
Identification and Quantification of Lipopeptide Homologues Induced and Produced by Bacillus amyloliquefaciens


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Reagents
2.2. Bacterial Culture
2.3. Biosynthesis and Extraction of Lipopeptides
2.4. Identification and Quantification of Homologues of Lipopeptides Using UPLC-MS/MS
2.5. Calibration Curves
2.6. Statistical Analysis
3. Results and Discussion
3.1. UPLC-ESI-MS Analysis
3.2. ESI-MS/MS Analysis of the Iturin Family
3.3. ESI-MS/MS Analysis of Fengycin A and B
3.4. ESI-MS/MS Analysis of Surfactin
3.5. Quantification of Lipopeptide Homologues
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Biniarz, P.; Coutte, F.; Gancel, F.; Łukaszewicz, M. High-Throughput Optimization of Medium Components and Culture Conditions for the Efficient Production of a Lipopeptide Pseudofactin by Pseudomonas fluorescens BD5. Microb. Cell Fact. 2018, 17, 121. [Google Scholar] [CrossRef] [PubMed]
- Bekiesch, P.; Zehl, M.; Domingo-Contreras, E.; Martín, J.; Pérez-Victoria, I.; Reyes, F.; Kaplan, A.; Rückert, C.; Busche, T.; Kalinowski, J.; et al. Viennamycins: Lipopeptides Produced by a Streptomyces Sp. J. Nat. Prod. 2020, 83, 2381–2389. [Google Scholar] [CrossRef] [PubMed]
- Biniarz, P.; Łukaszewicz, M.; Janek, T. Screening Concepts, Characterization and Structural Analysis of Microbial-Derived Bioactive Lipopeptides: A Review. Crit. Rev. Biotechnol. 2017, 37, 393–410. [Google Scholar] [PubMed]
- Balleza, D.; Mescola, A.; Marín–Medina, N.; Ragazzini, G.; Pieruccini, M.; Facci, P.; Alessandrini, A. Complex Phase Behavior of GUVs Containing Different Sphingomyelins. Biophys. J. 2019, 116, 503–517. [Google Scholar] [CrossRef]
- Wang, Q.H.; Ji, Y.P.; Qu, Y.Y.; Qi, Y.K.; Li, D.W.; Liu, Z.Y.; Wu, X.Q. The Response Strategies of Colletotrichum gloeosporioides s.s. Due to the Stress Caused by Biological Control Agent Bacillus amyloliquefaciens Deciphered by Transcriptome Analyses. Biol. Control. 2020, 150, 104372. [Google Scholar] [CrossRef]
- Li, F.Z.; Zeng, Y.J.; Zong, M.H.; Yang, J.G.; Lou, W.Y. Bioprospecting of a Novel Endophytic Bacillus velezensis FZ06 from Leaves of Camellia assamica: Production of Three Groups of Lipopeptides and the Inhibition against Food Spoilage Microorganisms. J. Biotechnol. 2020, 323, 42–53. [Google Scholar] [CrossRef]
- Ley-López, N.; Basilio Heredia, J.; San Martín-Hernández, C.; Ibarra-Rodríguez, J.R.; Angulo-Escalante, M.Á.; García-Estrada, R.S. Induced Biosynthesis of Fengycin and Surfactin in a Strain of Bacillus amyloliquefaciens with Oomyceticidal Activity on Zoospores of Phytophthora capsici. Rev. Argent. Microbiol. 2022, 54, 181–191. [Google Scholar] [CrossRef]
