
Glycoside Hydrolase Family 48 Cellulase: A Key Player in Cellulolytic Bacteria for Lignocellulose Biorefinery


Abstract
:1. Introduction
2. Key Role of GH48 Cellulases in Cellulose Degradation
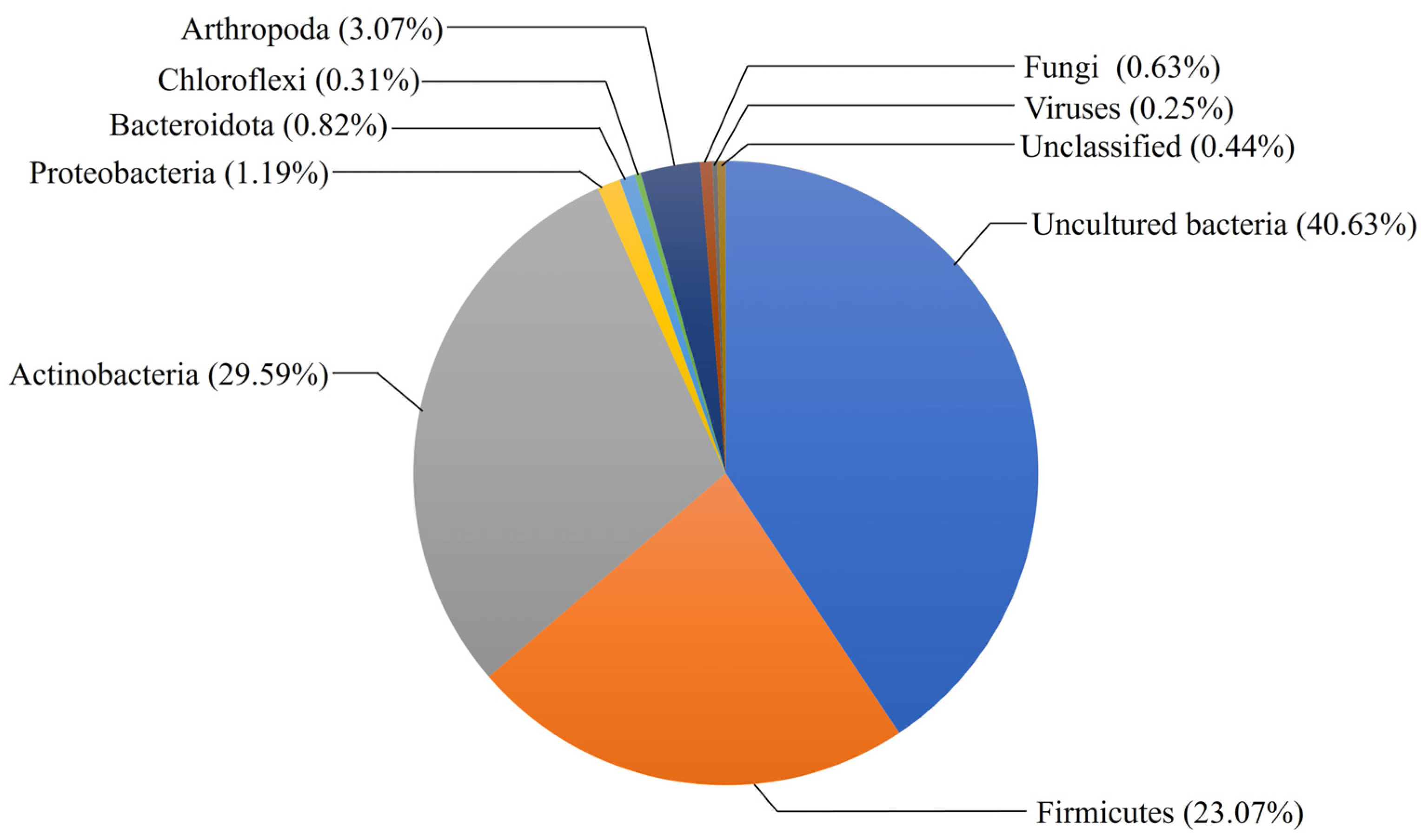
3. Diversity and Distribution of GH48 Family
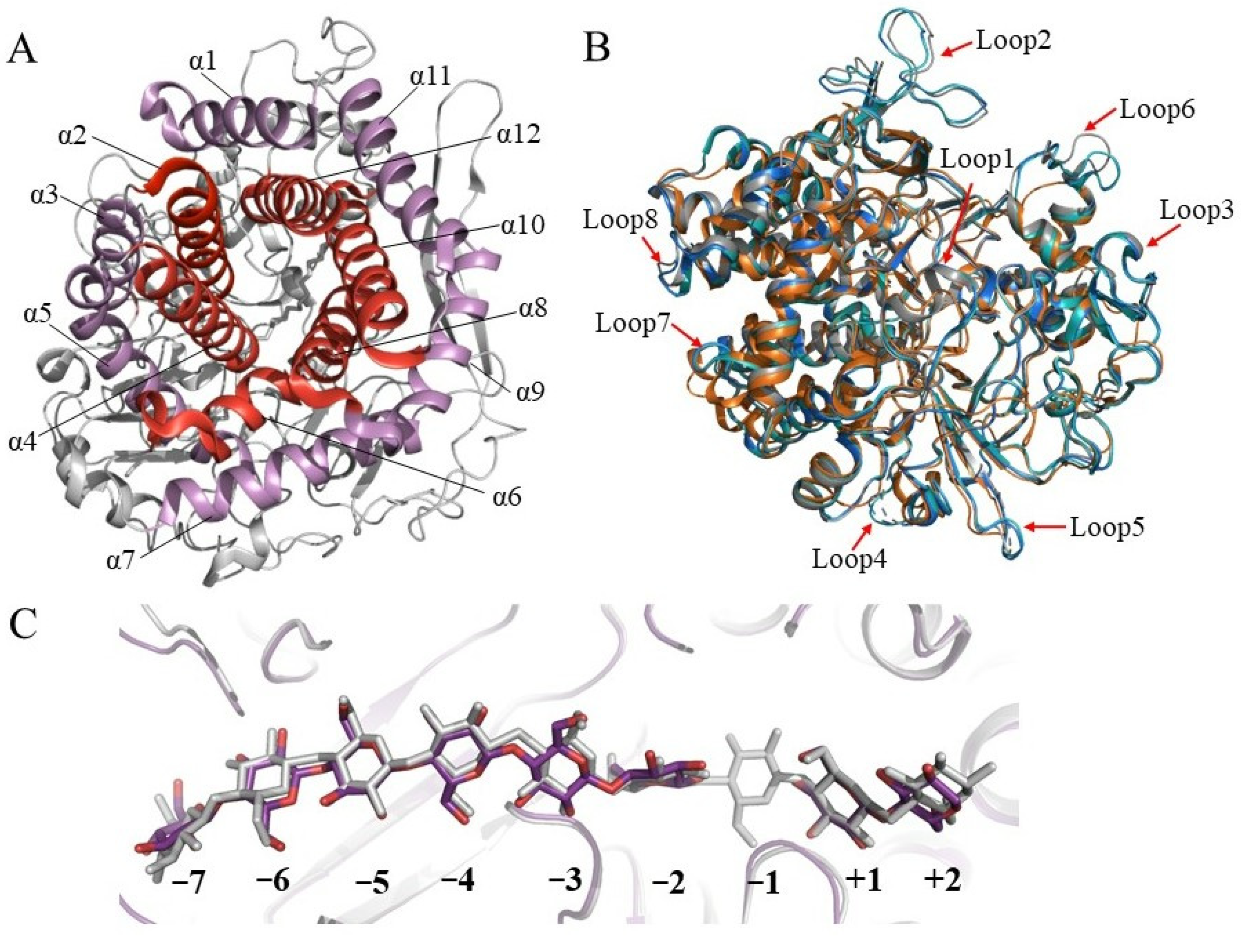
4. Structures of GH48 Cellulases
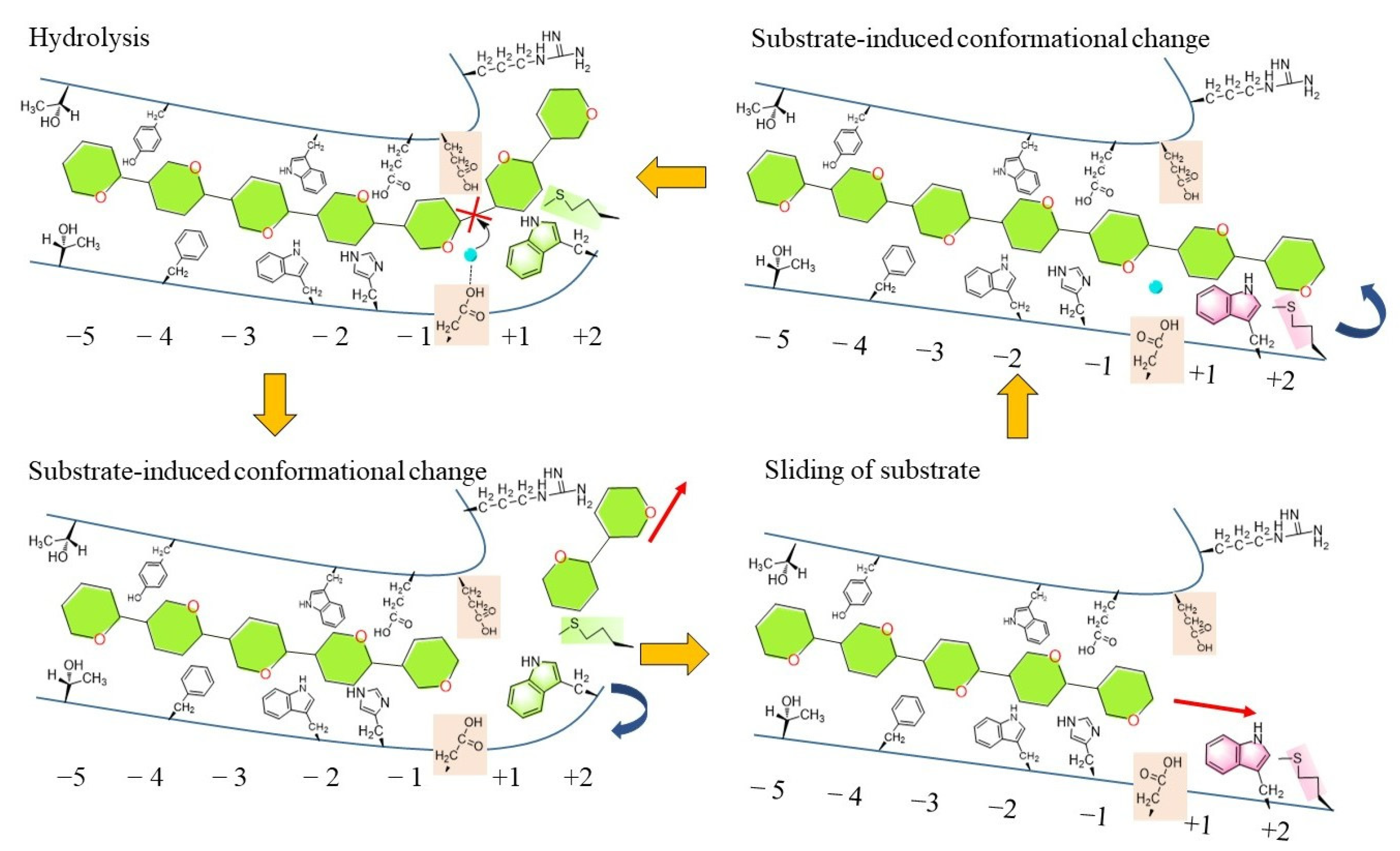
5. Catalytic Mechanism
6. Strategies and Progress of Engineering GH48 Cellulases
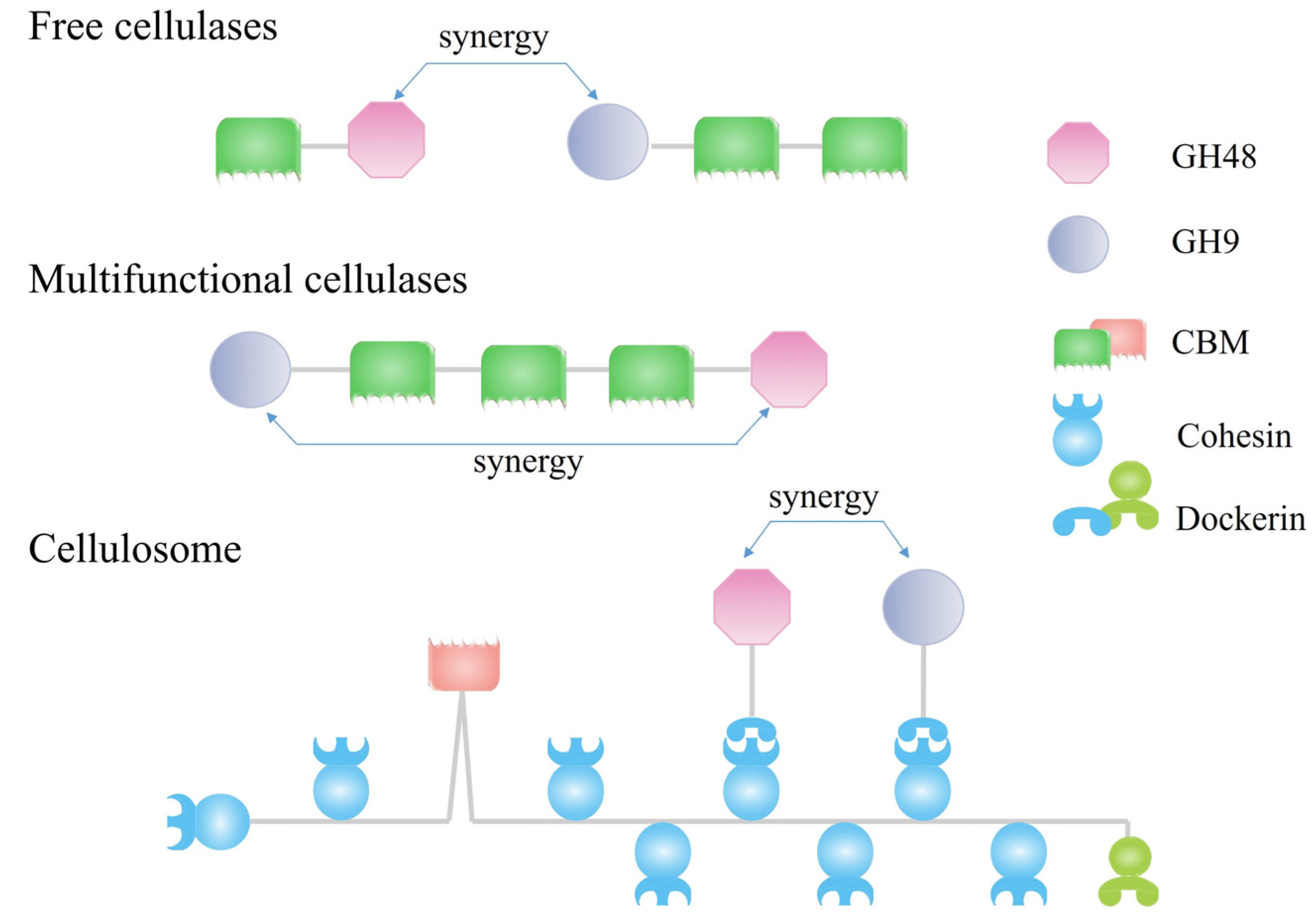
7. Synergistic Effects of GH48 Cellulases with Endocellulases
8. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McKendry, P. Energy production from biomass (part 1): Overview of biomass. Bioresour. Technol. 2002, 83, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Ogonda, L.A.; Saumonneau, A.; Dion, M.; Muge, E.K.; Wamalwa, B.M.; Mulaa, F.J.; Tellier, C. Characterization and engineering of two new GH9 and GH48 cellulases from a Bacillus pumilus isolated from Lake Bogoria. Biotechnol. Lett. 2021, 43, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Himmel, M.E.; Bayer, E.A. Lignocellulose conversion to biofuels: Current challenges, global perspectives. Curr. Opin. Biotechnol. 2009, 20, 316–317. [Google Scholar] [CrossRef] [PubMed]
- Cragg, S.M.; Beckham, G.T.; Bruce, N.C.; Bugg, T.D.H.; Distel, D.L.; Dupree, P.; Etxabe, A.G.; Goodell, B.S.; Jellison, J.; McGeehan, J.E.; et al. Lignocellulose degradation mechanisms across the Tree of Life. Curr. Opin. Chem. Biol. 2015, 29, 108–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taha, M.; Foda, M.; Shahsavari, E.; Aburto-Medina, A.; Adetutu, E.; Ball, A. Commercial feasibility of lignocellulose biodegradation: Possibilities and challenges. Curr. Opin. Biotechnol. 2016, 38, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Alessi, A.M.; Bird, S.M.; Oates, N.C.; Li, Y.; Dowle, A.A.; Novotny, E.H.; deAzevedo, E.R.; Bennett, J.P.; Polikarpov, I.; Young, J.P.W.; et al. Defining functional diversity for lignocellulose degradation in a microbial community using multi-omics studies. Biotechnol. Biofuels 2018, 11, 166. [Google Scholar] [CrossRef]
- Artzi, L.; Bayer, E.A.; Moraïs, S. Cellulosomes: Bacterial nanomachines for dismantling plant polysaccharides. Nat. Rev. Microbiol. 2017, 15, 83–95. [Google Scholar] [CrossRef]
- Payne, C.M.; Knott, B.C.; Mayes, H.B.; Hansson, H.; Himmel, M.E.; Sandgren, M.; Ståhlberg, J.; Beckham, G.T. Fungal cellulases. Chem. Rev. 2015, 115, 1308–1448. [Google Scholar] [CrossRef] [Green Version]
- Østby, H.; Hansen, L.D.; Horn, S.J.; Eijsink, V.G.H.; Várnai, A. Enzymatic processing of lignocellulosic biomass: Principles, recent advances and perspectives. J. Ind. Microbiol. Biotechnol. 2020, 47, 623–657. [Google Scholar] [CrossRef]
- Shallom, D.; Shoham, Y. Microbial hemicellulases. Curr. Opin. Microbiol. 2003, 6, 219–228. [Google Scholar] [CrossRef]
- Kamimura, N.; Sakamoto, S.; Mitsuda, N.; Masai, E.; Kajita, S. Advances in microbial lignin degradation and its applications. Curr. Opin. Biotechnol. 2019, 56, 179–186. [Google Scholar] [CrossRef]
- Liu, Y.-J.; Li, B.; Feng, Y.; Cui, Q. Consolidated bio-saccharification: Leading lignocellulose bioconversion into the real world. Biotechnol. Adv. 2020, 40, 107535. [Google Scholar] [CrossRef]
- Mankar, A.R.; Pandey, A.; Modak, A.; Pant, K.K. Pretreatment of lignocellulosic biomass: A review on recent advances. Bioresour. Technol. 2021, 334, 125235. [Google Scholar] [CrossRef] [PubMed]
- Hagen, L.H.; Brooke, C.G.; Shaw, C.A.; Norbeck, A.D.; Piao, H.; Arntzen, M.Ø.; Olson, H.M.; Copeland, A.; Isern, N.; Shukla, A.; et al. Proteome specialization of anaerobic fungi during ruminal degradation of recalcitrant plant fiber. ISME J. 2021, 15, 421–434. [Google Scholar] [CrossRef]
- Conway, J.M.; Crosby, J.R.; Hren, A.P.; Southerland, R.T.; Lee, L.L.; Lunin, V.V.; Alahuhta, P.; Himmel, M.E.; Bomble, Y.J.; Adams, M.W.W.; et al. Novel multidomain, multifunctional glycoside hydrolases from highly lignocellulolytic Caldicellulosiruptor species. AIChE J. 2018, 64, 4218–4228. [Google Scholar] [CrossRef]
- Li, J.; Su, X.; Tian, Y.; Dong, Z.; Hu, S.; Huang, L.; Dai, X. Gene diversity of the bacterial 48 family glycoside hydrolase (GH48) in rumen environment. Acta Microbiol. Sin. 2014, 54, 53–61. [Google Scholar]
- Chen, M.; Bu, L.; Alahuhta, M.; Brunecky, R.; Xu, Q.; Lunin, V.V.; Brady, J.W.; Crowley, M.F.; Himmel, M.E.; Bomble, Y.J. Strategies to reduce end-product inhibition in family 48 glycoside hydrolases. Proteins Struct. Funct. Bioinform. 2016, 84, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Vazana, Y.; Moraïs, S.; Barak, Y.; Lamed, R.; Bayer, E.A. Interplay between Clostridium thermocellum family 48 and family 9 cellulases in cellulosomal versus noncellulosomal states. Appl. Environ. Microbiol. 2010, 76, 3236–3243. [Google Scholar] [CrossRef] [Green Version]
- Bayer, E.A.; Lamed, R.; White, B.A.; Flint, H.J. From cellulosomes to cellulosomics. Chem. Rec. 2008, 8, 364–377. [Google Scholar] [CrossRef]
- Kim, S.-K.; Chung, D.; Himmel, M.E.; Bomble, Y.J.; Westpheling, J. Engineering the N-terminal end of CelA results in improved performance and growth of Caldicellulosiruptor bescii on crystalline cellulose. Biotechnol. Bioeng. 2017, 114, 945–950. [Google Scholar] [CrossRef]
- Yi, Z.; Su, X.; Revindran, V.; Mackie, R.I.; Cann, I. Molecular and biochemical analyses of CbCel9A/Cel48A, a highly secreted multi-modular cellulase by Caldicellulosiruptor bescii during growth on crystalline cellulose. PLoS ONE 2013, 8, e84172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rettenmaier, R.; Lo, Y.K.; Schmidt, L.; Munk, B.; Lagkouvardos, I.; Neuhaus, K.; Schwarz, W.; Liebl, W.; Zverlov, V. A novel primer mixture for GH48 genes: Quantification and identification of truly cellulolytic bacteria in biogas fermenters. Microorganisms 2020, 8, 1297. [Google Scholar] [CrossRef] [PubMed]
- Drula, E.; Garron, M.-L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2022, 50, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Brás, J.L.A.; Cartmell, A.; Carvalho, A.L.M.; Verzé, G.; Bayer, E.A.; Vazana, Y.; Correia, M.A.S.; Prates, J.A.M.; Ratnaparkhe, S.; Boraston, A.B.; et al. Structural insights into a unique cellulase fold and mechanism of cellulose hydrolysis. Proc. Natl. Acad. Sci. USA 2011, 108, 5237–5242. [Google Scholar] [CrossRef] [Green Version]
- Sukharnikov, L.O.; Alahuhta, M.; Brunecky, R.; Upadhyay, A.; Himmel, M.E.; Lunin, V.V.; Zhulin, I.B. Sequence, structure, and evolution of cellulases in glycoside hydrolase family 48. J. Biol. Chem. 2012, 287, 41068–41077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, L.C.; Amiraslanov, I.; Chen, H.R.; Chen, Y.W.; Lee, H.L.; Liang, P.H.; Liaw, Y.C. Structures of exoglucanase from Clostridium cellulovorans: Cellotetraose binding and cleavage. Acta Crystallogr. F Struct. Biol. Commun. 2015, 71, 1264–1272. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, J.-L.; Sun, L.; Niu, X.-D.; Wang, S.; Shan, Y.-M. Molecular dynamics simulation of the processive endocellulase Cel48F from Clostridium cellulolyticum: A novel “water-control mechanism” in enzymatic hydrolysis of cellulose. J. Mol. Recognit. 2014, 27, 438–447. [Google Scholar] [CrossRef]
- Araújo, E.A.; Dias, A.H.S.; Kadowaki, M.A.S.; Piyadov, V.; Pellegrini, V.O.A.; Urio, M.B.; Ramos, L.P.; Skaf, M.S.; Polikarpov, I. Impact of cellulose properties on enzymatic degradation by bacterial GH48 enzymes: Structural and mechanistic insights from processive Bacillus licheniformis Cel48B cellulase. Carbohydr. Polym. 2021, 264, 118059. [Google Scholar] [CrossRef]
- Penneru, S.K.; Saharay, M.; Krishnan, M. CelS-catalyzed processive cellulose degradation and cellobiose extraction for the production of bioethanol. J. Chem. Inf. Model. 2022, 62, 6628–6638. [Google Scholar] [CrossRef]
- Himmel, M.E.; Ding, S.-Y.; Johnson, D.K.; Adney, W.S.; Nimlos, M.R.; Brady, J.W.; Foust, T.D. Biomass recalcitrance: Engineering plants and enzymes for biofuels production. Science 2007, 315, 804–807. [Google Scholar] [CrossRef] [Green Version]
- Usmani, Z.; Sharma, M.; Awasthi, A.K.; Lukk, T.; Tuohy, M.G.; Gong, L.; Nguyen-Tri, P.; Goddard, A.D.; Bill, R.M.; Nayak, S.C.; et al. Lignocellulosic biorefineries: The current state of challenges and strategies for efficient commercialization. Renew. Sustain. Energy Rev. 2021, 148, 111258. [Google Scholar] [CrossRef]
- Liu, Y.; Cruz-Morales, P.; Zargar, A.; Belcher, M.S.; Pang, B.; Englund, E.; Dan, Q.; Yin, K.; Keasling, J.D. Biofuels for a sustainable future. Cell 2021, 184, 1636–1647. [Google Scholar] [CrossRef] [PubMed]
- Devillard, E.; Goodheart, D.B.; Karnati, S.K.R.; Bayer, E.A.; Lamed, R.; Miron, J.; Nelson, K.E.; Morrison, M. Ruminococcus albus 8 mutants defective in cellulose degradation are deficient in two processive endocellulases, Cel48A and Cel9B, both of which possess a novel modular architecture. J. Bacteriol. 2004, 186, 136–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izquierdo, J.A.; Sizova, M.V.; Lynd, L.R. Diversity of bacteria and glycosyl hydrolase family 48 genes in cellulolytic consortia enriched from thermophilic biocompost. Appl. Environ. Microbiol. 2010, 76, 3545–3553. [Google Scholar] [CrossRef] [Green Version]
- Olson, D.G.; Tripathi, S.A.; Giannone, R.J.; Lo, J.; Caiazza, N.C.; Hogsett, D.A.; Hettich, R.L.; Guss, A.M.; Dubrovsky, G.; Lynd, L.R. Deletion of the Cel48S cellulase from Clostridium thermocellum. Proc. Natl. Acad. Sci. USA 2010, 107, 17727–17732. [Google Scholar] [CrossRef] [Green Version]
- Young, J.; Chung, D.; Bomble, Y.J.; Himmel, M.E.; Westpheling, J. Deletion of Caldicellulosiruptor bescii CelA reveals its crucial role in the deconstruction of lignocellulosic biomass. Biotechnol. Biofuels 2014, 7, 142. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Tian, Y.; Li, J.; Luo, Y.; Liu, D.; Zheng, H.; Wang, J.; Dong, Z.; Hu, S.; Huang, L. Metatranscriptomic analyses of plant cell wall polysaccharide degradation by microorganisms in the cow rumen. Appl. Environ. Microbiol. 2015, 81, 1375–1386. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.; Hao, Z.; Wang, K.; Tu, T.; Huang, H.; Wang, Y.; Bai, Y.G.; Wang, Y.; Luo, H.; Yao, B.; et al. The GH10 and GH48 dual-functional catalytic domains from a multimodular glycoside hydrolase synergize in hydrolyzing both cellulose and xylan. Biotechnol. Biofuels 2019, 12, 279. [Google Scholar] [CrossRef]
- Lee, L.L.; Blumer-Schuette, S.E.; Izquierdo, J.A.; Zurawski, J.V.; Loder, A.J.; Conway, J.M.; Elkins, J.G.; Podar, M.; Clum, A.; Jones, P.C.; et al. Genus-wide assessment of lignocellulose utilization in the extremely thermophilic genus Caldicellulosiruptor by genomic, pangenomic, and metagenomic analyses. Appl. Environ. Microbiol. 2018, 84, e02694-17. [Google Scholar] [CrossRef] [Green Version]
- Pereyra, L.P.; Hiibel, S.R.; Prieto Riquelme, M.V.; Reardon, K.F.; Pruden, A. Detection and quantification of functional genes of cellulose- degrading, fermentative, and sulfate-reducing bacteria and methanogenic archaea. Appl. Environ. Microbiol. 2010, 76, 2192–2202. [Google Scholar] [CrossRef] [Green Version]
- Brunecky, R.; Alahuhta, M.; Xu, Q.; Donohoe, B.S.; Crowley, M.F.; Kataeva, I.A.; Yang, S.-J.; Resch, M.G.; Adams, M.W.W.; Lunin, V.V.; et al. Revealing nature’s cellulase diversity: The digestion mechanism of Caldicellulosiruptor bescii CelA. Science 2013, 342, 1513–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.K.; Kruus, K.; Wu, J.H.D. Cloning and DNA sequence of the gene coding for Clostridium thermocellum cellulase Ss (CelS), a major cellulosome component. J. Bacteriol. 1993, 175, 1293–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.K.; Kruus, K.; Wu, J.H.D. Cloning and expression of the Clostridium thermocellum celS gene in Escherichia coli. Appl. Microbiol. Biotechnol. 1994, 42, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Bagnara-Tardif, C.; Gaudin, C.; Belaich, A.; Hoest, P.; Citard, T.; Belaich, J.-P. Sequence analysis of a gene cluster encoding cellulases from Clostridium cellulolyticum. Gene 1992, 119, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Conway, J.M.; McKinley, B.S.; Seals, N.L.; Hernandez, D.; Khatibi, P.A.; Poudel, S.; Giannone, R.J.; Hettich, R.L.; Williams-Rhaesa, A.M.; Lipscomb, G.L.; et al. Functional analysis of the glucan degradation locus in Caldicellulosiruptor bescii reveals essential roles of component glycoside hydrolases in plant biomass deconstruction. Appl. Environ. Microbiol. 2017, 83, e01828-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zverlov, V.; Mahr, S.; Riedel, K.; Bronnenmeier, K. Properties and gene structure of a bifunctional cellulolytic enzyme (CelA) from the extreme thermophile ‘Anaerocellum thermophilum’ with separate glycosyl hydrolase family 9 and 48 catalytic domains. Microbiology 1998, 144, 457–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostylev, M.; Wilson, D. A distinct model of synergism between a processive endocellulase (TfCel9A) and an exocellulase (TfCel48A) from Thermobifida fusca. Appl. Environ. Microbiol. 2014, 80, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Eyun, S.-I.; Wang, H.; Pauchet, Y.; Ffrench-Constant, R.H.; Benson, A.K.; Valencia-Jiménez, A.; Moriyama, E.N.; Siegfried, B.D. Molecular evolution of glycoside hydrolase genes in the Western corn rootworm (Diabrotica virgifera virgifera). PLoS ONE 2014, 9, e94052. [Google Scholar] [CrossRef]
- Comtet-Marre, S.; Parisot, N.; Lepercq, P.; Chaucheyras-Durand, F.; Mosoni, P.; Peyretaillade, E.; Bayat, A.R.; Shingfield, K.J.; Peyret, P.; Forano, E. Metatranscriptomics reveals the active bacterial and eukaryotic fibrolytic communities in the rumen of dairy cow fed a mixed diet. Front. Microbiol. 2017, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Parsiegla, G.; Juy, M.; Reverbel-Leroy, C.; Tardif, C.; Belaïch, J.P.; Driguez, H.; Haser, R. The crystal structure of the processive endocellulase CelF of Clostridium cellulolyticum in complex with a thiooligosaccharide inhibitor at 2.0 Å resolution. EMBO J. 1998, 17, 5551–5562. [Google Scholar] [CrossRef] [Green Version]
- Parsiegla, G.; Reverbel-Leroy, C.; Tardif, C.; Belaich, J.P.; Driguez, H.; Haser, R. Crystal structures of the cellulase Cel48F in complex with inhibitors and substrates give insights into its processive action. Biochemistry 2000, 39, 11238–11246. [Google Scholar] [CrossRef] [PubMed]
- Parsiegla, G.; Reverbel, C.; Tardif, C.; Driguez, H.; Haser, R. Structures of mutants of cellulase Cel48F of Clostridium cellulolyticum in complex with long hemithiocellooligosaccharides give rise to a new view of the substrate pathway during processive action. J. Mol. Biol. 2008, 375, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-J.; Liu, S.; Dong, S.; Li, R.; Feng, Y.; Cui, Q. Determination of the native features of the exoglucanase Cel48S from Clostridium thermocellum. Biotechnol. Biofuels 2018, 11, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimarães, B.G.; Souchon, H.; Lytle, B.L.; David Wu, J.H.; Alzari, P.M. The crystal structure and catalytic mechanism of cellobiohydrolase CelS, the major enzymatic component of the Clostridium thermocellum cellulosome. J. Mol. Biol. 2002, 320, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Berger, E.; Zhang, D.; Zverlov, V.V.; Schwarz, W.H. Two noncellulosomal cellulases of Clostridium thermocellum, Cel9I and Cel48Y, hydrolyse crystalline cellulose synergistically. FEMS Microbiol. Lett. 2007, 268, 194–201. [Google Scholar] [CrossRef] [Green Version]
- Kruus, K.; Wang, W.K.; Ching, J.; Wu, J.H.D. Exoglucanase activities of the recombinant Clostridium thermocellum CelS, a major cellulosome component. J. Bacteriol. 1995, 177, 1641–1644. [Google Scholar] [CrossRef] [Green Version]
- Bronnenmeier, K.; Kundt, K.; Riedel, K.; Schwarz, W.H.; Staudenbauer, W.L. Structure of the Clostridium stercorarium gene celY encoding the exo-1,4-β-glucanase Avicelase II. Microbiology 1997, 143, 891–898. [Google Scholar] [CrossRef] [Green Version]
- Artzi, L.; Morag, E.; Shamshoum, M.; Bayer, E.A. Cellulosomal expansin: Functionality and incorporation into the complex. Biotechnol. Biofuels 2016, 9, 61. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-Z.; Zhang, Z.; Zhu, Z.; Sathitsuksanoh, N.; Yang, Y.; Zhang, Y.-H.P. The noncellulosomal family 48 cellobiohydrolase from Clostridium phytofermentans ISDg: Heterologous expression, characterization, and processivity. Appl. Microbiol. Biotechnol. 2010, 86, 525–533. [Google Scholar] [CrossRef]
- Brunecky, R.; Alahuhta, M.; Sammond, D.W.; Xu, Q.; Chen, M.; Wilson, D.B.; Brady, J.W.; Himmel, M.E.; Bomble, Y.J.; Lunin, V.V. Natural diversity of glycoside hydrolase family 48 exoglucanases: Insights from structure. Biotechnol. Biofuels 2017, 10, 274. [Google Scholar] [CrossRef] [Green Version]
- Kostylev, M.; Alahuhta, M.; Chen, M.; Brunecky, R.; Himmel, M.E.; Lunin, V.V.; Brady, J.; Wilson, D.B. Cel48A from Thermobifida fusca: Structure and site directed mutagenesis of key residues. Biotechnol. Bioeng. 2014, 111, 664–673. [Google Scholar] [CrossRef]
- Shen, H.; Gilkes, N.R.; Kilburn, D.G.; Miller Jr, R.C.; Warren, R.A. Cellobiohydrolase B, a second exo-cellobiohydrolase from the cellulolytic bacterium Cellulomonas fimi. Biochem. J. 1995, 311, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Li, J.; Hu, F.Z.; Zhang, K.; Luo, Y.; Janto, B.; Boissy, R.; Ehrlich, G.; Dong, X. Cellulosilyticum ruminicola, a newly described rumen bacterium that possesses redundant fibrolytic-protein-encoding genes and degrades lignocellulose with multiple carbohydrate- borne fibrolytic enzymes. Appl. Environ. Microbiol. 2010, 76, 3818–3824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez-Ramírez, N.; Romero-García, E.R.; Calderón, V.C.; Avitia, C.I.; Téllez-Valencia, A.; Pedraza-Reyes, M. Expression, characterization and synergistic interactions of Myxobacter Sp. AL-1 Cel9 and Cel48 glycosyl hydrolases. Int. J. Mol. Sci. 2008, 9, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, M.M.; Pastor, F.I.J.; Diaz, P. Exo-mode of action of cellobiohydrolase Cel48C from Paenibacillus sp. BP-23. A unique type of cellulase among Bacillales. Eur. J. Biochem. 2003, 270, 2913–2919. [Google Scholar] [CrossRef] [PubMed]
- Qian, M.; Guan, S.; Shan, Y.; Zhang, H.; Wang, S. Structural and molecular basis of cellulase Cel48F by computational modeling: Insight into catalytic and product release mechanism. J. Struct. Biol. 2016, 194, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Saharay, M.; Guo, H.; Smith, J.C. Catalytic mechanism of cellulose degradation by a cellobiohydrolase, CelS. PLoS ONE 2010, 5, e12947. [Google Scholar] [CrossRef]
- Kurasin, M.; Väljamäe, P. Processivity of cellobiohydrolases is limited by the substrate. J. Biol. Chem. 2011, 286, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, V.O.A.; Bernardes, A.; Rezende, C.A.; Polikarpov, I. Cellulose fiber size defines efficiency of enzymatic hydrolysis and impacts degree of synergy between endo- and exoglucanases. Cellulose 2018, 25, 1865–1881. [Google Scholar] [CrossRef]
- Morag, E.; Halevy, I.; Bayer, E.A.; Lamed, R. Isolation and properties of a major cellobiohydrolase from the cellulosome of Clostridium thermocellum. J. Bacteriol. 1991, 173, 4155–4162. [Google Scholar] [CrossRef] [Green Version]
- Irwin, D.C.; Zhang, S.; Wilson, D.B. Cloning, expression and characterization of a family 48 exocellulase, Cel48A, from Thermobifida fusca. Eur. J. Biochem. 2000, 267, 4988–4997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, L.; Crowley, M.F.; Himmel, M.E.; Beckham, G.T. Computational investigation of the pH dependence of loop flexibility and catalytic function in glycoside hydrolases. J. Biol. Chem. 2013, 288, 12175–12186. [Google Scholar] [CrossRef] [Green Version]
- Bu, L.; Nimlos, M.R.; Shirts, M.R.; Ståhlberg, J.; Himmel, M.E.; Crowley, M.F.; Beckham, G.T. Product binding varies dramatically between processive and nonprocessive cellulase enzymes. J. Biol. Chem. 2012, 287, 24807–24813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.-J.; Kang, T.-G.; Kim, Y.-W.; Lee, H.-S.; Kim, S.-K. Functional expression and extracellular secretion of Clostridium thermocellum Cel48S cellulase in Escherichia coli via the signal recognition particle-dependent translocation pathway. Enzyme Microb. Technol. 2021, 151, 109918. [Google Scholar] [CrossRef]
- Smith, M.A.; Rentmeister, A.; Snow, C.D.; Wu, T.; Farrow, M.F.; Mingardon, F.; Arnold, F.H. A diverse set of family 48 bacterial glycoside hydrolase cellulases created by structure-guided recombination. FEBS J. 2012, 279, 4453–4465. [Google Scholar] [CrossRef] [PubMed]
- Reese, E.T.; Siu, R.G.H.; Levinson, H.S. The biological degradation of soluble cellulose derivatives and its relationship to the mechanism of cellulose hydrolysis. J. Bacteriol. 1950, 59, 485–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, T.M.; McCrae, S.I. Synergism Between Enzymes Involved in the Solubilization of Native Cellulose. In Hydrolysis of Cellulose: Mechanisms of Enzymatic and Acid Catalysis; Brown, R.D., Jurasek, L., Eds.; American Chemical Society: Washington, DC, USA, 1979; pp. 181–209. ISBN 9780841204607. [Google Scholar]
- Kostylev, M.; Wilson, D. Synergistic interactions in cellulose hydrolysis. Biofuels 2012, 3, 61–70. [Google Scholar] [CrossRef]
- Xu, Q.; Ding, S.-Y.; Brunecky, R.; Bomble, Y.J.; Himmel, M.E.; Baker, J.O. Improving activity of minicellulosomes by integration of intra- and intermolecular synergies. Biotechnol. Biofuels 2013, 6, 126. [Google Scholar] [CrossRef] [Green Version]
- Irwin, D.C.; Spezio, M.; Walker, L.P.; Wilson, D.B. Activity studies of eight purified cellulases: Specificity, synergism, and binding domain effects. Biotechnol. Bioeng. 1993, 42, 1002–1013. [Google Scholar] [CrossRef]
- Lochner, A.; Giannone, R.J.; Rodriguez, M.; Shah, M.B.; Mielenz, J.R.; Keller, M.; Antranikian, G.; Graham, D.E.; Hettich, R.L. Use of label-free quantitative proteomics to distinguish the secreted cellulolytic systems of Caldicellulosiruptor bescii and Caldicellulosiruptor obsidiansis. Appl. Environ. Microbiol. 2011, 77, 4042–4054. [Google Scholar] [CrossRef] [Green Version]
- Bing, W.; Wang, H.; Zheng, B.; Zhang, F.; Zhu, G.; Feng, Y.; Zhang, Z. Caldicellulosiruptor changbaiensis sp. nov., a cellulolytic and hydrogen-producing bacterium from a hot spring. Int. J. Syst. Evol. Microbiol. 2015, 65, 293–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miroshnichenko, M.L.; Kublanov, I.V.; Kostrikina, N.A.; Tourova, T.P.; Kolganova, T.V.; Birkeland, N.-K.; Bonch-Osmolovskaya, E.A. Caldicellulosiruptor kronotskyensis sp. nov. and Caldicellulosiruptor hydrothermalis sp. nov., two extremely thermophilic, cellulolytic, anaerobic bacteria from Kamchatka thermal springs. Int. J. Syst. Evol. Microbiol. 2008, 58, 1492–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Liu, Y.; Cui, Q. Research progress in cellulosomes and their applications in synthetic biology. Synth. Biol. J. 2022, 3, 138–154. [Google Scholar] [CrossRef]
- Yao, X.; Chen, C.; Wang, Y.; Dong, S.; Liu, Y.-J.; Li, Y.; Cui, Z.; Gong, W.; Perrett, S.; Yao, L.; et al. Discovery and mechanism of a pH-dependent dual-binding-site switch in the interaction of a pair of protein modules. Sci. Adv. 2020, 6, eabd7182. [Google Scholar] [CrossRef]
- Bule, P.; Pires, V.; Fontes, C.M.; Alves, V.D. Cellulosome assembly: Paradigms are meant to be broken! Curr. Opin. Struct. Biol. 2018, 49, 154–161. [Google Scholar] [CrossRef]
- Yoav, S.; Barak, Y.; Shamshoum, M.; Borovok, I.; Lamed, R.; Dassa, B.; Hadar, Y.; Morag, E.; Bayer, E.A. How does cellulosome composition influence deconstruction of lignocellulosic substrates in Clostridium (Ruminiclostridium) thermocellum DSM 1313? Biotechnol. Biofuels 2017, 10, 222. [Google Scholar] [CrossRef] [Green Version]
- Ravachol, J.; Borne, R.; Tardif, C.; de Philip, P.; Fierobe, H.-P. Characterization of all family-9 glycoside hydrolases synthesized by the cellulosome-producing bacterium Clostridium cellulolyticum. J. Biol. Chem. 2014, 289, 7335–7348. [Google Scholar] [CrossRef] [Green Version]
- Moraïs, S.; Ben David, Y.; Bensoussan, L.; Duncan, S.H.; Koropatkin, N.M.; Martens, E.C.; Flint, H.J.; Bayer, E.A. Enzymatic profiling of cellulosomal enzymes from the human gut bacterium, Ruminococcus champanellensis, reveals a fine-tuned system for cohesin-dockerin recognition. Environ. Microbiol. 2016, 18, 542–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reverbel-Leroy, C.; Belaich, A.; Bernadac, A.; Gaudin, C.; Belaich, J.-P.; Tardif, C. Molecular study and overexpression of the Clostridium cellulolyticum celF cellulase gene in Escherichia coli. Microbiology 1996, 142, 1013–1023. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GH48s | Activities | Origin | PDB Code (Ligand) | Form | Reference |
|---|---|---|---|---|---|
| CelS (Cel48S) | Exocellulase | C. thermocellum DSM1313 | 5YJ6 | cellulosomal | [53] |
| CelS (Cel48A) | Exocellulase | C. thermocellum F7 | 1L1Y(cellobiose), 1L2A (cellobiose, cellohexaose) | cellulosomal | [54] |
| Cel48Y | Exocellulase | C. thermocellum ATCC 27405 | free-cellulase | [55] | |
| Cel48S | Exocellulase | C. thermocellum ATCC 27405 | cellulosomal | [56] | |
| CelY | Exocellulase | Clostridium stercorarium | free-cellulase | [57] | |
| GH48 | Exocellulase | Clostridium clariflavum DSM 19732 | cellulosomal | [58] | |
| CpCel48 | Exocellulase | Clostridium phytofermentans ISDg | free-cellulase | [59] | |
| ExgS | Exocellulase | Clostridium cellulovorans ATCC 35296 | 4XWL (PEG), 4XWM (cellobiose), 4XWN (cellobiose, cellopentaose) | cellulosomal | [26] |
| Cel48F | Processive endocellulase | C. cellulolyticum H10 | 1FCE (inhibitor IG4), 1F9D (cellotetraose), 1FBW (cellohexaose), 1FAE (cellobiose), 1FBO (cellobiitol), 1F9O (inhibitor PIPS-IG3), 1G9G (glucose), 2QNO (thiocellodecaose), 1G9J (hemithiocellooligosaccharide) | cellulosomal | [50,51,52] |
| CbCel48A | Exocellulase | C. bescii DSM 6725 | 4EL8, 4L0G (cellobiose), 4TXT (cellotriose), 4L6X | multi- module | [41] |
| Cdan_2053 | Cellulase | Caldicellulosiruptor danielii | 6D5D (cellobiose) | multi-module | [15] |
| BlCel48B | Processive cellulase | Bacillus licheniformis DSM 13 | 7KW6 (cellobiose, cellotetraose) | free-cellulase | [28] |
| BpCel48 | Cellulase | Bacillus pmilus SAFR-032 | 5BV9 (cellobiose), 5CVY (cellobiose, cellohexaose) | free-cellulase | [17,60] |
| BpGH48 | Cellulase | Bacillus pmilus SH-B9 | 5VMA (cellobio-derived isofagomine) | free-cellulase | To be published |
| TfCel48A | Exocellulase | T. fusca YX | 4JJJ (cellobiose, cellohexaose) | free-cellulase | [61] |
| HcheGH48 | Cellulase | H. chejuensis KCTC 2396 | 4FUS (cellobiose) | free-cellulase | [25] |
| CbhB | Exocellulase | Cellulomonas fimi ATCC 484 | free-cellulase | [62] | |
| Cel48A | Exocellulase | C. ruminicola H1 | free-cellulase | [63] | |
| Cel48 | Exocellulase | Myxobacter sp. AL-1 | free-cellulase | [64] | |
| Cel48C | Exocellulase | Paenibacillus sp. BP-23 | free-cellulase | [65] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
You, C.; Liu, Y.-J.; Cui, Q.; Feng, Y. Glycoside Hydrolase Family 48 Cellulase: A Key Player in Cellulolytic Bacteria for Lignocellulose Biorefinery. Fermentation 2023, 9, 204. https://doi.org/10.3390/fermentation9030204
You C, Liu Y-J, Cui Q, Feng Y. Glycoside Hydrolase Family 48 Cellulase: A Key Player in Cellulolytic Bacteria for Lignocellulose Biorefinery. Fermentation. 2023; 9(3):204. https://doi.org/10.3390/fermentation9030204
Chicago/Turabian StyleYou, Cai, Ya-Jun Liu, Qiu Cui, and Yingang Feng. 2023. "Glycoside Hydrolase Family 48 Cellulase: A Key Player in Cellulolytic Bacteria for Lignocellulose Biorefinery" Fermentation 9, no. 3: 204. https://doi.org/10.3390/fermentation9030204
APA StyleYou, C., Liu, Y. -J., Cui, Q., & Feng, Y. (2023). Glycoside Hydrolase Family 48 Cellulase: A Key Player in Cellulolytic Bacteria for Lignocellulose Biorefinery. Fermentation, 9(3), 204. https://doi.org/10.3390/fermentation9030204