Show, P.L.; Ibrahim, T.N.B.T.; Chew, K.W.; Culaba, A.; Chang, J.-S. Pretreatment methods for lignocellulosic biofuels production: Current advances, challenges and future prospects. Biofuel Res. J. 2020, 7, 1115–1127. [Google Scholar] [CrossRef]
- Chen, H.; Huo, W.; Wang, B.; Wang, Y.; Wen, H.; Cai, D.; Zhang, C.; Wu, Y.; Qin, P. L-lactic acid production by simultaneous saccharification and fermentation of dilute ethylediamine pre-treated rice straw. Ind. Crop. Prod. 2019, 141, 111749. [Google Scholar] [CrossRef]
- Li, P.; Cai, D.; Zhang, C.; Li, S.; Qin, P.; Chen, C.; Wang, Y.; Wang, Z. Comparison of two-stage acid-alkali and alkali-acid pretreatments on enzymatic saccharification ability of the sweet sorghum fiber and their physicochemical characterizations. Bioresour. Technol. 2016, 221, 636–644. [Google Scholar] [CrossRef]
- Zhang, C.; Wen, H.; Zheng, J.; Fu, C.; Chen, C.; Cai, D.; Qin, P.; Wang, Z. A combination of evaporation and chemical preservation for long-term storage of fresh sweet sorghum juice and subsequent bioethanol production. J. Food Process. Preserv. 2018, 42, e13825. [Google Scholar] [CrossRef]
- Li, P.; Cai, D.; Luo, Z.; Qin, P.; Chen, C.; Wang, Y.; Zhang, C.; Wang, Z.; Tan, T. Effect of acid pretreatment on different parts of corn stalk for second generation ethanol production. Bioresour. Technol. 2016, 206, 86–92. [Google Scholar] [CrossRef]
- Yu, Y.; Wu, J.; Ren, X.; Lau, A.; Rezaei, H.; Takada, M.; Bi, X.; Sokhansanj, S. Steam explosion of lignocellulosic biomass for multiple advanced bioenergy processes: A review. Renew. Sustain. Energy Rev. 2022, 154, 111871. [Google Scholar] [CrossRef]
- Singh, J.; Suhag, M.; Dhaka, A. Augmented digestion of lignocellulose by steam explosion, acid and alkaline pretreatment methods: A review. Carbohydr. Polym. 2015, 117, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Jacquet, N.; Maniet, G.; Vanderghem, C.; Delvigne, F.; Richel, A. Application of Steam Explosion as Pretreatment on Lignocellulosic Material: A Review. Ind. Eng. Chem. Res. 2015, 54, 2593–2598. [Google Scholar] [CrossRef]
- Brandt, B.A.; Jansen, T.; Görgens, J.F.; van Zyl, W.H. Overcoming lignocellulose-derived microbial inhibitors: Advancing the Saccharomyces cerevisiae resistance toolbox. Biofuels Bioprod. Biorefining 2019, 13, 1520–1536. [Google Scholar] [CrossRef]
- Qin, L.; Dong, S.; Yu, J.; Ning, X.; Xu, K.; Zhang, S.-J.; Xu, L.; Li, B.-Z.; Li, J.; Yuan, Y.-J.; et al. Stress-driven dynamic regulation of multiple tolerance genes improves robustness and productive capacity of Saccharomyces cerevisiae in industrial lignocellulose fermentation. Metab. Eng. 2020, 61, 160–170. [Google Scholar] [CrossRef]
- Sasano, Y.; Watanabe, D.; Ukibe, K.; Inai, T.; Ohtsu, I.; Shimoi, H.; Takagi, H. Overexpression of the yeast transcription activator Msn2 confers furfural resistance and increases the initial fermentation rate in ethanol production. J. Biosci. Bioeng. 2012, 113, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; Cheong, D.-Y.; Aita, G.M. Optimization of activated carbon detoxification of dilute ammonia pretreated energy cane bagasse enzymatic hydrolysate by response surface methodology. Ind. Crop. Prod. 2018, 115, 166–173. [Google Scholar] [CrossRef]
