
Impact of Spontaneous Fermentation and Inoculum with Natural Whey Starter on Peptidomic Profile and Biological Activities of Cheese Whey: A Comparative Study

 ,
, 
 and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Fermentation Trials and Chemico-Physical Analysis
2.3. Microbiological Analysis
2.4. Assessment of Protein Hydrolysos during the Fermnetation Trials
2.5. Low-Molecular Weight Peptide Fractions Extraction and Biological Activity Assays
2.6. Analysis of the Peptide Profiles by High-Resolution Mass Spectrometry
2.7. Peptidomics Analysis
2.7.1. Peptides Sequencing
2.7.2. Label-Free MS Peak Quantification
2.7.3. Bioactive Peptides Identification and Relative Quantification
2.8. Absolute Quantification of the Bioactive Peptides IPP and VPP
2.9. Statistical Analysis
3. Results and Discussion
3.1. NWS Microbial Caracterization
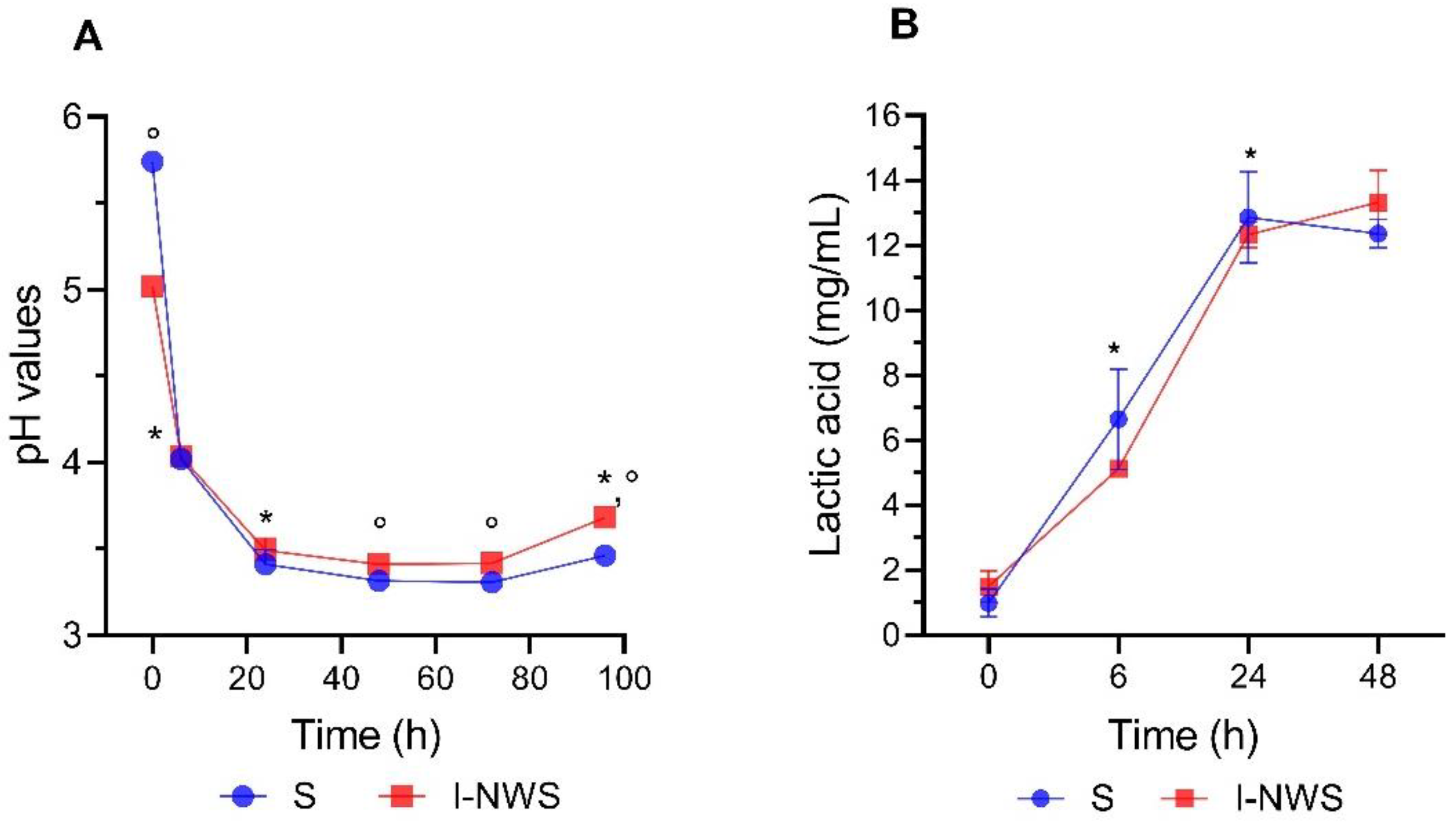
3.2. Whey Fermentation
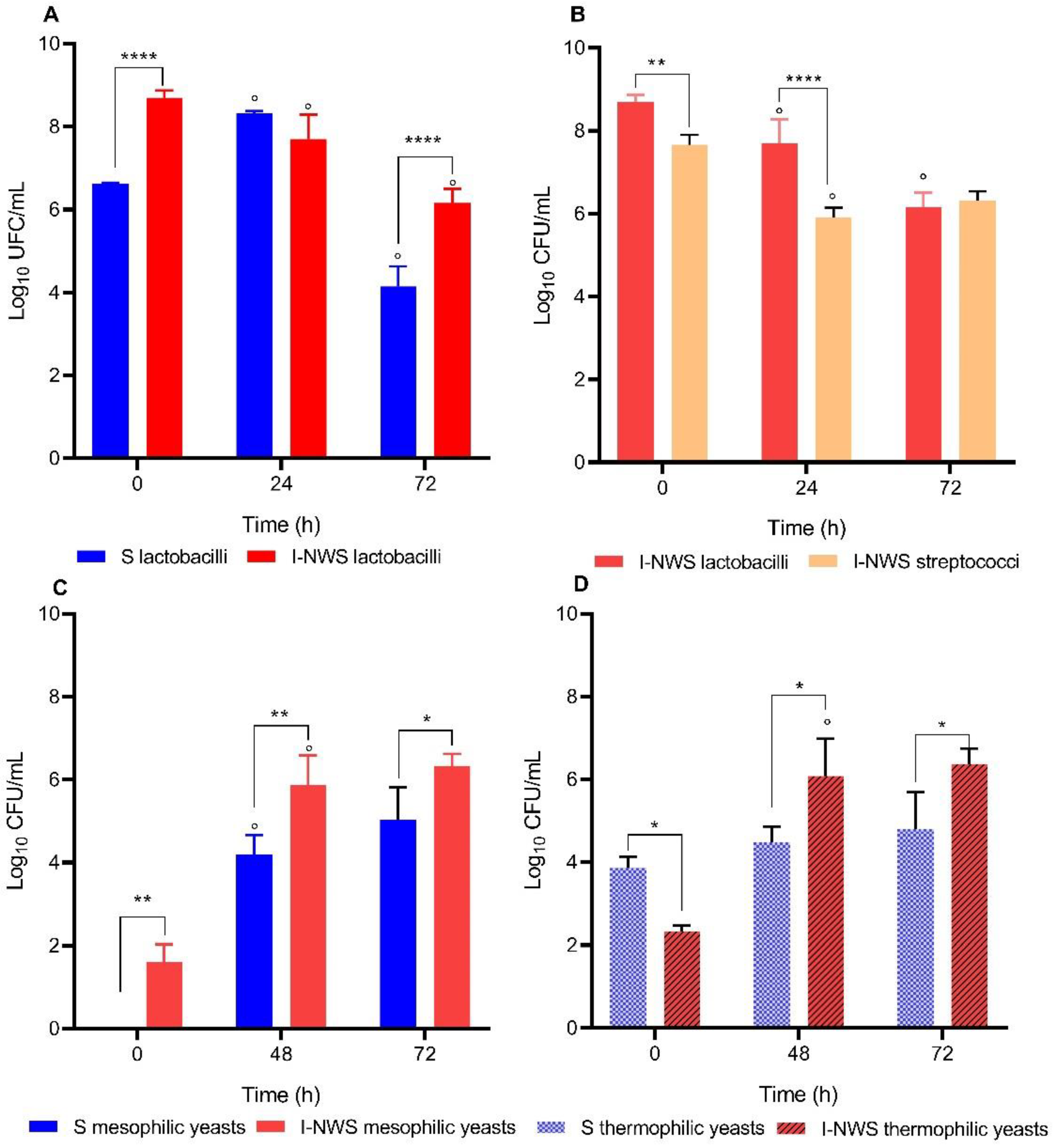
3.3. Enumeration of Lactic Acid Bacteria and Yeasts Populations
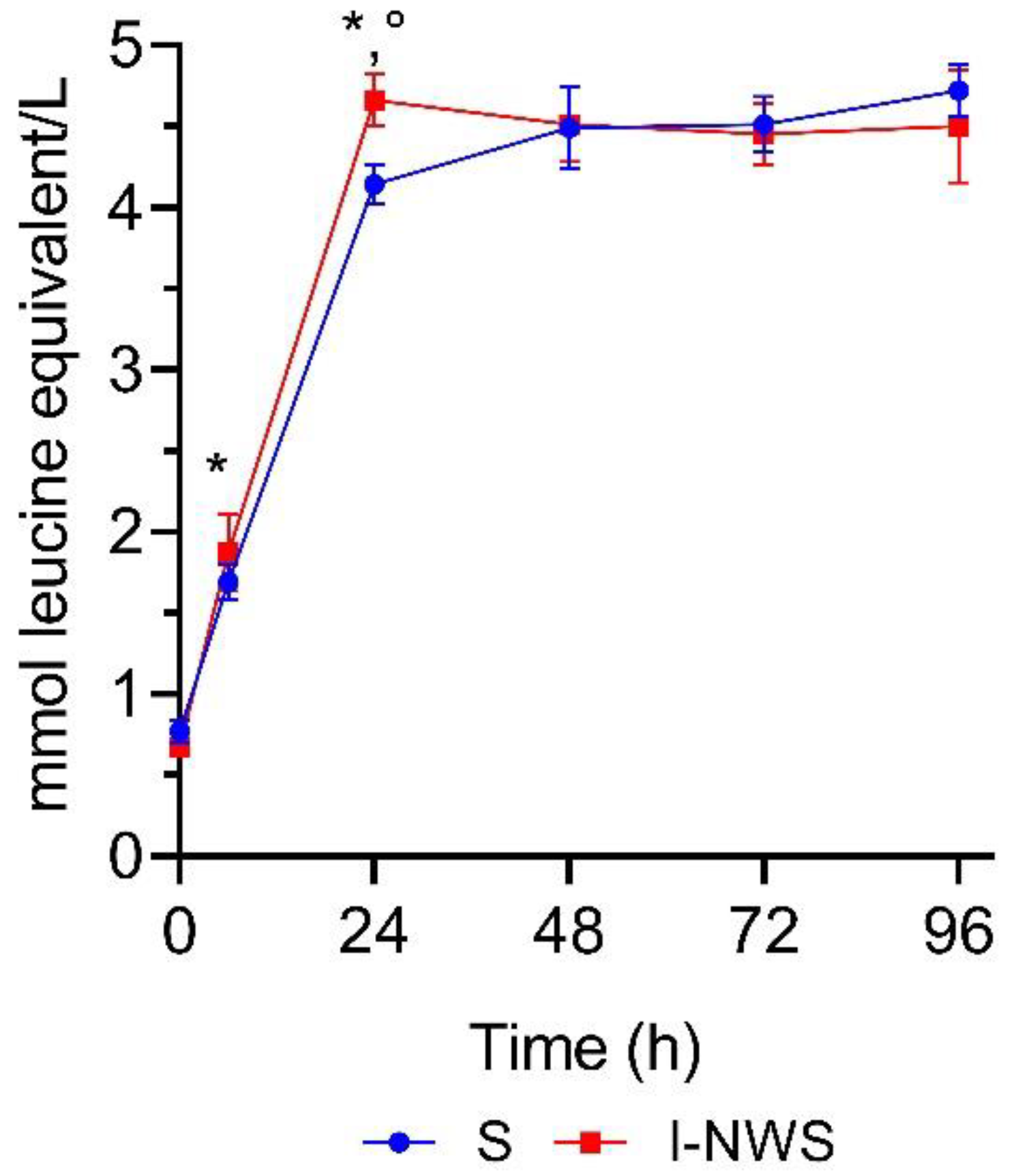
3.4. Assessment of Proteolytic Activity during Whey Fermentation
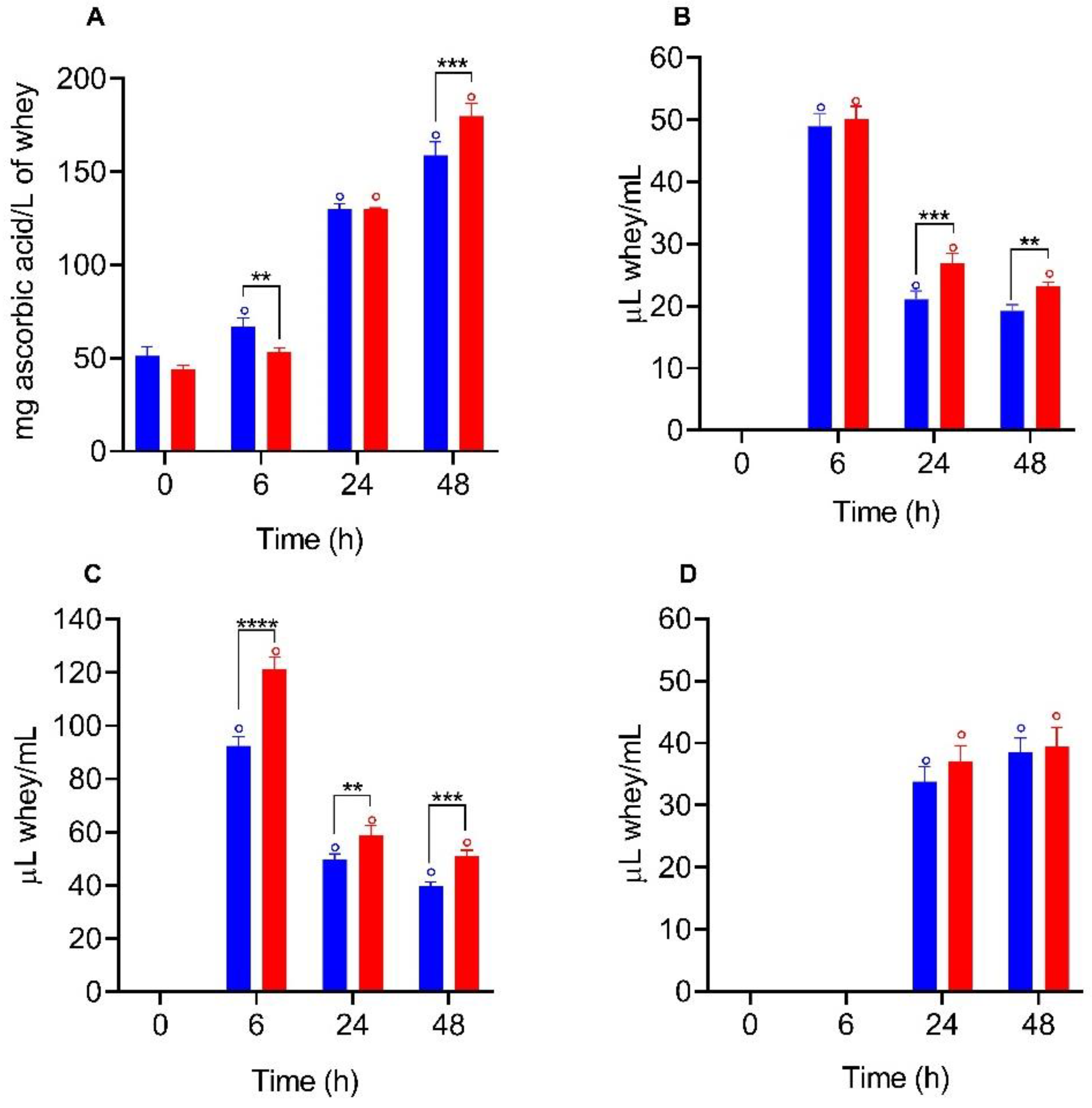
3.5. Biological Activities Determination
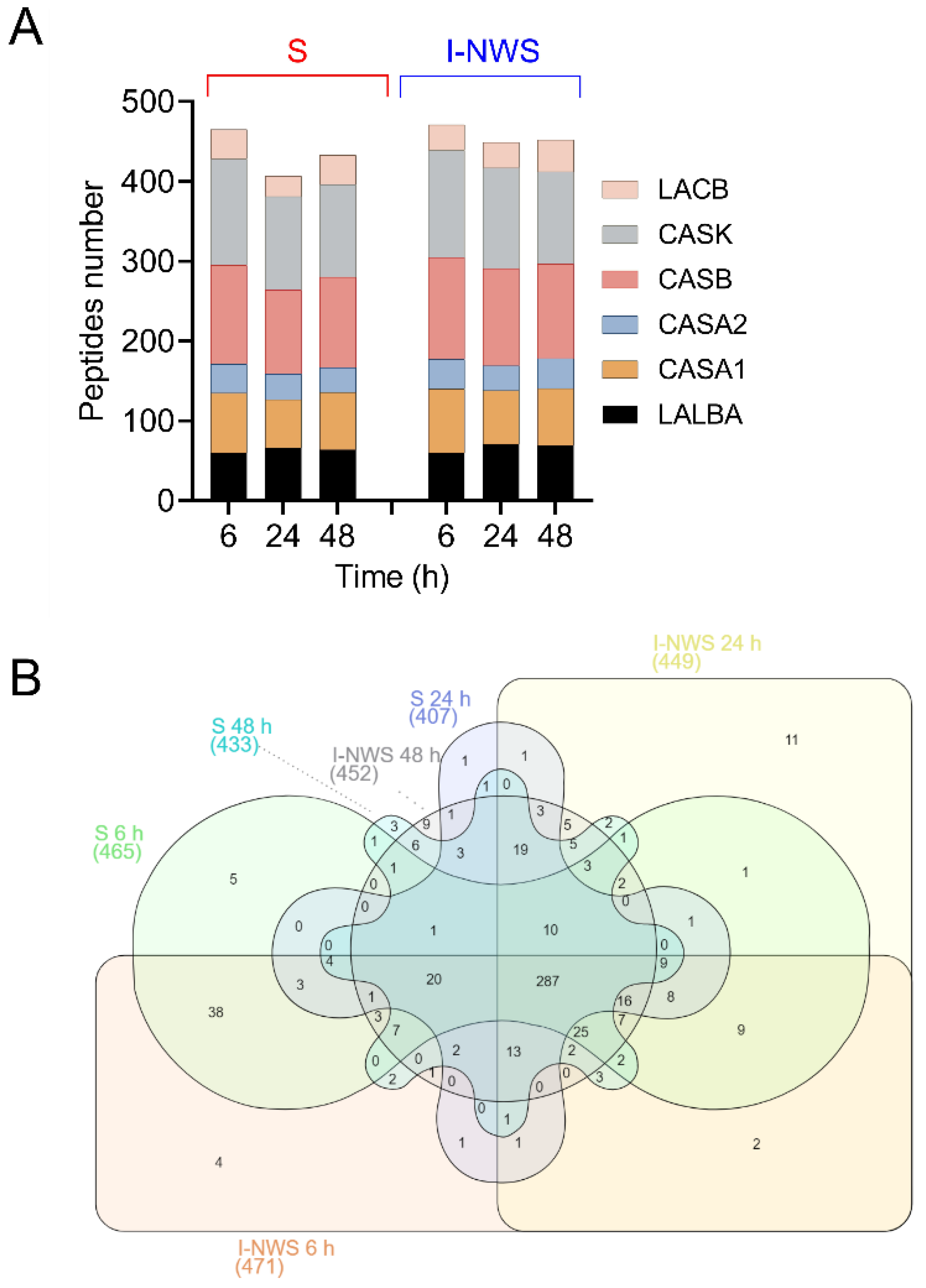
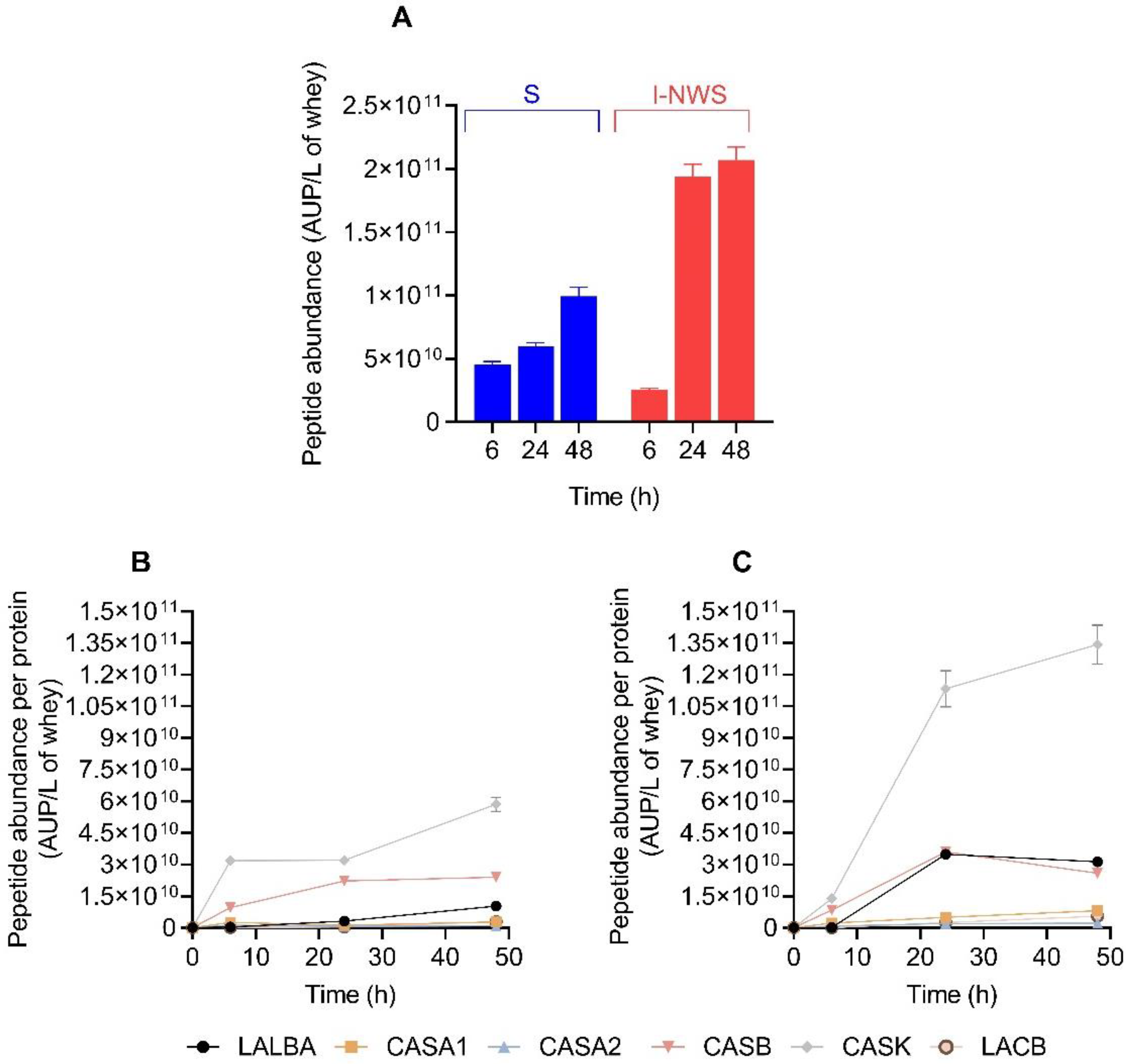
3.6. Peptidomics Profile of Fermented Cheese Whey
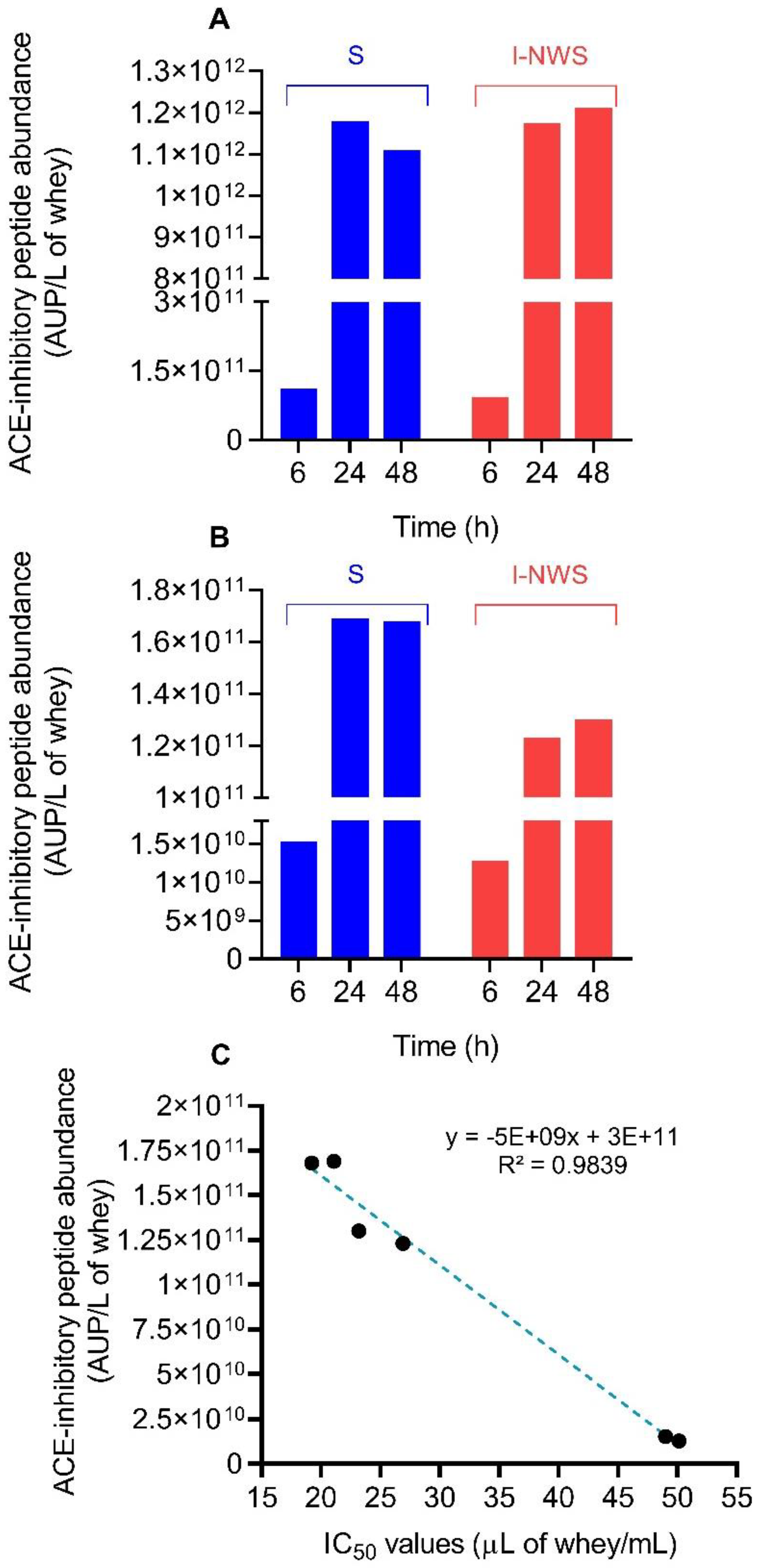
3.7. Bioactive Peptides Profile of Fermented Cheese Whey
3.8. Quantification of the Anti-Hypertensive Lactotripeptides VPP and IPP
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Timmis, K.; de Vos, W.M.; Ramos, J.L.; Vlaeminck, S.E.; Prieto, A.; Danchin, A.; Verstraete, W.; de Lorenzo, V.; Lee, S.Y.; Brüssow, H.; et al. The contribution of microbial biotechnology to sustainable development goals. Microbial. Biotechnol. 2017, 10, 984–987. [Google Scholar] [CrossRef] [PubMed]