- Sa, R.B.; An, X.; Sui, J.K.; Wang, X.H.; Ji, C.; Wang, C.Q.; Li, Q.; Hu, Y.R.; Liu, X. Purification and Structural Characterization of Fengycin Homologues Produced by Bacillus subtilis from Poplar Wood Bark. Australas. Plant Pathol. 2018, 47, 259–268. [Google Scholar] [CrossRef]
- De Faria, A.F.; Stéfani, D.; Vaz, B.G.; Silva, Í.S.; Garcia, J.S.; Eberlin, M.N.; Grossman, M.J.; Alves, O.L.; Durrant, L.R. Purification and Structural Characterization of Fengycin Homologues Produced by Bacillus subtilis LSFM-05 Grown on Raw Glycerol. J. Ind. Microbiol. Biotechnol. 2011, 38, 863–871. [Google Scholar] [CrossRef]
- Jajor, P.; Piłakowska-Pietras, D.; Krasowska, A.; Łukaszewicz, M. Surfactin Analogues Produced by Bacillus subtilis Strains Grown on Rapeseed Cake. J. Mol. Struct. 2016, 1126, 141–146. [Google Scholar] [CrossRef]
- Tanaka, K.; Amaki, Y.; Ishihara, A.; Nakajima, H. Supporting Information Synergistic Effects of [Ile 7] Surfactin Homologues with Bacillomycin D in Suppression of Gray Mold Disease by Bacillus amyloliquefaciens Biocontrol Strain SD-32. J. Agric. Food Chem. 2015, 63, 5344–5353. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Wang, H.; Tan, Z.; Xuan, Z.; Dahar, G.Y.; Li, Q.X.; Miao, W.; Liu, W. Antifungal Mechanism of Bacillomycin D from Bacillus velezensis HN-2 against Colletotrichum gloeosporioides Penz. Pestic. Biochem. Physiol. 2020, 163, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Sun, C. Fengycins, Cyclic Lipopeptides from Marine Bacillus subtilis Strains, Kill the Plant-Pathogenic Fungus Magnaporthe Grisea by Inducing Reactive Oxygen Species Production and Chromatin Condensation. Appl. Environ. Microbiol. 2018, 84, e00445-18. [Google Scholar] [CrossRef] [PubMed]
- Jemil, N.; Manresa, A.; Rabanal, F.; Ben Ayed, H.; Hmidet, N.; Nasri, M. Structural Characterization and Identification of Cyclic Lipopeptides Produced by Bacillus methylotrophicus DCS1 Strain. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 2017, 1060, 374–386. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Guo, Q.; Ma, Y.; Li, S.; Lu, X.; Zhang, X.; Ma, P. DegQ Regulates the Production of Fengycins and Biofilm Formation of the Biocontrol Agent Bacillus subtilis NCD-2. Microbiol. Res. 2015, 178, 42–50. [Google Scholar] [CrossRef]
- Qian, S.; Sun, J.; Lu, H.; Lu, F.; Bie, X.; Lu, Z. L-Glutamine Efficiently Stimulates Biosynthesis of Bacillomycin D in Bacillus subtilis FmbJ. Process Biochem. 2017, 58, 224–229. [Google Scholar] [CrossRef]