- Dong, J.-J.; Han, R.-Z.; Xu, G.-C.; Gong, L.; Xing, W.-R.; Ni, Y. Detoxification of furfural residues hydrolysate for butanol fermentation by Clostridium saccharobutylicum DSM 13864. Bioresour. Technol. 2018, 259, 40–45. [Google Scholar] [CrossRef]
- Kim, D. Physico-Chemical Conversion of Lignocellulose: Inhibitor Effects and Detoxification Strategies: A Mini Review. Molecules 2018, 23, 309. [Google Scholar] [CrossRef]
- Li, B.; Liu, N.; Zhao, X. Response mechanisms of Saccharomyces cerevisiae to the stress factors present in lignocellulose hydrolysate and strategies for constructing robust strains. Biotechnol. Biofuels Bioprod. 2022, 15, 28. [Google Scholar] [CrossRef]
- Wang, L.; Li, B.; Su, R.-R.; Wang, S.-P.; Xia, Z.-Y.; Xie, C.-Y.; Tang, Y.-Q. Screening novel genes by a comprehensive strategy to construct multiple stress-tolerant industrial Saccharomyces cerevisiae with prominent bioethanol production. Biotechnol. Biofuels Bioprod. 2022, 15, 11. [Google Scholar] [CrossRef]
- Hemansi; Himanshu; Patel, A.K.; Saini, J.K.; Singhania, R.R. 23. Hemansi; Himanshu; Patel, A.K.; Saini, J.K.; Singhania, R.R. Development of multiple inhibitor tolerant yeast via adaptive laboratory evolution for sustainable bioethanol production. Bioresour. Technol. 2022, 344 Pt B, 126247. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shen, Y.; Wu, M.; Hou, J.; Jiao, C.; Li, Z.; Liu, X.; Bao, X. Engineering a wild-type diploid Saccharomyces cerevisiae strain for second-generation bioethanol production. Bioresour. Bioprocess. 2016, 3, 51. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Li, G.; Liu, Q.; Pereira, R.; Chen, Y.; Nielsen, J. Metabolic network remodelling enhances yeast’s fitness on xylose using aerobic glycolysis. Nat. Catal. 2021, 4, 783–796. [Google Scholar] [CrossRef]
- Ochoa-Chacón, A.; Martinez, A.; Poggi-Varaldo, H.M.; Villa-Tanaca, L.; Ramos-Valdivia, A.C.; Ponce-Noyola, T. Xylose Metabolism in Bioethanol Production: Saccharomyces cerevisiae vs Non-Saccharomyces Yeasts. BioEnergy Res. 2021, 15, 905–923. [Google Scholar] [CrossRef]
- Park, H.; Jeong, D.; Shin, M.; Kwak, S.; Oh, E.J.; Ko, J.K.; Kim, S.R. Xylose utilization in Saccharomyces cerevisiae during conversion of hydrothermally pretreated lignocellulosic biomass to ethanol. Appl. Microbiol. Biotechnol. 2020, 104, 3245–3252. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wen, J.; Wang, K.; Su, C.; Chen, C.; Cui, Z.; Cai, D.; Cheng, S.; Cao, H.; Qin, P. Understanding the Dynamics of the Saccharomyces cerevisiae and Scheffersomyces stipitis Abundance in Co-culturing Process for Bioethanol Production from Corn Stover. Waste Biomass Valorization 2022, 14, 43–55. [Google Scholar] [CrossRef]
- Mei, G.-Y.; Bajwa, P.K.; Dashtban, M.; Ho, C.-Y.; Lee, H. Transfer of plasmid into the pentose-fermenting yeast Pachysolen tannophilus. J. Microbiol. Methods 2018, 148, 97–103. [Google Scholar] [CrossRef]
- Li, Y.-J.; Wang, M.-M.; Chen, Y.-W.; Wang, M.; Fan, L.-H.; Tan, T.-W. Engineered yeast with a CO2-fixation pathway to improve the bio-ethanol production from xylose-mixed sugars. Sci. Rep. 2017, 7, 43875. [Google Scholar] [CrossRef]
- Borgström, C.; Wasserstrom, L.; Almqvist, H.; Broberg, K.; Klein, B.; Noack, S.; Lidén, G.; Gorwa-Grauslund, M.F. Identification of modifications procuring growth on xylose in recombinant Saccharomyces cerevisiae strains carrying the Weimberg pathway. Metab. Eng. 2019, 55, 1–11. [Google Scholar] [CrossRef]
- Demeke, M.M.; Dumortier, F.; Li, Y.; Broeckx, T.; Foulquié-Moreno, M.R.; Thevelein, J.M. Combining inhibitor tolerance and D-xylose fermentation in industrial Saccharomyces cerevisiae for efficient lignocellulose-based bioethanol production. Biotechnol. Biofuels 2013, 6, 120. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Hao, B.; Yu, H.; Tu, F.; Wei, X.; Xiong, K.; Zeng, Y.; Zeng, H.; Liu, P.; Tu, Y.; et al. Double integrating XYL2 into engineered Saccharomyces cerevisiae strains for consistently enhanced bioethanol production by effective xylose and hexose co-consumption of steam-exploded lignocellulose in bioenergy crops. Renew. Energy 2022, 186, 341e349. [Google Scholar] [CrossRef]
- Chen, S.; Xu, Z.; Ding, B.; Zhang, Y.; Liu, S.; Cai, C.; Li, M.; Dale, B.E.; Jin, M. Big data mining, rational modification, and ancestral sequence reconstruction inferred multiple xylose isomerases for biorefinery. Sci. Adv. 2023, 9, eadd8835. [Google Scholar] [CrossRef]
- Tang, R.; Ye, P.; Alper, H.S.; Liu, Z.; Zhao, X.; Bai, F. Identification and characterization of novel xylose isomerases from a Bos taurus fecal metagenome. Appl. Microbiol. Biotechnol. 2019, 103, 9465–9477. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.E.; Romaní, A.; Cunha, J.T.; Johansson, B.; Domingues, L. Integrated approach for selecting efficient Saccharomyces cerevisiae for industrial lignocellulosic fermentations: Importance of yeast chassis linked to process conditions. Bioresour. Technol. 2017, 227, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Zhang, C.; Wu, Y.; Zhu, Q.; Wen, J.; Wang, Y.; Zhao, J.; Liu, Y.; Qin, P.; Cai, D. Combination of pH adjusting and intermittent feeding can improve fermentative acetone-butanol-ethanol (ABE) production from steam exploded corn stover. Renew. Energy 2022, 200, 592–600. [Google Scholar] [CrossRef]
- Cai, D.; Dong, Z.; Wang, Y.; Chen, C.; Li, P.; Qin, P.; Wang, Z.; Tan, T. Biorefinery of corn cob for microbial lipid and bio-ethanol production: An environmental friendly process. Bioresour. Technol. 2016, 211, 677–684. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Wang, Z.; Zhang, Y.; Shi, S.; Nielsen, J.; Liu, Z. A gRNA-tRNA array for CRISPR-Cas9 based rapid multiplexed genome editing in Saccharomyces cerevisiae. Nat. Commun. 2019, 10, 1053. [Google Scholar] [CrossRef]
- Xia, F.; Du, J.; Wang, K.; Liu, L.; Ba, L.; Liu, H.; Liu, Y. Application of Multiple Strategies To Debottleneck the Biosynthesis of Longifolene by Engineered Saccharomyces cerevisiae. J. Agric. Food Chem. 2022, 70, 11336–11343. [Google Scholar] [CrossRef]
- Wu, Y.; Wen, J.; Su, C.; Jiang, C.; Zhang, C.; Wang, Y.; Jiang, Y.; Ren, W.; Qin, P.; Cai, D. Inhibitions of microbial fermentation by residual reductive lignin oil: Concerns on the bioconversion of reductive catalytic fractionated carbohydrate pulp. Chem. Eng. J. 2023, 452, 139267. [Google Scholar] [CrossRef]
- Lyu, G.; Yoo, C.G.; Pan, X. Alkaline oxidative cracking for effective depolymerization of biorefining lignin to mono-aromatic compounds and organic acids with molecular oxygen. Biomass-Bioenergy 2018, 108, 7–14. [Google Scholar] [CrossRef]