- Hatti-Kaul, R.; Chen, L.; Dishisha, T.; Enshasy, H.E. Lactic acid bacteria: From starter cultures to producers of chemicals. FEMS Microbiol. Lett. 2018, 365, fny213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gänzle, M. The periodic table of fermented foods: Limitations and opportunities. Appl. Microbiol. Biotechnol. 2022, 106, 2815–2826. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, P.; Carvalho, A.L.; Vinga, S.; Santos, H.; Neves, A.R. From physiology to systems metabolic engineering for the production of biochemicals by lactic acid bacteria. Biotechnol. Adv. 2013, 31, 764–788. [Google Scholar] [CrossRef] [PubMed]
- López-Gómez, J.P.; Pérez-Rivero, C.; Venus, J. Valorisation of solid biowastes: The lactic acid alternative. Process Biochem. 2020, 99, 222–235. [Google Scholar] [CrossRef]
- Sharma, P.; Gaur, V.K.; Kim, S.H.; Pandey, A. Microbial strategies for bio-transforming food waste into resources. Bioresour. Technol. 2020, 299, 122580. [Google Scholar] [CrossRef]
- Khubber, S.; Marti-Quijal, F.J.; Tomasevic, I.; Remize, F.; Barba, F.J. Lactic acid fermentation as a useful strategy to recover antimicrobial and antioxidant compounds from food and by-products. Curr. Opin. Food Sci. 2022, 43, 189–198. [Google Scholar] [CrossRef]
- Raman, J.; Kim, J.S.; Choi, K.R.; Eun, H.; Yang, D.; Ko, Y.J.; Kim, S.J. Application of lactic acid bacteria (LAB) in sustainable agriculture: Advantages and limitations. Int. J. Mol. Sci. 2022, 23, 7784. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Lee, B.H.; Oh, D.H. Current trends and perspectives of bioactive peptides. Crit. Rev. Food Sci. Nutr. 2018, 58, 2273–2284. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Martini, S.; Solieri, L. Bioprospecting for bioactive peptide production by lactic acid bacteria isolated from fermented dairy food. Fermentation 2019, 5, 96. [Google Scholar] [CrossRef] [Green Version]
- Chourasia, R.; Chiring Phukon, L.; Abedin, M.M.; Padhi, S.; Singh, S.P.; Rai, A.K. Bioactive peptides in fermented foods and their application: A critical review. Syst. Microbiol. Biomanuf. 2023, 3, 88–109. [Google Scholar] [CrossRef]
- Agriculture, Forestry and Fishery Statistics, Eurostat, 2018 Edition. Available online: https://ec.europa.eu/eurostat/web/products-statistical-books/-/ks-fk-18-001 (accessed on 18 February 2023).
- Smithers, G.W. Whey and Whey Proteins—From ‘gutter-to-gold’. Int. Dairy J. 2008, 18, 695–704. [Google Scholar] [CrossRef]
- Jelen, P. Whey processing: Utilization and products. In Encyclopedia of Dairy Sciences, 1st ed.; Roginski, H., Fuquay, J.W., Fox, P.F., Eds.; Academic Press: New York, NY, USA, 2003; pp. 2739–2745. [Google Scholar]
- Dullius, A.; Goettert, M.I.; de Souza, C.F.V. Whey protein hydrolysates as a source of bioactive peptides for functional foods—Biotechnological facilitation of industrial scale-up. J. Funct. Foods 2018, 42, 58–74. [Google Scholar] [CrossRef]
- Zotta, T.; Solieri, L.; Iacumin, L.; Picozzi, C.; Gullo, M. Valorization of cheese whey using microbial fermentations. Appl. Microbiol. Biotechnol. 2020, 104, 2749–2764. [Google Scholar] [CrossRef]
- Pescuma, M.; Hébert, E.M.; Mozzi, F.; de Valdez, G.F. Functional fermented whey-based beverage using lactic acid bacteria. Int. J. Food Microbiol. 2010, 141, 73–81. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Lee, B.H.; Park, B.-J.; Kim, S.-H.; Oh, D.-H. Antihypertensive peptides from whey proteins fermented by lactic acid bacteria. Food Sci. Biotechnol. 2018, 27, 1781–1789. [Google Scholar] [CrossRef]
- Ali, E.; Nielsen, S.D.; Abd-El Aal, A.; El-Leboudy, A.; Saleh, E.; LaPointe, G. Use of mass spectrometry to profile peptides in whey protein isolate medium fermented by Lactobacillus helveticus LH-2 and Lactobacillus acidophilus La-5. Front. Nutr. 2019, 6, 152. [Google Scholar] [CrossRef] [Green Version]
- Skrzypczak, K.; Gustaw, W.; Fornal, E.; Kononiuk, A.; Michalak-Majewska, M.; Radzki, W.; Waśko, A. Functional and technological potential of whey protein isolate in production of milk beverages fermented by new strains of Lactobacillus helveticus. Appl. Sci. 2020, 10, 7089. [Google Scholar] [CrossRef]
- Mazorra-Manzano, M.A.; Robles-Porchas, G.R.; González-Velázquez, D.A.; Torres-Llanez, M.J.; Martínez-Porchas, M.; García-Sifuentes, C.O.; González-Córdova, A.F.; Vallejo-Córdoba, B. Cheese whey fermentation by its native microbiota: Proteolysis and bioactive peptides release with ACE-inhibitory activity. Fermentation 2020, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Mazorra-Manzano, M.A.; Robles-Porchas, G.R.; Martínez-Porchas, M.; Ramírez-Suárez, J.C.; García-Sifuentes, C.O.; Torres-Llanez, M.J.; González-Córdova, A.F.; Hernández-Mendoza, A.; Vallejo-Cordoba, B. Bacterial Diversity and Dynamics during Spontaneous Cheese Whey Fermentation at Different Temperatures. Fermentation 2022, 8, 342. [Google Scholar] [CrossRef]
- León-López, A.; Pérez-Marroquín, X.A.; Estrada-Fernández, A.G.; Campos-Lozada, G.; Morales-Peñaloza, A.; Campos-Montiel, R.G.; Aguirre-Álvarez, G. Milk Whey Hydrolysates as High Value-Added Natural Polymers: Functional Properties and Applications. Polymers 2022, 14, 1258. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.V.S.; Cappato, L.P.; Silva, R.; Rocha, R.S.; Guimarães, J.T.; Balthazar, C.F.; Esmerino, E.A.; Freitas, M.Q.; Rodrigues, F.N.; Granato, D.; et al. Ohmic heating for processing of whey-raspberry flavored beverage. Food Chem. 2019, 297, 125018. [Google Scholar] [CrossRef] [PubMed]
- Mucchettti, G.; Gatti, M.; Nocetti, M.; Reverberi, P.; Bianchi, A.; Galati, F.; Petroni, A. Segmentation of Parmigiano Reggiano dairies according to cheese-making technology and relationships with the aspect of the cheese curd surface at the moment of its extraction from the cheese vat. J. Dairy Sci. 2014, 97, 1202–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottari, B.; Agrimonti, C.; Gatti, M.; Neviani, E.; Marmiroli, N. Development of a multiplex real time PCR to detect thermophilic lactic acid bacteria in natural whey starters. Int. J. Food Microbiol. 2013, 160, 290–297. [Google Scholar] [CrossRef]
- Gatti, M.; Lazzi, C.; Rossetti, L.; Mucchetti, G.; Neviani, E. Biodiversity in Lactobacillus helveticus strains present in natural whey starter used for Parmigiano Reggiano cheese. J. Appl. Microbiol. 2003, 93, 463–470. [Google Scholar] [CrossRef]
- Rossetti, L.; Fornasari, M.E.; Gatti, M.; Lazzi, C.; Neviani, E.; Giraffa, G. Grana Padano cheese whey starters: Microbial composition and strain distribution. Int. J. Food Microbiol. 2008, 127, 168–171. [Google Scholar] [CrossRef]
- Santarelli, M.; Gatti, M.; Lazzi, C.; Bernini, V.; Zapparoli, G.A.; Neviani, E. Whey Starter for Grana Padano Cheese: Effect of technological parameters on viability and composition of the microbial community. J. Dairy Sci. 2008, 91, 883–891. [Google Scholar] [CrossRef] [Green Version]
- Sola, L.; Quadu, E.; Bortolazzo, E.; Bertoldi, L.; Randazzo, C.L.; Pizzamiglio, V.; Solieri, L. Insights on the bacterial composition of Parmigiano Reggiano Natural Whey Starter by a culture-dependent and 16S rRNA metabarcoding portrait. Sci. Rep. 2022, 12, 17322. [Google Scholar] [CrossRef]
- Solieri, L.; Valentini, M.; Cattivelli, A.; Sola, L.; Helal, A.; Martini, S.; Tagliazucchi, D. Fermentation of whey protein concentrate by Streptococcus thermophilus strains releases peptides with biological activities. Process Biochem. 2022, 121, 590–600. [Google Scholar] [CrossRef]
- Bertani, G.; Levante, A.; Lazzi, C.; Bottari, B.; Gatti, M.; Neviani, E. Dynamics of a natural bacterial community under technological and environmental pressures: The case of natural whey starter for Parmigiano Reggiano cheese. Food Res. Int. 2020, 129, 108860. [Google Scholar] [CrossRef]
- da Silva Duarte, V.; Carlot, M.; Pakroo, S.; Tarrah, A.; Lombardi, A.; Santiago, H.; Corich, V.; Giacomini, A. Comparative evaluation of cheese whey microbial composition from four Italian cheese factories by viable counts and 16S rRNA gene amplicon sequencing. Int. Dairy J. 2020, 104, 104656. [Google Scholar] [CrossRef]
- Martini, S.; Bonazzi, M.; Malorgio, I.; Pizzamiglio, V.; Tagliazucchi, D.; Solieri, L. Characterization of yeasts isolated from Parmigiano Reggiano cheese natural whey starter: From spoilage agents to potential cell factories for whey valorization. Microorganisms 2021, 9, 2288. [Google Scholar] [CrossRef] [PubMed]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzene sulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef] [PubMed]
- Tagliazucchi, D.; Shamsia, S.; Helal, A.; Conte, A. Angiotensin-converting enzyme inhibitory peptides from goats’ milk released by in vitro gastro-intestinal digestion. Int. Dairy J. 2017, 71, 6–16. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Rad. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Martini, S.; Shamsia, S.; Helal, A.; Conte, A. Biological activity and peptidomic profile of in vitro digested cow, camel, goat and sheep milk. Int. Dairy J. 2018, 81, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Martini, S.; Solieri, L.; Cattivelli, A.; Pizzamiglio, V.; Tagliazucchi, D. An Integrated Peptidomics and In Silico Approach to Identify Novel Anti-Diabetic Peptides in Parmigiano-Reggiano Cheese. Biology 2021, 10, 563. [Google Scholar] [CrossRef]
- Perkins, D.N.; Pappin, D.J.; Creasy, D.M.; Cottrell, J.S. Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophoresis 1999, 20, 3551–3567. [Google Scholar] [CrossRef]
- Dallas, D.; Nielsen, S.D. Milk peptidomics to identify functional peptides and for quality control of dairy products. Methods Mol. Biol. 2018, 1719, 223–240. [Google Scholar]
- MacLean, B.; Tomazela, D.M.; Shulman, N.; Chambers, M.; Finney, G.L.; Frewen, B.; Kern, R.; Tabb, D.L.; Liebler, D.C.; MacCoss, M.J. Skyline: An open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 2010, 26, 966–968. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, S.D.; Beverly, R.L.; Qu, Y.; Dallas, D.C. Milk bioactive peptide database: A comprehensive database of milk protein-derived bioactive peptides and novel visualization. Food Chem. 2017, 232, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Martini, S.; Conte, A.; Tagliazucchi, D. Effect of ripening and in vitro digestion on the evolution and fate of bioactive peptides in Parmigiano-Reggiano cheese. Int. Dairy J. 2020, 105, 104668. [Google Scholar] [CrossRef]
- Pescuma, M.; Valdez, G.F.D.; Mozi, F. Whey-derived valuable products obtained by microbial fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 6183–6196. [Google Scholar] [CrossRef] [PubMed]
- Pereira, Á.M.S.; de Farias, D.R.B.; de Queiroz, B.B.; Nobre, M.S.C.; Cavalcanti, M.T.; Salles, H.O.; Dos Santos, K.M.O.; de Medeiros, A.C.D.; Florentino, E.R.; Alonso Buriti, F.C. Influence of a Co-culture of Streptococcus thermophilus and Lactobacillus casei on the Proteolysis and ACE-Inhibitory Activity of a Beverage Based on Reconstituted Goat Whey Powder. Probiotics Antimicrob. Proteins 2019, 11, 273–282. [Google Scholar] [CrossRef]
- Worsztynowicz, P.; Białas, W.; Grajek, W. Integrated approach for obtaining bioactive peptides from whey proteins hydrolysed using a new proteolytic lactic acid bacteria. Food Chem. 2020, 312, 126035. [Google Scholar] [CrossRef]
- Yvon, M.; Chabanet, C.; Pélissier, J.P. Solubility of peptides in trichloroacetic acid (TCA) solutions. Hypothesis on the precipitation mechanism. Int. J. Pept. Protein Res. 1989, 34, 166–176. [Google Scholar] [CrossRef]
- Kieliszek, M.; Pobiega, K.; Piwowarek, K.; Kot, A.M. Characteristics of the proteolytic enzymes produced by lactic acid bacteria. Molecules 2021, 26, 1858. [Google Scholar] [CrossRef]
- Tavares, T.; Sevilla, M.Á.; Montero, M.J.; Carrón, R.; Malcata, F.X. Acute effect of whey peptides upon blood pressure of hypertensive rats, and relationship with their angiotensin-converting enzyme inhibitory activity. Mol. Nutr. Food Res. 2012, 56, 316–324. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Gerocarni, B.; Laghi, L.; Borghi, C. Blood pressure lowering effect of lactotripeptides assumed as functional foods: A meta-analysis of current available clinical trials. J. Hum. Hypertens. 2011, 25, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Fekete, A.A.; Givens, D.I.; Lovegrove, J.A. Casein-derived lactotripeptides reduce systolic and diastolic blood pressure in a meta-analysis of randomised clinical trials. Nutrients 2015, 7, 659–681. [Google Scholar] [CrossRef] [Green Version]
- Kohmura, M.; Nio, N.; Kubo, K.; Minoshima, Y.; Munekata, E.; Ariyoshi, Y. Inhibition of Angiotensin-converting Enzyme by Synthetic Peptides of Human β-Casein. Agric. Biol. Chem. 1989, 53, 2107–2114. [Google Scholar]
- Quirós, A.; Hernández-Ledesma, B.; Ramos, M.; Amigo, L.; Recio, I. Angiotensin-converting enzyme inhibitory activity of peptides derived from caprine kefir. J. Dairy Sci. 2005, 88, 3480–3487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Ledesma, B.; Recio, I.; Ramos, M.; Amigo, L. Preparation of ovine and caprine β-lactoglobulin hydrolysates with ACE-inhibitory activity. Identification of active peptides from caprine β-lactoglobulin hydrolysed with thermolysin. Int. Dairy J. 2002, 12, 805–812. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Helal, A.; Verzelloni, E.; Conte, A. Bovine milk antioxidant properties: Effect of in vitro digestion and identification of antioxidant compounds. Dairy Sci. Technol. 2016, 96, 657–676. [Google Scholar] [CrossRef] [Green Version]
- Tonolo, F.; Fiorese, F.; Moretto, L.; Folda, A.; Scalcon, V.; Grinzato, A.; Ferro, S.; Arrigoni, G.; Bindoli, A.; Feller, E.; et al. Identification of New Peptides from Fermented Milk Showing Antioxidant Properties: Mechanism of Action. Antioxidants 2020, 9, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amigo, L.; Martínez-Maqueda, D.; Hernández-Ledesma, B. In Silico and In Vitro Analysis of Multifunctionality of Animal Food-Derived Peptides. Foods 2020, 9, 991. [Google Scholar] [CrossRef]
- