
Phages in Fermented Foods: Interactions and Applications



Abstract
:1. Introduction
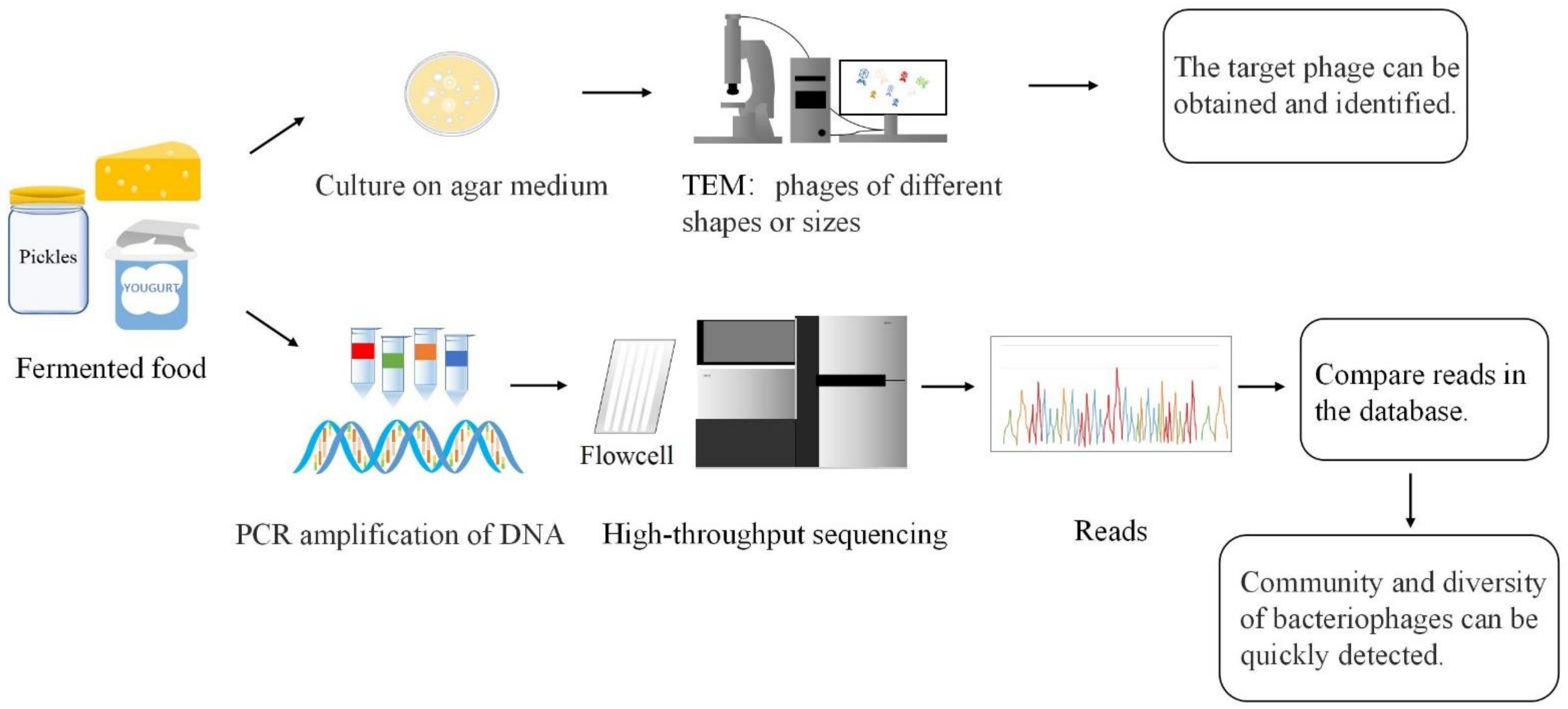
2. Phage Detection
2.1. Cultivation Methods
2.2. The Culture-Free Method
2.3. Diversity of Phages in Fermented Foods
3. Interactions between Bacteriophages and Fermenting Microorganisms
3.1. Genetic Basis of Lactic Acid Bacteria (LAB) Phages
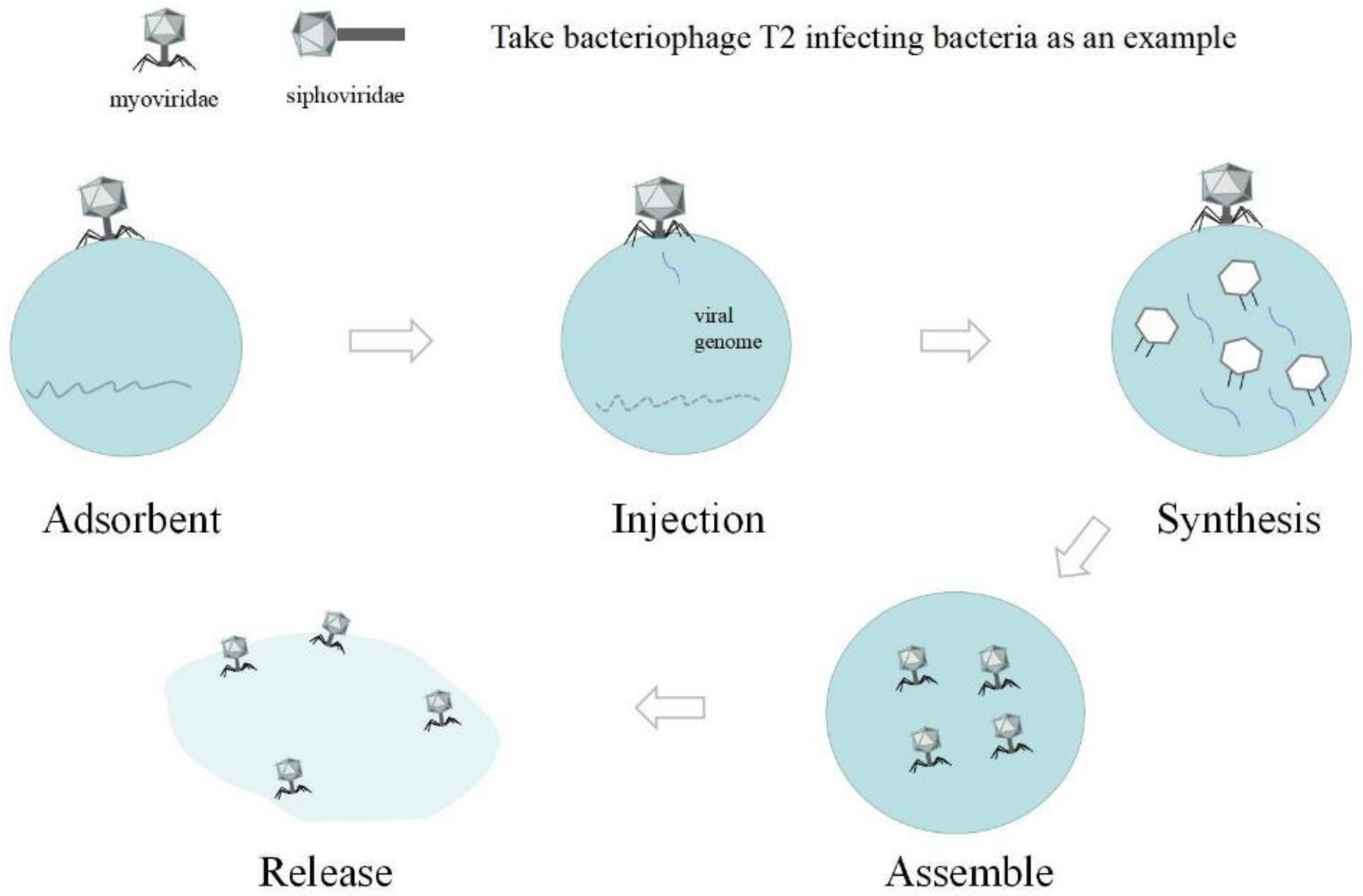
3.2. Mechanism of Phage Binding to Receptors
3.3. Phage Resistance Mechanisms
3.3.1. Preventing Phage Adsorption
3.3.2. Preventing Phage DNA Entry
3.3.3. Cutting Phage Nucleic Acids
3.3.4. CRISPR-Based Prokaryotic Adaptive Immune System (CRISPR-Associated System, Cas)
3.3.5. Abortive Infection Systems (Abi)
3.4. Positive Effects of Phages on the Fermentation Microbial Community
3.5. Negative Effects of Bacteriophages on Fermenting Microorganisms
4. Application of Bacteriophages in Fermented Food
4.1. Bacteriophages as an Ideal Antibacterial Agent
4.2. Bacterial Mutants Obtained after Bacteriophage Treatment as Starters for Fermented Food
5. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| GRAS | generally recognized as safe |
| TEM | Transmission Electron Microscope |
| PG | cell wall peptidoglycan |
| MAB | mesophilic anaerobe |
| HHP | high hydrostatic pressure |
References
- Gill, J.J.; Hyman, P. Phage Choice, Isolation, and Preparation for Phage Therapy. Curr. Pharm. Biotechnol. 2010, 11, 2–14. [Google Scholar] [CrossRef]
- Wilkinson, L. William C Summers, Félix d’Herelle and the origins of molecular biology, New Haven and London, Yale University Press, 1999, pp. xii, 230, £20.00 (hardback 0-300-07127-2). Med. Hist. 2001, 45, 294–295. [Google Scholar] [CrossRef] [Green Version]
- Eiben, S.; Koch, C.; Altintoprak, K.; Southan, A.; Tovar, G.; Laschat, S.; Weiss, I.M.; Wege, C. Plant virus-based materials for biomedical applications: Trends and prospects. Adv. Drug Deliv. Rev. 2018, 145, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.J. The bionanoscience of plant viruses: Templates and synthons for new materials. J. Mater. Chem. 2008, 18, 3746–3754. [Google Scholar] [CrossRef]
- Hudson, J.A.; Billington, C.; Carey-Smith, G.; Greening, G. Bacteriophages as Biocontrol Agents in Food. J. Food Prot. 2005, 68, 426–437. [Google Scholar] [CrossRef]
- Yang, M.; Mao, C. Biomaterials based on phages and other viruses. Adv. Drug Deliv. Rev. 2019, 145, 1–3. [Google Scholar] [CrossRef]
- Kim, B.O.; Kim, E.S.; Yoo, Y.J.; Bae, H.W.; Chung, I.Y.; Cho, Y.H. Phage-Derived Antibacterials: Harnessing the Simplicity, Plasticity, and Diversity of Phages. Viruses 2019, 11, 268. [Google Scholar] [CrossRef] [Green Version]
- Loc-Carrillo, C.; Abedon, S.T. Pros and cons of phage therapy. Bacteriophage 2011, 1, 111–114. [Google Scholar] [CrossRef] [Green Version]
- Wittebole, X.; De Roock, S.; Opal, S.M. A historical overview of bacteriophage therapy as an alternative to antibiotics for the treatment of bacterial pathogens. Virulence 2014, 5, 226–235. [Google Scholar] [CrossRef]
- Hull, R. Assay, Detection, and Diagnosis of Plant Viruses. In Plant Virology; Academic Press: Cambridge, MA, USA, 2014; pp. 755–808. [Google Scholar]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus taxonomy: The database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, D708–D717. [Google Scholar] [CrossRef] [Green Version]
- Walker, P.J.; Benmansour, A.; Dietzgen, R. Family Rhabdoviridae. In Virus Taxonomy and Nomenclature of Viruses; Seventh Report of the International Committee on Taxonomy of Viruses; Academic Press: San Diego, CA, USA, 2000; pp. 563–583. [Google Scholar]
- Dion, M.B.; Oechslin, F.; Moineau, S. Phage diversity, genomics and phylogeny. Nat. Rev. Microbiol. 2020, 18, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.; Mohsin, M.; Ijaz, H.; Tulain, U.R.; Ashraf, M.A.; Fayyaz, A. Lactic acid bacteria in traditional fermented Asian foods. Pak. J. Pharm. Sci. 2017, 30, 1803–1814. [Google Scholar] [PubMed]