- Emanuel, R.V.; César Arturo, P.U.; Lourdes Iveth, M.R.; Homero, R.d.l.C.; Mauricio Nahuam, C.A. In Vitro Growth of Colletotrichum gloeosporioides Is Affected by Butyl Acetate, a Compound Produced during the Co-Culture of Trichoderma Sp. and Bacillus subtilis. 3 Biotech 2020, 10, 329. [Google Scholar] [CrossRef]
- Ghazala, I.; Bouassida, M.; Krichen, F.; Manuel Benito, J.; Ellouz-Chaabouni, S.; Haddar, A. Anionic Lipopeptides from Bacillus mojavensis I4 as Effective Antihypertensive Agents: Production, Characterization, and Identification. Eng. Life Sci. 2017, 17, 1244–1253. [Google Scholar] [CrossRef]
- Ley-López, N.; Márquez-Zequera, I.; Carrillo-Fasio, J.A.; León-Félix, J.; Cruz-Lachica, I.; García-Estrada, R.S.; Allende-Molar, R. Efecto de Biocontrol e Inhibición Germinativa de Bacillus Spp. Sobre Zoosporas de Phytophthora capsici. Rev. Mex. Fitopatol. Mex. J. Phytopathol. 2018, 36, 215–232. [Google Scholar] [CrossRef]
- Landy, M.; Warren, G.H.; Rosenmanm, S.B.; Colio, L.G. Bacillomycin: An Antibiotic from Bacillus subtilis Active against Pathogenic Fungi. Proc. Soc. Exp. Biol. Med. 1948, 67, 539–541. [Google Scholar] [CrossRef]
- Unás, J.H.; de Alexandria Santos, D.; Azevedo, E.B.; Nitschke, M. Brevibacterium luteolum Biosurfactant: Production and Structural Characterization. Biocatal. Agric. Biotechnol. 2018, 13, 160–167. [Google Scholar] [CrossRef]
- Lee, M.H.; Lee, J.; Nam, Y.D.; Lee, J.S.; Seo, M.J.; Yi, S.H. Characterization of Antimicrobial Lipopeptides Produced by Bacillus Sp. LM7 Isolated from Chungkookjang, a Korean Traditional Fermented Soybean Food. Int. J. Food Microbiol. 2016, 221, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Gong, Q.; Zhang, C.; Lu, F.; Zhao, H.; Bie, X.; Lu, Z. Identification of Bacillomycin D from Bacillus subtilis FmbJ and Its Inhibition Effects against Aspergillus flavus. Food Control 2014, 36, 8–14. [Google Scholar] [CrossRef]
- Luna-Bulbarela, A.; Tinoco-Valencia, R.; Corzo, G.; Kazuma, K.; Konno, K.; Galindo, E.; Serrano-Carreón, L. Effects of Bacillomycin D Homologues Produced by Bacillus amyloliquefaciens 83 on Growth and Viability of Colletotrichum gloeosporioides at Different Physiological Stages. Biol. Control 2018, 127, 145–154. [Google Scholar] [CrossRef]
- Nasir, M.N.; Besson, F. Conformational Analyses of Bacillomycin D, a Natural Antimicrobial Lipopeptide, Alone or in Interaction with Lipid Monolayers at the Air-Water Interface. J. Colloid Interface Sci. 2012, 387, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhou, Y.; Liu, H.; Ni, J.; Lu, F.; Bie, X.; Lu, Z.; Lu, Y. Anti-Toxicogenic Fungi and Toxin-Reducing Effects of Bacillomycin D in Combination with Fungicides. Toxicon 2022, 216, 107–113. [Google Scholar] [CrossRef]
- Bie, X.; Lu, Z.; Lu, F. Identification of Fengycin Homologues from Bacillus subtilis with ESI-MS/CID. J. Microbiol. Methods 2009, 79, 272–278. [Google Scholar] [CrossRef]
- Ramarathnam, R.; Bo, S.; Chen, Y.; Fernando, W.G.D.; Xuewen, G.; De Kievit, T. Molecular and Biochemical Detection of Fengycin- and Bacillomycin D-Producing Bacillus Spp., Antagonistic to Fungal Pathogens of Canola and Wheat. Can. J. Microbiol. 2007, 53, 901–911. [Google Scholar] [CrossRef]
- Hu, L.B.; Shi, Z.Q.; Zhang, T.; Yang, Z.M. Fengycin Antibiotics Isolated from B-FS01 Culture Inhibit the Growth of Fusarium moniliforme Sheldon ATCC 38932. FEMS Microbiol. Lett. 2007, 272, 91–98. [Google Scholar] [CrossRef]
- Wang, J.; Liu, J.; Wang, X.; Yao, J.; Yu, Z. Application of Electrospray Ionization Mass Spectrometry in Rapid Typing of Fengycin Homologues Produced by Bacillus subtilis. Lett. Appl. Microbiol. 2004, 39, 98–102. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, J.; Sun, J.; Zhu, X.; Zhou, L.; Lu, Z.; Lu, Y. C16-Fengycin A Affect the Growth of Candida albicans by Destroying Its Cell Wall and Accumulating Reactive Oxygen Species. Appl. Microbiol. Biotechnol. 2019, 103, 8963–8975. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lu, J.; Sun, J.; Lu, F.; Bie, X.; Lu, Z. Membrane Disruption and DNA Binding of Fusarium graminearum Cell Induced by C16-Fengycin A Produced by Bacillus amyloliquefaciens. Food Control 2019, 102, 206–213. [Google Scholar] [CrossRef]
- Liao, J.H.; Chen, P.Y.; Yang, Y.L.; Kan, S.C.; Hsieh, F.C.; Liu, Y.C. Clarification of the Antagonistic Effect of the Lipopeptides Produced by Bacillus amyloliquefaciens BPD1 against Pyricularia oryzae via in Situ MALDI-TOF IMS Analysis. Molecules 2016, 21, 1670. [Google Scholar] [CrossRef]
- De Faria, A.F.; Teodoro-Martinez, D.S.; De Oliveira Barbosa, G.N.; Gontijo Vaz, B.; Serrano Silva, Í.; Garcia, J.S.; Tótola, M.R.; Eberlin, M.N.; Grossman, M.; Alves, O.L.; et al. Production and Structural Characterization of Surfactin (C 14/Leu7) Produced by Bacillus subtilis Isolate LSFM-05 Grown on Raw Glycerol from the Biodiesel Industry. Process Biochem. 2011, 46, 1951–1957. [Google Scholar] [CrossRef]
- Chen, Z.; Wu, Q.; Wang, L.; Chen, S.; Lin, L.; Wang, H.; Xu, Y. Identification and Quantification of Surfactin, a Nonvolatile Lipopeptide in Moutai Liquor. Int. J. Food Prop. 2020, 23, 189–198. [Google Scholar] [CrossRef]
- Moro, G.V.; Almeida, R.T.R.; Napp, A.P.; Porto, C.; Pilau, E.J.; Lüdtke, D.S.; Moro, A.V.; Vainstein, M.H. Identification and Ultra-High-Performance Liquid Chromatography Coupled with High-Resolution Mass Spectrometry Characterization of Biosurfactants, Including a New Surfactin, Isolated from Oil-Contaminated Environments. Microb. Biotechnol. 2018, 11, 759–769. [Google Scholar] [CrossRef]
- Peypoux, F.; Bonmatin, J.-M.; Labbe, H.; Grangemard, I.; Das, B.C.; Ptak, M.; Wallach, J.; Michel, G. [Ala4]Surfactin, a Novel Isoform from Bacillus subtilis Studied by Mass and NMR Spectroscopies. Eur. J. Biochem. 1994, 224, 89–96. [Google Scholar] [CrossRef]
- Savadogo, A.; Tapi, A.; Chollet, M.; Wathelet, B.; Traoré, A.S.; Jacques, P. Identification of Surfactin Producing Strains in Soumbala and Bikalga Fermented Condiments Using Polymerase Chain Reaction and Matrix Assisted Laser Desorption/Ionization-Mass Spectrometry Methods. Int. J. Food Microbiol. 2011, 151, 299–306. [Google Scholar] [CrossRef]
- Jakab, Á.; Kovács, F.; Balla, N.; Tóth, Z.; Ragyák, Á.; Sajtos, Z.; Csillag, K.; Nagy-Köteles, C.; Nemes, D.; Bácskay, I.; et al. Physiological and Transcriptional Profiling of Surfactin Exerted Antifungal Effect against Candida Albicans. Biomed. Pharmacother. 2022, 152, 113220. [Google Scholar] [CrossRef]
- Fooladi, T.; Moazami, N.; Abdeshahian, P.; Kadier, A.; Ghojavand, H.; Wan Yusoff, W.M.; Hamid, A.A. Characterization, Production and Optimization of Lipopeptide Biosurfactant by New Strain Bacillus pumilus 2IR Isolated from an Iranian Oil Field. J. Pet. Sci. Eng. 2016, 145, 510–519. [Google Scholar] [CrossRef]
- Abouseoud, M.; Maachi, R.; Amrane, A.; Boudergua, S.; Nabi, A. Evaluation of Different Carbon and Nitrogen Sources in Production of Biosurfactant by Pseudomonas fluorescens. Desalination 2008, 223, 143–151. [Google Scholar] [CrossRef]
- Ghribi, D.; Ellouze-Chaabouni, S. Enhancement of Bacillus subtilis Lipopeptide Biosurfactants Production through Optimization of Medium Composition and Adequate Control of Aeration. Biotechnol. Res. Int. 2011, 2011, 653654. [Google Scholar] [CrossRef] [PubMed]
- Yaseen, Y.; Gancel, F.; Drider, D.; Béchet, M.; Jacques, P. Influence of Promoters on the Production of Fengycin in Bacillus Spp. Res. Microbiol. 2016, 167, 272–281. [Google Scholar] [CrossRef]
- Zhang, T.C.; Li, W.; Luo, X.G.; Feng, C.X.; Zhang, M.H.; Du, W.; Ma, D.Y. Increase of the Lycopene Production in the Recombinant Strains of Escherichia coli by Supplementing with Fructose. In Advances in Applied Biotechnology. Proceedings of the 2nd International Conference on Applied Biotechnology (ICAB 2014)-Volume I; Springer: Berlin/Heidelberg, Germany, 2015; Volume 332, pp. 29–35. [Google Scholar]
- Tsuge, K.; Ano, T.; Hirai, M.; Nakamura, Y.; Shoda, M. The Genes DegQ, Pps, and Lpa-8 (Sfp) Are Responsible for Conversion of Bacillus subtilis 168 to Plipastatin Production. Antimicrob. Agents Chemother. 1999, 43, 2183–2192. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, D.; Wei, G.; Shao, N. Enhanced Co-Production of S-Adenosylmethionine and Glutathione by an ATP-Oriented Amino Acid Addition Strategy. Bioresour. Technol. 2012, 107, 19–24. [Google Scholar] [CrossRef]
- Sip, A.; Grajek, W.; Boyaval, P. Enhancement of Bacteriocin Production by Carnobacterium divergens AS7 in the Presence of a Bacteriocin-Sensitive Strain Carnobacterium piscicola. Int. J. Food Microbiol. 1998, 42, 63–69. [Google Scholar] [CrossRef]
- Leveau, J.H.J.; Preston, G.M. Bacterial Mycophagy: Definition and Diagnosis of a Unique Bacterial-Fungal Interaction. New Phytol. 2008, 177, 859–876. [Google Scholar]
- Singh, S.; Nimse, S.B.; Mathew, D.E.; Dhimmar, A.; Sahastrabudhe, H.; Gajjar, A.; Ghadge, V.A.; Kumar, P.; Shinde, P.B. Microbial Melanin: Recent Advances in Biosynthesis, Extraction, Characterization, and Applications. Biotechnol. Adv. 2021, 53, 107773. [Google Scholar]
- Chernin, L.; Ismailov, Z.; Haran, S.; Chet, I. Chitinolytic Enterobacter agglomerans Antagonistic to Fungal Plant Pathogens. Appl. Environ. Microbiol. 1995, 61, 1720–1726. [Google Scholar] [CrossRef]
- Budi, S.W.; Van Tuinen, D.; Arnould, C.; Dumas-Gaudot, E.; Gianinazzi-Pearson, V.; Gianinazzi, S. Hydrolytic Enzyme Activity of Paenibacillus Sp. Strain B2 and Effects of the Antagonistic Bacterium on Cell Integrity of Two Soil-Borne Pathogenic Fungi. Appl. Soil Ecol. 2000, 15, 191–199. [Google Scholar] [CrossRef]
- De Weert, S.; Kuiper, I.; Lagendijk, E.L.; Lamers, G.E.M.; Lugtenberg, B.J.J. Role of Chemotaxis Toward Fusaric Acid in Colonization of Hyphae of Fusarium oxysporum f. Sp. Radicis-Lycopersici by Pseudomonas fluorescens WCS365. Mol. Plant-Microbe Interact. 2004, 17, 1185–1191. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Inductors | Concentration |
|---|---|---|
| T1 | Glutamic acid | 4 g L−1 |
| T2 | Without inducer | - |
| T3 | Iron | 0.3 mg L−1 |
| T4 | Inactive cells of Colletotrichum sp. | mL−1 (1 × 106) |
| T5 | Cellulose | 20 g L−1 |
| T6 | Chitin | 4 g L−1 |
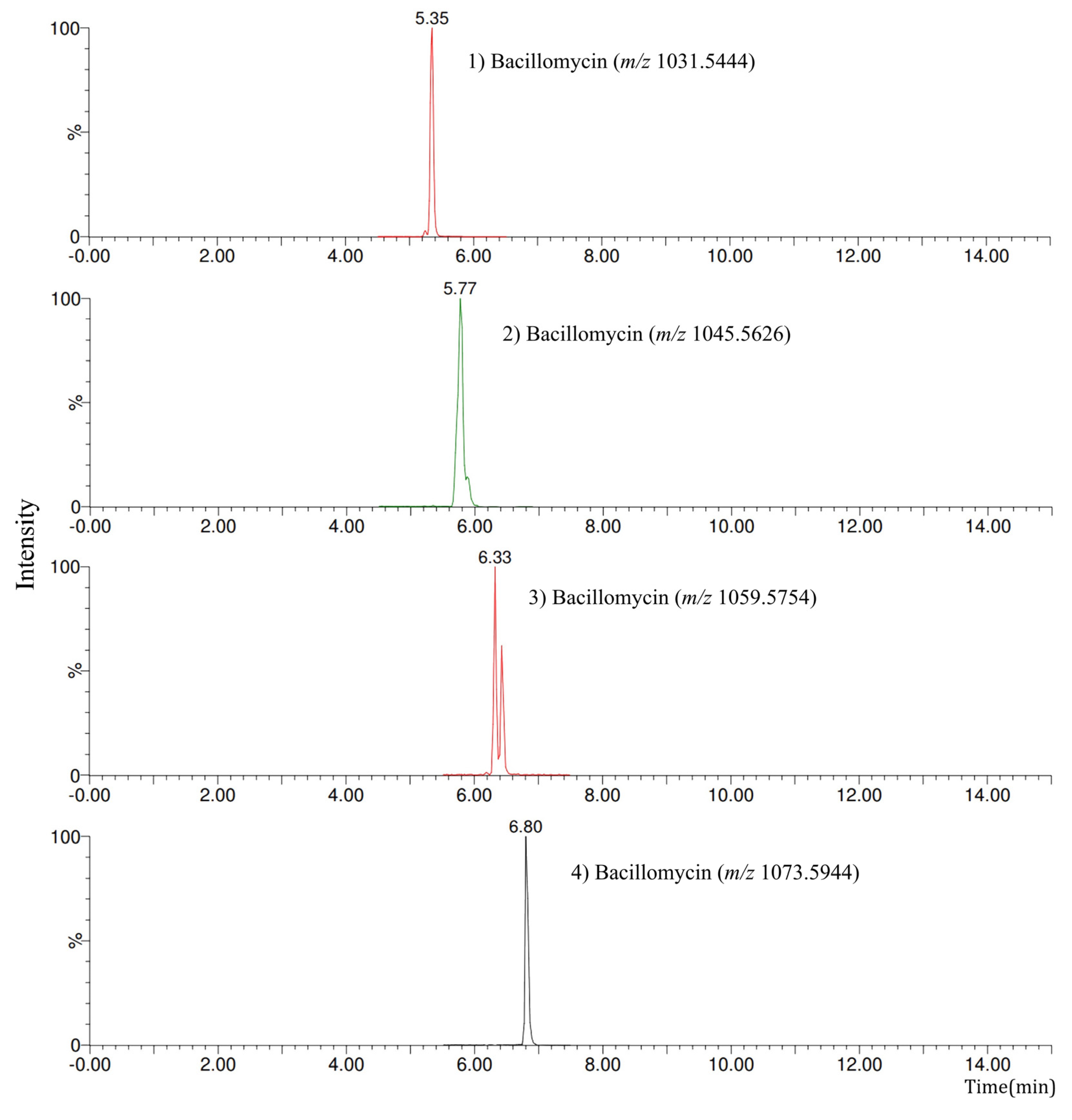
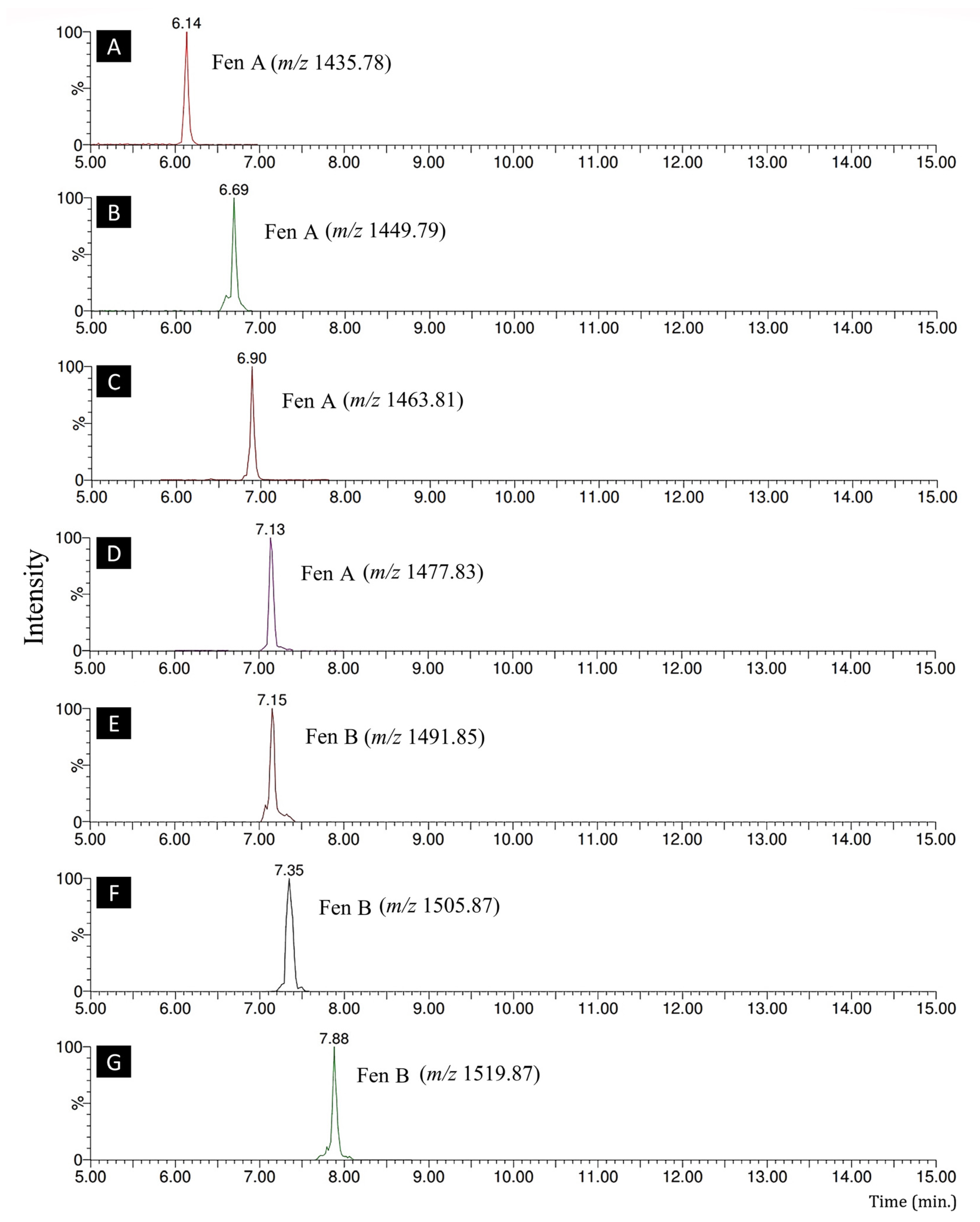
| LPs (Family) | Homologues | Retention Times (min) | Exact Mass [M + H]+ | Fragments |
|---|---|---|---|---|
| Iturin | Bacillomycin | 5.35 | 1031.54 | 227.1028, 754.4310, 978.4970, 1014.5230, 1031.5444 |
| Bacillomycin | 5.77 | 1045.56 | 227.1028, 768.4553, 992.5057, 1011.5144, 1028.5370, 1045.5626 | |
| Bacillomycin | 6.33 | 1059.57 | 227.1028, 782.4656, 1007.5164, 1025.5273, 1042.5546, 1059.5754 | |
| Bacillomycin | 6.80 | 1073.59 | 227.0997, 796.4755, 1020.5471, 1039.5442, 1073.5944 | |
| Fengycin | Fengycin A | 6.14 | 1435.78 | 966.4698, 1080.5443, 1435.7811 |
| Fengycin A | 6.69 | 1449.79 | 966.4698, 1080.5442, 1449.7935 | |
| Fengycin A | 6.90 | 1463.81 | 351.1653, 966.4698, 1080.5443, 1463.8191 | |
| Fengycin A | 7.13 | 1477.83 | 966.4507, 1080.5443, 1477.8335 | |
| Fengycin B | 7.15 | 1491.85 | 994.4919, 1108.57, 1491.8516 | |
| Fengycin B | 7.35 | 1505.87 | 994.4919, 1108.57, 1505.8722 | |
| Fengycin B | 7.88 | 1519.87 | 349.1362, 994.4919, 1108.57, 1519.8787 | |
| Surfactin | Surfactin | 10.49 | 994.64 | 373.1229, 441.2964, 653.4440, 685.4539, 976.6290, 994.6469 |
| Surfactin | 10.84 | 1008.65 | 441.2964, 667.4569, 685.4431, 990.6501, 1008.6597 | |
| Surfactin | 11.44 | 1022.67 | 441.2681, 685.4485, 1004.6674, 1022.6791 | |
| Surfactin | 11.76 | 1036.68 | 441.2645, 685.4485, 1018.6786, 1036.6897 | |
| Surfactin | 12.36 | 1050.70 | 441.2650, 685.4539, 1032.6882, 1050.7029 |
| Inducer Treatment | Bacillomycin (1031.54) | Bacillomycin (1045.55) | Bacillomycin (1059.57) | Bacillomycin (1073.58) |
|---|---|---|---|---|
| Glutamic acid | 39.4 ± 4.09 c,d | 0.1 ± 0.01 c | 8.5 ± 1.09 b,c | 4.6 ± 0.64 b |
| Mly B17 * | 54.3 ± 3.97 b | 0.1 ± 0.02 c | 16.6 ± 1.81 b | 3.7 ± 0.69 b |
| Iron | 32.5 ± 3.13 d | 0.2 ± 0.06 b | 6.7 ± 0.87 c | 3.6 ± 0.56 b |
| Inactive cells | 173.1 ± 7.14 a | 0.4 ± 0.05 a | 108.5 ± 3.80 a | 10.7 ± 1.97 a |
| Cellulose | 37.5 ± 1.99 c,d | 0.1 ± 0.02 c | 8.4 ± 1.86 b,c | 2.8 ± 0.20 b |
| Chitin | 43.4 ± 3.56 c | 0.1 ± 0.01 c | 14.1 ± 0.13 b,c | 2.7 ± 0.49 b |
| Inducer Treatment | Fen A (1435.79) | Fen A (1449.79) | Fen A (1463.79) | Fen A (1477.79) | Fen B (1491.79) | Fen B (1505.79) | Fen B (1519.9) |
|---|---|---|---|---|---|---|---|
| Glutamic acid | 94 ± 12.8 b | 53 ± 8.2 c,d | 656 ± 111.7 b | 143 ± 30.0 b | 54 ± 10.9 c | 13 ± 1.5 b | 33 ± 2.6 c,d |
| Mly B17 * | 74 ± 10.9 b | 146 ± 40.2 b | 749 ± 191.3 b | 134 ± 30.2 b | 128 ± 21.1 ab | 18 ± 4.0 b | 28 ± 6.1 c,d |
| Iron | 74 ± 12.0 b | 35 ± 8.8 d | 753 ± 99.5 b | 115 ± 20.2 b | 46 ± 10.7 c | 17 ± 2.8 b | 35 ± 6.6 c |
| Inactive cells | 350 ± 47.0 a | 216 ± 32.2 a | 3288 ± 528.5 a | 385 ± 50.0 a | 168 ± 34.5 a | 45 ± 5.5 a | 182 ± 3.8 a |
| Cellulose | 45 ± 7.0 b | 96 ± 7.5 b,c | 427 ± 105.3 b | 84 ± 19.6 b | 100 ± 6.0 b,c | 11 ± 3.7 b | 22 ± 2.1 d |
| Chitin | 92 ± 17.5 b | 112 ± 9.1 b,c | 756 ± 112.6 b | 146 ± 37.7 b | 97 ± 20.5 b,c | 13 ± 2.8 b | 59 ± 3.7 b |
| Inducer Treatment | Surfactin (994.65) | Surfactin (1008.65) | Surfactin (1022.66) | Surfactin (1036.68) | Surfactin (1050.69) |
|---|---|---|---|---|---|
| Glutamic acid | 0.8 ± 0.09 b | 1.5 ± 0.16 b | 15.8 ± 3.02 b | 27.8 ± 1.94 b | 0.3 ± 0.03 b |
| Mly B17 * | 0.3 ± 0.02 b | 0.9 ± 0.06 b,c | 6.6 ± 0.92 c,d | 21.6 ± 3.01 b,c | 0.4 ± 0.02 b |
| Iron | 0.6 ± 0.08 b | 1.2 ± 0.15 b,c | 12.0 ± 2.67 b,c | 18.6 ± 2.66 c,d | 0.4 ± 0.24 b |
| Inactive cells | 4.2 ± 0.80 a | 7.2 ± 0.82 a | 42.5 ± 3.85 a | 61.5 ± 3.01 a | 1.5 ± 0.28 a |
| Cellulose | 0.2 ± 0.01 b | 0.5 ± 0.09 c | 4.2 ± 0.79 d | 13.8 ± 2.01 d | 0.2 ± 0.03 b |
| Chitin | 0.3 ± 0.09 b | 0.6 ± 0.04 b,c | 6.0 ± 1.06 c,d | 15.0 ± 1.99 d | 0.2 ± 0.02 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ley-López, N.; Heredia, J.B.; Martín-Hernández, C.S.; Cruz-Lachica, I.; Márquez-Zequera, I.; Medina-López, R.; García-Estrada, R.S. Identification and Quantification of Lipopeptide Homologues Induced and Produced by Bacillus amyloliquefaciens. Fermentation 2023, 9, 944. https://doi.org/10.3390/fermentation9110944
Ley-López N, Heredia JB, Martín-Hernández CS, Cruz-Lachica I, Márquez-Zequera I, Medina-López R, García-Estrada RS. Identification and Quantification of Lipopeptide Homologues Induced and Produced by Bacillus amyloliquefaciens. Fermentation. 2023; 9(11):944. https://doi.org/10.3390/fermentation9110944
Chicago/Turabian StyleLey-López, Nancy, José Basilio Heredia, Cesar San Martín-Hernández, Isabel Cruz-Lachica, Isidro Márquez-Zequera, Raymundo Medina-López, and Raymundo Saúl García-Estrada. 2023. "Identification and Quantification of Lipopeptide Homologues Induced and Produced by Bacillus amyloliquefaciens" Fermentation 9, no. 11: 944. https://doi.org/10.3390/fermentation9110944
APA StyleLey-López, N., Heredia, J. B., Martín-Hernández, C. S., Cruz-Lachica, I., Márquez-Zequera, I., Medina-López, R., & García-Estrada, R. S. (2023). Identification and Quantification of Lipopeptide Homologues Induced and Produced by Bacillus amyloliquefaciens. Fermentation, 9(11), 944. https://doi.org/10.3390/fermentation9110944