- Vanmarcke, G.; Demeke, M.M.; Foulquié-Moreno, M.R.; Thevelein, J.M. Identification of the major fermentation inhibitors of recombinant 2G yeasts in diverse lignocellulose hydrolysates. Biotechnol. Biofuels 2021, 14, 92. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.; Palmqvist, E.; Hahn-Hägerdal, B.; Tengborg, C.; Stenberg, K.; Zacchi, G.; Nilvebrant, N.-O. The generation of fermentation inhibitors during dilute acid hydrolysis of softwood. Enzym. Microb. Technol. 1999, 24, 151–159. [Google Scholar] [CrossRef]
- Keshav, P.K.; Banoth, C.; Kethavath, S.N.; Bhukya, B. Lignocellulosic ethanol production from cotton stalk: An overview on pretreatment, saccharification and fermentation methods for improved bioconversion process. Biomass Convers. Biorefinery 2021, 13, 4477–4493. [Google Scholar] [CrossRef]
- Jhariya, U.; Dafale, N.A.; Srivastava, S.; Bhende, R.S.; Kapley, A.; Purohit, H.J. Understanding Ethanol Tolerance Mechanism in Saccharomyces cerevisiae to Enhance the Bioethanol Production: Current and Future Prospects. BioEnergy Res. 2021, 14, 670–688. [Google Scholar] [CrossRef]
- Liu, C.-G.; Li, K.; Li, K.-Y.; Sakdaronnarong, C.; Mehmood, M.A.; Zhao, X.-Q.; Bai, F.-W. Intracellular Redox Perturbation in Saccharomyces cerevisiae Improved Furfural Tolerance and Enhanced Cellulosic Bioethanol Production. Front. Bioeng. Biotechnol. 2020, 8, 615. [Google Scholar] [CrossRef]
- Menardo, S.; Balsari, P.; Tabacco, E.; Borreani, G. Effect of Conservation Time and the Addition of Lactic Acid Bacteria on the Biogas and Methane Production of Corn Stalk Silage. BioEnergy Res. 2015, 8, 1810–1823. [Google Scholar] [CrossRef]
- Martín, C.; Galbe, M.; Wahlbom, C.; Hahn-Hägerdal, B.; Jönsson, L.J. Ethanol production from enzymatic hydrolysates of sugarcane bagasse using recombinant xylose-utilising Saccharomyces cerevisiae. Enzym. Microb. Technol. 2002, 31, 274–282. [Google Scholar] [CrossRef]
- Bensid, A.; El Abed, N.; Houicher, A.; Regenstein, J.M.; Özogul, F. Antioxidant and antimicrobial preservatives: Properties, mechanism of action and applications in food—A review. Crit. Rev. Food Sci. Nutr. 2020, 62, 2985–3001. [Google Scholar] [CrossRef]
- Pei, W.; Deng, J.; Wang, P.; Wang, X.; Zheng, L.; Zhang, Y.; Huang, C. Sustainable lignin and lignin-derived compounds as potential therapeutic agents for degenerative orthopaedic diseases: A systemic review. Int. J. Biol. Macromol. 2022, 212, 547–560. [Google Scholar] [CrossRef]
- Karhumaa, K.; Sanchez, R.G.; Hahn-Hägerdal, B.; Gorwa-Grauslund, M.-F. Comparison of the xylose reductase-xylitol dehydrogenase and the xylose isomerase pathways for xylose fermentation by recombinant Saccharomyces cerevisiae. Microb. Cell Factories 2007, 6, 5. [Google Scholar] [CrossRef]
- Watanabe, S.; Abu Saleh, A.; Pack, S.P.; Annaluru, N.; Kodaki, T.; Makino, K. Ethanol production from xylose by recombinant Saccharomyces cerevisiae expressing protein-engineered NADH-preferring xylose reductase from Pichia stipitis. Microbiology 2007, 153 Pt 9, 3044–3054. [Google Scholar] [CrossRef] [PubMed]
- Cunha, J.T.; Romaní, A.; Costa, C.E.; Sá-Correia, I.; Domingues, L. Molecular and physiological basis of Saccharomyces cerevisiae tolerance to adverse lignocellulose-based process conditions. Appl. Microbiol. Biotechnol. 2018, 103, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Kumakiri, I.; Yokota, M.; Tanaka, R.; Shimada, Y.; Kiatkittipong, W.; Lim, J.W.; Murata, M.; Yamada, M. Process Intensification in Bio-Ethanol Production–Recent Developments in Membrane Separation. Processes 2021, 9, 1028. [Google Scholar] [CrossRef]
- Vane, L.M. Separation technologies for the recovery and dehydration of alcohols from fermentation broths. Biofuels, Bioprod. Biorefining 2008, 2, 553–588. [Google Scholar] [CrossRef]
- Stanley, D.; Bandara, A.; Fraser, S.; Chambers, P.; Stanley, G. The ethanol stress response and ethanol tolerance of Saccharomyces cerevisiae. J. Appl. Microbiol. 2010, 109, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Huang, X.; Zhang, L.; Zhao, N.; Yang, D.; Zhang, K. Tolerance and stress response to ethanol in the yeast Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2009, 85, 253–263. [Google Scholar] [CrossRef]
- Sun, C.; Meng, X.; Sun, F.; Zhang, J.; Tu, M.; Chang, J.-S.; Reungsang, A.; Xia, A.; Ragauskas, A.J. Advances and perspectives on mass transfer and enzymatic hydrolysis in the enzyme-mediated lignocellulosic biorefinery: A review. Biotechnol. Adv. 2023, 62, 108059. [Google Scholar] [CrossRef]
- da Silva, A.S.; Espinheira, R.P.; Teixeira, R.S.S.; de Souza, M.F.; Ferreira-Leitão, V.; Bon, E.P.S. Constraints and advances in high-solids enzymatic hydrolysis of lignocellulosic biomass: A critical review. Biotechnol. Biofuels 2020, 13, 58. [Google Scholar] [CrossRef]
- Su, C.; Qi, L.; Cai, D.; Chen, B.; Chen, H.; Zhang, C.; Si, Z.; Wang, Z.; Li, G.; Qin, P. Integrated ethanol fermentation and acetone-butanol-ethanol fermentation using sweet sorghum bagasse. Renew. Energy 2020, 162, 1125–1131. [Google Scholar] [CrossRef]
- Modenbach, A.A.; Nokes, S.E. Enzymatic hydrolysis of biomass at high-solids loadings—A review. Biomass- Bioenergy 2013, 56, 526–544. [Google Scholar] [CrossRef]
- Li, M.; Wang, L.; Zhao, Q.; Chen, H. High Concentration of Fermentable Sugars Prepared from Steam Exploded Lignocellulose in Periodic Peristalsis Integrated Fed-Batch Enzymatic Hydrolysis. Appl. Biochem. Biotechnol. 2022, 194, 5255–5273. [Google Scholar] [CrossRef]
- Afedzi, A.E.K.; Rattanaporn, K.; Parakulsuksatid, P. Impeller selection for mixing high-solids lignocellulosic biomass in stirred tank bioreactor for ethanol production. Bioresour. Technol. Rep. 2022, 17, 100935. [Google Scholar] [CrossRef]
- Diao, L.; Liu, Y.; Qian, F.; Yang, J.; Jiang, Y.; Yang, S. Construction of fast xylose-fermenting yeast based on industrial ethanol-producing diploid Saccharomyces cerevisiaeby rational design and adaptive evolution. BMC Biotechnol. 2013, 13, 110. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.K.; Enkh-Amgalan, T.; Gong, G.; Um, Y.; Lee, S. Improved bioconversion of lignocellulosic biomass by Saccharomyces cerevisiae engineered for tolerance to acetic acid. GCB Bioenergy 2019, 12, 90–100. [Google Scholar] [CrossRef]
- Tran, P.H.N.; Ko, J.K.; Gong, G.; Um, Y.; Lee, S.-M. Improved simultaneous co-fermentation of glucose and xylose by Saccharomyces cerevisiae for efficient lignocellulosic biorefinery. Biotechnol. Biofuels 2020, 13, 12. [Google Scholar] [CrossRef]
- Tomás-Pejó, E.; Oliva, J.M.; Ballesteros, M.; Olsson, L. Comparison of SHF and SSF processes from steam-exploded wheat straw for ethanol production by xylose-fermenting and robust glucose-fermentingSaccharomyces cerevisiae strains. Biotechnol. Bioeng. 2008, 100, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Cunha, J.T.; Soares, P.O.; Romaní, A.; Thevelein, J.M.; Domingues, L. Xylose fermentation efficiency of industrial Saccharomyces cerevisiae yeast with separate or combined xylose reductase/xylitol dehydrogenase and xylose isomerase pathways. Biotechnol. Biofuels 2019, 12, 20. [Google Scholar] [CrossRef]
- Wei, F.; Li, M.; Wang, M.; Li, H.; Li, Z.; Qin, W.; Wei, T.; Zhao, J.; Bao, X. A C6/C5 co-fermenting Saccharomyces cerevisiae strain with the alleviation of antagonism between xylose utilization and robustness. GCB Bioenergy 2020, 13, 83–97. [Google Scholar] [CrossRef]
- Yuan, Z.; Li, G.; Hegg, E.L. Enhancement of sugar recovery and ethanol production from wheat straw through alkaline pre-extraction followed by steam pretreatment. Bioresour. Technol. 2018, 266, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Tsuge, Y.; Sasaki, D.; Teramura, H.; Inokuma, K.; Hasunuma, T.; Ogino, C.; Kondo, A. Mechanical milling and membrane separation for increased ethanol production during simultaneous saccharification and co-fermentation of rice straw by xylose-fermenting Saccharomyces cerevisiae. Bioresour. Technol. 2015, 185, 263–268. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibitors | Formate | Acetate | Lactate | Furfural | 5-HMF | Phenol | 4-Methylphenol | Benzoic Acid | 4-Hydroxy-Benzaldehyde |
|---|---|---|---|---|---|---|---|---|---|
| Concentration (g/L) | 2.110 ± 0.198 | 6.153 ± 0.803 | 3.355 ± 0.136 | 0.333 ± 0.001 | 0.412 ± 0.004 | 0.021 ± 0.001 | 0.010 ± 0.000 | 0.123 ± 0.008 | 0.022 ± 0.004 |
| Inhibitors | p-Hydroxyacetophenone | 3-Hydroxy-3-(4-hydroxyphenyl)propanoic acid | 4-Hydroxyphenethyl alcohol | 4-Hydroxy benzonic acid | 4-Hydroxy-benzenepropanoic acid | Methylhydroquinone | Protocatechuate | 3-(4-Hydroxyphenyl)lactate | p-Coumaric acid |
| Concentration (g/L) | 0.014 ± 0.002 | 0.002 ± 0.001 | 0.003 ± 0.001 | 0.015 ± 0.005 | 0.002 ± 0.001 | 0.002 ± 0.000 | 0.011 ± 0.004 | 0.032 ± 0.014 | 0.035 ± 0.006 |
| Inhibitors | 2-Methoxyphenol | Vanillin | 4-Hydroxy-3-methoxyphenethylene glycol | Acetovanillone | Vanillic acid | 4-Hydroxy-3-methoxybenzeneisopropanoic acid | 4-Hydroxy-3-methoxyphenylacetic acid | Vanillylmandelic acid | 2-hydroxy-3-(4-hydroxy-3-methoxyphenyl) propanoic acid |
| Concentration (g/L) | 0.012 ± 0.004 | 0.064 ± 0.012 | 0.012 ± 0.005 | 0.010 ± 0.003 | 0.044 ± 0.001 | 0.007 ± 0.001 | 0.023 ± 0.007 | 0.008 ± 0.002 | 0.001 ± 0.000 |
| Inhibitors | Ferulic acid | 2,6-dimethoxyphenol | Syringaldehyde | Syringic acid | |||||
| Concentration (g/L) | 0.034 ± 0.002 | 0.015 ± 0.006 | 0.010 ± 0.002 | 0.020 ± 0.007 |
| Strains | Substrate | Pretreatment Conditions | Description | Detoxification | Nutrient Supplements | Ethanol Concentration (g/L) | Ethanol Yield (g/g) | Refs. |
|---|---|---|---|---|---|---|---|---|
| S. cerevisiae YL13-2 | Corn stover | SE | M3013, mXYL1, XYL1, XYL2, XKS1, TAL1, TKL1, RKI1, RPE1, ∆GRE3, ∆PHO13, ALE | No | Without additional nutrients | 51.12 | 0.436 | This study |
| S. cerevisiae XUSAE57 | Sugarcane bagasse | Dilute acid pretreatment | BY4741, xylA*3, TAL1, XKS1, RPE1, ΔGRE3, ΔPHO13, ALE | Yes | Without additional nutrients | 23.2 | 0.49 | [66] |
| S. cerevisiae XUSEA | Micanthus sacchariflorus Goedae-Uksae | H2SO4 pretreatment | BY4741, xylA*3, TAL1, XKS1, RPE1, ΔGRE3, ΔPHO13, ΔACS1, ALE | Yes | Without additional nutrients | 30.1 | 0.48 | [67] |
| S. cerevisiae F12 | Wheat straw | SE | TMB3001, XYL1, XYL2, XKS1, ALE | No | vitamin solution and ergosterol | 23.7 | 0.43 | [68] |
| S. cerevisiae PE-2ΔGRE3-X | Corn cob | Hydrothermal treatment | PE-2, xylA, ∆GRE3 | Yes | Yeast extract 10 g/L, peptone 20 g/L | 11.6 | 0.44 | [69] |
| S. cerevisiae LF1 | Corn stover | SE | BSIF (diploid), Ru-xylA, TAL1, MGT05196N360F, TKL1, RKI1, RPE1, ∆GRE3, ∆PHO13, ALE | No | Yeast extract 10 g/L, peptone 20 g/L | 50.81 | 0.413 | [24] |
| S. cerevisiae XM20 | Corn stover | Ethylenediamine (EDA) pretreatment | L2612, XYL1, mXYL2, XKS1, TKL1, RKI1, SOD1, GSH1, GLR1, ZWF1, GND2, ACS1, ΔPHO13 | No | Without additional nutrients | 90.7 | / | [16] |
| S. cerevisiae CRD5HS | Corn stover | Alkaline/acidic pretreatment | CRD3, hasxylA, ALE | No | Yeast extract 5 g/L and tryptone 10 g/L | 85.95 | / | [34] |
| S. cerevisiae 6M-15 | Corn straw | SE | LF1, ARTP mutagenesis | No | Yeast extract 10 g/L, peptone 20 g/L | 30.95 | 0.43 | [70] |
| S. cerevisiae Lg8-1 | Wheat straw | Alkaline pre-extraction and acid catalyzed steam treatment | / | No | Without additional nutrients | 54.5 | 0.46 | [71] |
| S. cerevisiae MN8140X/TF-TF | Rice straw | Hydrothermally pretreated and mechanically milled | MT8-10, XYL1, XYL2, XKS1, TAL1, FDH1 | No | Yeast extract 10 g/L, peptone 20 g/L | 52.0 | 0.38 | [72] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Su, C.; Zhang, G.; Liao, Z.; Wen, J.; Wang, Y.; Jiang, Y.; Zhang, C.; Cai, D. High-Titer Bioethanol Production from Steam-Exploded Corn Stover Using an Engineering Saccharomyces cerevisiae Strain with High Inhibitor Tolerance. Fermentation 2023, 9, 906. https://doi.org/10.3390/fermentation9100906
Wu Y, Su C, Zhang G, Liao Z, Wen J, Wang Y, Jiang Y, Zhang C, Cai D. High-Titer Bioethanol Production from Steam-Exploded Corn Stover Using an Engineering Saccharomyces cerevisiae Strain with High Inhibitor Tolerance. Fermentation. 2023; 9(10):906. https://doi.org/10.3390/fermentation9100906
Chicago/Turabian StyleWu, Yilu, Changsheng Su, Gege Zhang, Zicheng Liao, Jieyi Wen, Yankun Wang, Yongjie Jiang, Changwei Zhang, and Di Cai. 2023. "High-Titer Bioethanol Production from Steam-Exploded Corn Stover Using an Engineering Saccharomyces cerevisiae Strain with High Inhibitor Tolerance" Fermentation 9, no. 10: 906. https://doi.org/10.3390/fermentation9100906
APA StyleWu, Y., Su, C., Zhang, G., Liao, Z., Wen, J., Wang, Y., Jiang, Y., Zhang, C., & Cai, D. (2023). High-Titer Bioethanol Production from Steam-Exploded Corn Stover Using an Engineering Saccharomyces cerevisiae Strain with High Inhibitor Tolerance. Fermentation, 9(10), 906. https://doi.org/10.3390/fermentation9100906