- Hong, S.W.; Choi, Y.J.; Lee, H.W.; Yang, J.H.; Lee, M.A. Microbial Community Structure of Korean Cabbage Kimchi and Ingredients with Denaturing Gradient Gel Electrophoresis. J. Microbiol. Biotechnol. 2016, 26, 1057–1062. [Google Scholar] [CrossRef] [PubMed]
- Chirkena, K.; Ulusoy, B.; Asnake, W.; Yldrm, F.K. In vitro probiotic and industrial properties of bacteria isolated from fermented food products. Int. Food Res. J. 2021, 28, 638–653. [Google Scholar]
- Haaland, R. Porridge and Pot, Bread and Oven: Food Ways and Symbolism in Africa and the Near East from the Neolithic to the Present. Camb. Archaeol. J. 2007, 17, 165–182. [Google Scholar] [CrossRef]
- Dillon, S.L.; Shapter, F.M.; Henry, R.J.; Cordeiro, G.; Izquierdo, L.; Lee, L.S. Domestication to Crop Improvement: Genetic Resources for Sorghum and Saccharum (Andropogoneae). Ann. Bot. 2007, 100, 975–989. [Google Scholar] [CrossRef] [Green Version]
- Hsiung, R.T.; Fang, W.T.; Lepage, B.A.; Hsu, S.A.; Hsu, C.H.; Chou, J.Y. In Vitro Properties of Potential Probiotic Indigenous Yeasts Originating from Fermented Food and Beverages in Taiwan. Probiotics Antimicrob. Proteins 2021, 13, 113–124. [Google Scholar] [CrossRef]
- Rezac, S.; Kok, C.R.; Heermann, M.; Hutkins, R. Fermented Foods as a Dietary Source of Live Organisms. Front. Microbiol. 2018, 9, 1785. [Google Scholar] [CrossRef]
- Chaïb, A.; Philippe, C.; Jaomanjaka, F.; Barchi, Y.; Oviedo-Hernandez, F.; Claisse, O.; Le Marrec, C. Phage-host interactions as a driver of population dynamics during wine fermentation: Betting on underdogs. Int. J. Food Microbiol. 2022, 383, 109936. [Google Scholar] [CrossRef]
- Bertozzi Silva, J.; Sauvageau, D. Bacteriophages as antimicrobial agents against bacterial contaminants in yeast fermentation processes. Biotechnol. Biofuels 2014, 7, 123. [Google Scholar] [CrossRef]
- Bueno, E.; Garcia, P.; Martinez, B.; Rodríguez, A. Phage inactivation of Staphylococcus aureus in fresh and hard-type cheeses. Int. J. Food Microbiol. 2012, 158, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Tofalo, R.; Fusco, V.; Bohnlein, C.; Kabisch, J.; Logrieco, A.F.; Habermann, D.; Cho, G.S.; Benomar, N.; Abriouel, H.; Schmidt-Heydt, M.; et al. The life and times of yeasts in traditional food fermentations. Crit. Rev. Food Sci. Nutr. 2020, 60, 3103–3132. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Kim, K.H.; Abell, G.C.; Kim, M.S.; Roh, S.W.; Bae, J.W. Metagenomic Analysis of the Viral Communities in Fermented Foods. Appl. Environ. Microbiol. 2011, 77, 1284–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, K.; Kang, H.S.; Hyun, W.B.; Kim, K.P. High prevalence of Bacillus subtilis-infecting bacteriophages in soybean-based fermented foods and its detrimental effects on the process and quality of Cheonggukjang. Food Microbiol. 2018, 76, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Schuster, H. Bacteriophages, von MH Adams; Interscience Publishers, Inc.: New York, NY, USA, 1959; p. 164. [Google Scholar]
- Blanch, A.R.; Lucena, F.; Muniesa, M.; Jofre, J. Fast and easy methods for the detection of coliphages. J. Microbiol. Methods 2020, 173, 105940. [Google Scholar] [CrossRef]
- Levine, M. Mutations in the temperate phage P22 and lysogeny in Salmonella. Virology 1957, 3, 22–41. [Google Scholar] [CrossRef] [PubMed]
- Jebri, S.; Hmaied, F.; Yahya, M.; Ben Ammar, A.; Hamdi, M. Total coliphages removal by activated sludge process and their morphological diversity by transmission electron microscopy. Water Sci. Technol. 2016, 74, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.D.; Park, J.H. Genomic analysis of WCP30 Phage of Weissella cibaria for Dairy Fermented Foods. Korean J. Food Sci. Anim. Resour. 2017, 37, 884–888. [Google Scholar]
- Kim, E.J.; Hong, J.W.; Yun, N.R.; Lee, Y.N. Characterization of Bacillus phage-K2 isolated from chungkookjang, a fermented soybean foodstuff. J. Ind. Microbiol. Biotechnol. 2011, 38, 39–42. [Google Scholar] [CrossRef]
- Shin, H.; Bandara, N.; Shin, E.; Ryu, S.; Kim, K.P. Prevalence of Bacillus cereus bacteriophages in fermented foods and characterization of phage JBP901. Res. Microbiol. 2011, 162, 791–797. [Google Scholar] [CrossRef]
- Sunthornthummas, S.; Doi, K.; Rangsiruji, A.; Sarawaneeyaruk, S.; Pringsulaka, O. Isolation and characterization of Lactobacillus paracasei LPC and phage Phi T25 from fermented milk. Food Control 2017, 73, 1353–1361. [Google Scholar] [CrossRef]
- Rajnovic, D.; Mas, J. Fluorometric detection of phages in liquid media: Application to turbid samples. Anal. Chim. Acta 2020, 1111, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Romero, D.A.; Magill, D.; Millen, A.; Horvath, P.; Fremaux, C. Dairy lactococcal and streptococcal phage-host interactions: An industrial perspective in an evolving phage landscape. FEMS Microbiol. Rev. 2020, 44, 909–932. [Google Scholar] [CrossRef] [PubMed]
- Paillet, T.; Dugat-Bon, E. Bacteriophage ecology of fermented foods: Anything new under the sun? Curr. Opin. Food Sci. 2021, 40, 102–111. [Google Scholar] [CrossRef]
- Zünd, M.; Ruscheweyh, H.J.; Field, C.M.; Meyer, N.; Cuenca, M.; Hoces, D.; Hardt, W.D.; Sunagawa, S. High throughput sequencing provides exact genomic locations of inducible prophages and accurate phage-to-host ratios in gut microbial strains. Microbiome 2021, 9, 77. [Google Scholar] [CrossRef]
- Sprotte, S.; Fagbemigun, O.; Brinks, E.; Cho, G.S.; Casey, E.; Oguntoyinbo, F.A.; Neve, H.; Mahony, J.; van Sinderen, D.; Franz, C. Novel Siphoviridae phage PMBT4 belonging to the group b Lactobacillus delbrueckii subsp. bulgaricus phages. Virus Res. 2022, 308, 198635. [Google Scholar] [CrossRef]
- Zheng, X.-F.; Yang, Z.-Q.; Zhang, H.; Jin, W.X.; Xu, C.W.; Gao, L.; Rao, S.Q.; Jiao, X.A. Isolation of virulent phages infecting dominant mesophilic aerobic bacteria in cucumber pickle fermentation. Food Microbiol. 2020, 86, 103330. [Google Scholar] [CrossRef]
- Komora, N.; Bruschi, C.; Ferreira, V.; Maciel, C.; Brandão, T.R.S.; Fernandes, R.; Saraiva, J.A.; Castro, S.M.; Teixeira, P. The protective effect of food matrices on Listeria lytic bacteriophage P100 application towards high pressure processing. Food Microbiol. 2018, 76, 416–425. [Google Scholar] [CrossRef]
- Lavelle, K.; Murphy, J.; Fitzgerald, B.; Lugli, G.A.; Zomer, A.; Neve, H.; Ventura, M.; Franz, C.M.; Cambillau, C.; van Sinderen, D.; et al. A Decade of Streptococcus thermophilus Phage Evolution in an Irish Dairy Plant. Appl. Environ. Microbiol. 2018, 84, e02855-17. [Google Scholar] [CrossRef] [Green Version]
- Sunthornthummas, S.; Doi, K.; Fujino, Y.; Rangsiruji, A.; Sarawaneeyaruk, S.; Insian, K.; Pringsulaka, O. Genomic characterisation of Lacticaseibacillus paracasei phage ΦT25 and preliminary analysis of its derived endolysin. Int. Dairy J. 2021, 116, 104968. [Google Scholar] [CrossRef]
- Umene, K.; Shiraishi, A. Complete nucleotide sequence of Bacillus subtilis (natto) bacteriophage PM1, a phage associated with disruption of food production. Virus Genes 2013, 46, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Pérez-Díaz, I.M.; Hayes, J.S.; Breidt, F. Bacteriophage ecology in a commercial cucumber fermentation. Appl. Environ. Microbiol. 2012, 78, 8571–8578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez, M.; Velázquez, R.; Maqueda, M.; López-Piñeiro, A.; Ribas, J.C. A new wine Torulaspora delbrueckii killer strain with broad antifungal activity and its toxin-encoding double-stranded RNA virus. Front. Microbiol. 2015, 6, 983. [Google Scholar] [CrossRef] [Green Version]
- De Lima, C.P.; Bruno, L.M.; de Figueiredo, E.A.T.; del Lujan Quiberoni, A.; Carvalho, J.D.G.; de Carvalho, A.K.F. Phage resistance of acid lactic bacteria isolated from Coalho cheese industries. Cienc. Rural 2012, 42, 1117–1122. [Google Scholar]
- Spinelli, S.; Tremblay, D.; Moineau, S.; Cambillau, C.; Goulet, A. Structural Insights into Lactococcal Siphophage p2 Baseplate Activation Mechanism. Viruses 2020, 12, 878. [Google Scholar] [CrossRef] [PubMed]
- Lanza, B.; Zago, M.; Carminati, D.; Rossetti, L.; Meucci, A.; Marfisi, P.; Russi, F.; Iannucci, E.; Di Serio, M.G.; Giraffa, G. Isolation and preliminary characterization of Lactobacillus plantarum bacteriophages from table olive fermentation. Ann. Microbiol. 2012, 62, 1467–1472. [Google Scholar] [CrossRef]
- Kelly, W.J.; Altermann, E.; Lambie, S.C.; Leahy, S.C. Interaction between the genomes of Lactococcus lactis and phages of the P335 species. Front. Microbiol. 2013, 4, 257. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, Z.; Abedon, S.T. Diversity of phage infection types and associated terminology: The problem with ‘Lytic or lysogenic’. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef] [Green Version]
- Sechaud, L.; Cluzel, P.J.; Rousseau, M.; Baumgartner, A.; Accolas, J.P. Bacteriophages of lactobacilli. Biochimie 1988, 70, 401–410. [Google Scholar] [CrossRef]
- Casey, E.; Mahony, J.; Neve, H.; Noben, J.P.; Dal Bello, F.; van Sinderen, D. Novel phage group infecting Lactobacillus delbrueckii subsp. lactis, as revealed by genomic and proteomic analysis of bacteriophage Ldl1. Appl. Environ. Microbiol. 2015, 81, 1319–1326. [Google Scholar] [CrossRef] [Green Version]
- Desiere, F.; Lucchini, S.; Canchaya, C.; Ventura, M.; Brüssow, H. Comparative genomics of phages and prophages in lactic acid bacteria. Antonie Van Leeuwenhoek 2002, 82, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Mahony, J.; Cambillau, C.; Van Sinderen, D. Host recognition by lactic acid bacterial phages. FEMS Microbiol. Rev. 2017, 41 (Suppl. S1), S16–S26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durmaz, E.; Klaenhammer, T.R. Genetic analysis of chromosomal regions of Lactococcus lactis acquired by recombinant lytic phages. Appl. Environ. Microbiol. 2000, 66, 895–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botstein, D. A theory of modular evolution for bacteriophages. Ann. N. Y. Acad. Sci. 1980, 354, 484–490. [Google Scholar] [CrossRef]
- Hendrix, R.W.; Smith, M.C.; Burns, R.N.; Ford, M.E.; Hatfull, G.F. Evolutionary relationships among diverse bacteriophages and prophages: All the world’s a phage. Proc. Natl. Acad. Sci. USA 1999, 96, 2192–2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, K.; Vogensen, F.K.; Neve, H.; Bresciani, J.; Josephsen, J. Identification of the receptor-binding protein in 936-species lactococcal bacteriophages. Appl. Environ. Microbiol. 2004, 70, 5818–5824. [Google Scholar] [CrossRef] [Green Version]
- Moineau, S.; Pandian, S.; Klaenhammer, T.R. Evolution of a Lytic Bacteriophage via DNA Acquisition from the Lactococcus lactis Chromosome. Appl. Environ. Microbiol. 1994, 60, 1832–1841. [Google Scholar] [CrossRef] [Green Version]
- Zago, M.; Scaltriti, E.; Fornasari, M.E.; Rivetti, C.; Grolli, S.; Giraffa, G.; Ramoni, R.; Carminati, D. Epifluorescence and atomic force microscopy: Two innovative applications for studying phage-host interactions in Lactobacillus helveticus. J. Microbiol. Methods 2012, 88, 41–46. [Google Scholar] [CrossRef]
- Vodzinska, N.; Galkin, B.; Ishkov, Y.; Kirichenko, A.; Kondratyuk, A.; Filipova, T. Inhibition of lactophage activity by quinolinilporphyrin and its zinc compex. Pol. J. Microbiol. 2011, 60, 229–232. [Google Scholar] [CrossRef]
- Qimron, U.; Marintcheva, B.; Tabor, S.; Richardson, C.C. Genomewide screens for Escherichia coli genes affecting growth of T7 bacteriophage. Proc. Natl. Acad. Sci. USA 2006, 103, 19039–19044. [Google Scholar] [CrossRef] [Green Version]
- Mahony, J.; Ainsworth, S.; Stockdale, S.; van Sinderen, D. Phages of lactic acid bacteria: The role of genetics in understanding phage-host interactions and their co-evolutionary processes. Virology 2012, 434, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batra, N.G.; Sharma, A.; Agarwal, N. Evaluation of microbiological criteria and quality of packed fruit juices. Int. Food Res. J. 2018, 25, 458–461. [Google Scholar]
- Ledormand, P.; Desmasures, N.; Bernay, B.; Goux, D.; Rué, O.; Midoux, C.; Monnet, C.; Dalmasso, M. Molecular approaches to uncover phage-lactic acid bacteria interactions in a model community simulating fermented beverages. Food Microbiol. 2022, 107, 104069. [Google Scholar] [CrossRef]
- Mahony, J.; Mcdonnell, B.; Casey, E.; van Sinderen, D. Phage-Host Interactions of Cheese-Making Lactic Acid Bacteria. Annu. Rev. Food Sci. Technol. 2016, 7, 267–285. [Google Scholar] [CrossRef]
- Farenc, C.; Spinelli, S.; Vinogradov, E.; Tremblay, D.; Blangy, S.; Sadovskaya, I.; Moineau, S.; Cambillau, C. Molecular insights on the recognition of a Lactococcus lactis cell wall pellicle by the phage 1358 receptor binding protein. J. Virol. 2014, 88, 7005–7015. [Google Scholar] [CrossRef] [Green Version]
- Chaturongakul, S.; Ounjai, P. Phage-host interplay: Examples from tailed phages and Gram-negative bacterial pathogens. Front. Microbiol. 2014, 5, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapot-Chartier, M.P. Interactions of the cell-wall glycopolymers of lactic acid bacteria with their bacteriophages. Front. Microbiol. 2014, 5, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymczak, P.; Filipe, S.R.; Covas, G.; Vogensen, F.K.; Neves, A.R.; Janzen, T. Cell Wall Glycans Mediate Recognition of the Dairy Bacterium Streptococcus thermophilus by Bacteriophages. Appl. Environ. Microbiol. 2018, 84, e01847-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcelli, B.; De Jong, A.; Karsens, H.; Janzen, T.; Kok, J.; Kuipers, O.P. A Specific Sugar Moiety in the Lactococcus lactis Cell Wall Pellicle Is Required for Infection by CHPC971, a Member of the Rare 1706 Phage Species. Appl. Environ. Microbiol. 2019, 85, e01224-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapot-Chartier, M.P.; Vinogradov, E.; Sadovskaya, I.; Andre, G.; Mistou, M.Y.; Trieu-Cuot, P.; Furlan, S.; Bidnenko, E.; Courtin, P.; Péchoux, C.; et al. Cell surface of Lactococcus lactis is covered by a protective polysaccharide pellicle. J. Biol. Chem. 2010, 285, 10464–10471. [Google Scholar] [CrossRef] [Green Version]
- Lavelle, K.; Sadovskaya, I.; Vinogradov, E.; Kelleher, P.; Lugli, G.A.; Ventura, M.; van Sinderen, D.; Mahony, J. Brussowvirus SW13 Requires a Cell Surface-Associated Polysaccharide To Recognize Its Streptococcus thermophilus Host. Appl. Environ. Microbiol. 2022, 88, e0172321. [Google Scholar] [CrossRef] [PubMed]
- Arguijo-Hernández, E.S.; Hernandez-Sanchez, J.; Briones-Peña, S.J.; Oviedo, N.; Mendoza-Hernández, G.; Guarneros, G.; Kameyama, L. Cor interacts with outer membrane proteins to exclude FhuA-dependent phages. Arch. Virol. 2018, 163, 2959–2969. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Capalash, N.; Sharma, P. Restriction endonucleases: Natural and directed evolution. Appl. Microbiol. Biotechnol. 2012, 94, 583–599. [Google Scholar] [CrossRef] [PubMed]
- Bertozzi Silva, J.; Storms, Z.; Sauvageau, D. Host receptors for bacteriophage adsorption. FEMS Microbiol. Lett. 2016, 363, fnw002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medhekar, B.; Miller, J.F. Diversity-generating retroelements. Curr. Opin. Microbiol. 2007, 10, 388–395. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, I.W.; Hughes, K.A.; Skillman, L.C.; Tait, K. The interaction of phage and biofilms. FEMS Microbiol. Lett. 2004, 232, 1–6. [Google Scholar] [CrossRef]
- Liu, M.; Deora, R.; Doulatov, S.R.; Gingery, M.; Eiserling, F.A.; Preston, A.; Maskell, D.J.; Simons, R.W.; Cotter, P.A.; Parkhill, J.; et al. Reverse transcriptase-mediated tropism switching in Bordetella bacteriophage. Science 2002, 295, 2091–2094. [Google Scholar] [CrossRef]
- Destoumieux-Garzón, D.; Duquesne, S.; Peduzzi, J.; Goulard, C.; Desmadril, M.; Letellier, L.; Rebuffat, S.; Boulanger, P. The iron-siderophore transporter FhuA is the receptor for the antimicrobial peptide microcin J25: Role of the microcin Val11-Pro16 beta-hairpin region in the recognition mechanism. Biochem. J. 2005, 389, 869–876. [Google Scholar] [CrossRef] [Green Version]
- Moak, M.; Molineux, I.J. Role of the Gp16 lytic transglycosylase motif in bacteriophage T7 virions at the initiation of infection. Mol. Microbiol. 2000, 37, 345–355. [Google Scholar] [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Uc-Mass, A.; Loeza, E.J.; De La Garza, M.; Guarneros, G.; Hernández-Sánchez, J.; Kameyama, L. An orthologue of the cor gene is involved in the exclusion of temperate lambdoid phages. Evidence that Cor inactivates FhuA receptor functions. Virology 2004, 329, 425–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tock, M.R.; Dryden, D.T. The biology of restriction and anti-restriction. Curr. Opin. Microbiol. 2005, 8, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.J.; Belfort, M.; Bestor, T.; Bhagwat, A.S.; Bickle, T.A.; Bitinaite, J.; Blumenthal, R.M.; Degtyarev, S.; Dryden, D.T.; Dybvig, K.; et al. A nomenclature for restriction enzymes, DNA methyltransferases, homing endonucleases and their genes. Nucleic Acids Res. 2003, 31, 1805–1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Attar, S.; Westra, E.R.; Van Der Oost, J.; Brouns, S.J. Clustered regularly interspaced short palindromic repeats (CRISPRs): The hallmark of an ingenious antiviral defense mechanism in prokaryotes. Biol. Chem. 2011, 392, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Hille, F.; Richter, H.; Wong, S.P.; Bratovič, M.; Ressel, S.; Charpentier, E. The Biology of CRISPR-Cas: Backward and Forward. Cell 2018, 172, 1239–1259. [Google Scholar] [CrossRef] [Green Version]
- Rahmatabadi, S.S.; Nezafat, N.; Negahdaripour, M.; Hajighahramani, N.; Morowvat, M.H.; Ghasemi, Y. Studying the features of 57 confirmed CRISPR loci in 29 strains of Escherichia coli. J. Basic Microbiol. 2016, 56, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Garneau, J.E.; Dupuis, M.E.; Villion, M.; Romero, D.A.; Barrangou, R.; Boyaval, P.; Fremaux, C.; Horvath, P.; Magadán, A.H.; Moineau, S. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature 2010, 468, 67–71. [Google Scholar] [CrossRef]
- Zinno, P.; Janzen, T.; Bennedsen, M.; Ercolini, D.; Mauriello, G. Characterization of Streptococcus thermophilus lytic bacteriophages from mozzarella cheese plants. Int. J. Food Microbiol. 2010, 138, 137–144. [Google Scholar] [CrossRef]
- Ma, C.; Ma, A.; Gong, G.; Liu, Z.; Wu, Z.; Guo, B.; Chen, Z. Cracking Streptococcus thermophilus to stimulate the growth of the probiotic Lactobacillus casei in co-culture. Int. J. Food Microbiol. 2015, 210, 42–46. [Google Scholar] [CrossRef]
- Komora, N.; Maciel, C.; Pinto, C.A.; Ferreira, V.; Brandão, T.R.S.; Saraiva, J.M.A.; Castro, S.M.; Teixeira, P. Non-thermal approach to Listeria monocytogenes inactivation in milk: The combined effect of high pressure, pediocin PA-1 and bacteriophage P100. Food Microbiol. 2020, 86, 103315. [Google Scholar] [CrossRef]
- Cho, W.I.; Kim, E.J.; Hwang, H.J.; Cha, Y.H.; Chung, M.S.J.I.F.S.; Technologies, E. Continuous ohmic heating system for the pasteurization of fermented red pepper paste. Innov. Food Sci. Emerg. Technol. 2017, 42, 190–196. [Google Scholar] [CrossRef]
- Song, B.-S.; Park, J.-G.; Park, J.-N.; Han, I.J.; Choi, J.I.; Kim, J.H.; Byun, M.W.; Kang, S.W.; Choi, G.H.; Lee, J.W.; et al. High-dose processing and application to Korean space foods. Radiat. Phys. Chem. 2009, 78, 671–674. [Google Scholar] [CrossRef]
- Yang, F.; Zhu, L.L.; Diao, Y.D.; Gao, P.; Yu, D.W.; Yu, P.P.; Jiang, Q.X.; Xu, Y.S.; Xia, W.S.; Zhan, X.B. Preparation of High-Quality Fermented Fish Product. J. Vis. Exp. 2019, 150, e60265. [Google Scholar]
- Qian, B.J.; Deng, Y.; Xie, L.Y.; Han, Z.; Zhang, S. Physical Properties of Yoghurt at Varying Pasteurization Time and Temperature. Philipp. Agric. Sci. 2010, 93, 299–306. [Google Scholar]
- Komora, N.; Maciel, C.; Amaral, R.A.; Fernandes, R.; Castro, S.M.; Saraiva, J.A.; Teixeira, P. Innovative hurdle system towards Listeria monocytogenes inactivation in a fermented meat sausage model—High pressure processing assisted by bacteriophage P100 and bacteriocinogenic Pediococcus acidilactici. Food Res. Int. 2021, 148, 110628. [Google Scholar] [CrossRef]
- Sulakvelidze, A. Using lytic bacteriophages to eliminate or significantly reduce contamination of food by foodborne bacterial pathogens. J. Sci. Food Agric. 2013, 93, 3137–3146. [Google Scholar] [CrossRef]
- Maura, D.; Debarbieux, L. Bacteriophages as twenty-first century antibacterial tools for food and medicine. Appl. Microbiol. Biotechnol. 2011, 90, 851–859. [Google Scholar] [CrossRef]
- Rees, C.E.; Dodd, C.E. Phage for rapid detection and control of bacterial pathogens in food. Adv. Appl. Microbiol. 2006, 59, 159–186. [Google Scholar]
- Zhang, L.; Qu, M.; Yao, J.; Wang, P.; Liao, X.; Hu, X.; Chen, F. Effect of high hydrostatic pressure on the viability of Streptococcus thermophilus bacteriophages isolated from cheese. Innov. Food Sci. Emerg. Technol. 2015, 29, 113–118. [Google Scholar] [CrossRef]
- Axelsson, L.; Bjerke, G.A.; Mcleod, A.; Berget, I.; Holck, A.L. Growth Behavior of Listeria monocytogenes in a Traditional Norwegian Fermented Fish Product (Rakfisk), and Its Inhibition through Bacteriophage Addition. Foods 2020, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Tabla, R.; Gómez, A.; Rebollo, J.E.; Molina, F.; Roa, I. Effectiveness of a bacteriophage cocktail in reducing cheese early blowing caused by Escherichia coli. LWT-Food Sci. Technol. 2022, 153, 112430. [Google Scholar] [CrossRef]
- Bandara, N.; Jo, J.; Ryu, S.; Kim, K.P. Bacteriophages BCP1-1 and BCP8-2 require divalent cations for efficient control of Bacillus cereus in fermented foods. Food Microbiol. 2012, 31, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, S.; Bebeacua, C.; Orlov, I.; Tremblay, D.; Klaholz, B.P.; Moineau, S.; Cambillau, C. Cryo-electron microscopy structure of lactococcal siphophage 1358 virion. J. Virol. 2014, 88, 8900–8910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villion, M.; Moineau, S. Bacteriophages of Lactobacillus. Front. Biosci. 2009, 14, 1661–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naumenko, O.; Skrypkina, I.; Zhukova, Y.; Vakulenko, M.; Kigel, N. Selection and analysis of bacteriophage-insensitive mutants of Streptococcus thermophilus isolated in Ukraine. Int. J. Dairy Technol. 2019, 72, 515–523. [Google Scholar] [CrossRef]
- Marcó, M.B.; Zacarías, M.; Vinderola, G.; Reinheimer, J.A.; Quiberoni, A. Biological and probiotic characterisation of spontaneous phage-resistant mutants of Lactobacillus plantarum. Int. Dairy J. 2014, 39, 64–70. [Google Scholar] [CrossRef]
- Bao, Z.; Wang, X.; Wang, Q.; Zou, L.; Peng, L.; Li, L.; Tu, W.; Li, Q. A novel method of domestication combined with ARTP to improve the reduction ability of Bacillus velezensis to Cr (VI). J. Environ. Chem. Eng. 2023, 11, 109091. [Google Scholar] [CrossRef]
- Kleppen, H.P.; Holo, H.; Jeon, S.R.; Nes, I.F.; Yoon, S.S. Novel Podoviridae family bacteriophage infecting Weissella cibaria isolated from Kimchi. Appl. Environ. Microbiol. 2012, 78, 7299–7308. [Google Scholar] [CrossRef] [Green Version]
- Tu, W.; Cao, X.; Cheng, J.; Li, L.; Zhang, T.; Wu, Q.; Xiang, P.; Shen, C.; Li, Q. Chinese Baijiu: The perfect works of microorganisms. Front. Microbiol. 2022, 13, 919044. [Google Scholar] [CrossRef]
- Li, L.; Cao, X.; Zhang, T.; Wu, Q.; Xiang, P.; Shen, C.; Zou, L.; Li, Q. Recent developments in Surface-Enhanced Raman Spectroscopy and its application in food analysis: Alcoholic beverages as an example. Foods 2022, 11, 2165. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Types of Fermented Foods | The Strain (Host) Used for Phage Isolation | Phage Group | Nature | Form | References |
|---|---|---|---|---|---|
| Kimchi | Lactic acid bacteria | Podoviridae | lytic phages | Regular polyhedron head + soft tail | [40] |
| Fermented dairy products | Lactic acid bacteria | Siphoviridae | lytic phages | Hexagonal head + a long noncontracted tail | [42,43] |
| Fermented soy products | Bacillus subtilis | Spounavirinae | lytic phages | Regular icosahedron + tail | [26,44,45,46,48] |
| Cheese | Ligilactobacillus salivarius, Lactobacillus | Myoviridae | lytic phages | Not mentioned | [49,50] |
| Fermented olives | Lactiplantibacillus plantarum | Not mentioned | Not mentioned | Not mentioned | [51] |
| Fermented corn | Lactobacillus | Not mentioned | lysogenic and lytic phages | Not mentioned | [52] |
| Sterilization Method | Method | Types of Fermented Foods | Advantages and Disadvantages | References |
|---|---|---|---|---|
| Common sterilization methods | Ultrahigh-pressure sterilization technology | Pickles, yogurt | Advantages: no temperature change, can better retain the original color, taste, and nutrients Disadvantage: high energy consumption | [94,95,96,97,98] |
| Irradiation Sterilization | Kimchi | Pros: thiamine can be stored for years after irradiation without refrigeration Disadvantages: high energy consumption, vitamins E and C will reduce or even eliminate mutations that may be caused through radiation and lead to radiation resistance | ||
| Pasteurization | Fermented peppers, yogurt | Advantages: better preservation of the nutrition and natural flavor of yogurt Cons: can store at 4 °C for approximately a week | ||
| High-temperature sterilization | Fermented fish | Advantages: high-temperature instantaneous sterilization can kill all live bacteria in a short time Disadvantages: changes in flavor and taste, some stubborn bacteria are sublethal | ||
| Bacteriophage antibacterial method | Direct interaction with the host or indirect interaction with released particles | Cheese, fermented soy products | Advantages: no energy consumption, no additives, can improve flavor and quality. Disadvantages: can only inhibit or eliminate specific harmful bacteria |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.; Li, L.; Xiang, P.; Zhang, T.; Peng, L.; Zou, L.; Li, Q. Phages in Fermented Foods: Interactions and Applications. Fermentation 2023, 9, 201. https://doi.org/10.3390/fermentation9030201
Wu Q, Li L, Xiang P, Zhang T, Peng L, Zou L, Li Q. Phages in Fermented Foods: Interactions and Applications. Fermentation. 2023; 9(3):201. https://doi.org/10.3390/fermentation9030201
Chicago/Turabian StyleWu, Qian, Luming Li, Peng Xiang, Ting Zhang, Lianxin Peng, Liang Zou, and Qiang Li. 2023. "Phages in Fermented Foods: Interactions and Applications" Fermentation 9, no. 3: 201. https://doi.org/10.3390/fermentation9030201
APA StyleWu, Q., Li, L., Xiang, P., Zhang, T., Peng, L., Zou, L., & Li, Q. (2023). Phages in Fermented Foods: Interactions and Applications. Fermentation, 9(3), 201. https://doi.org/10.3390/fermentation9030201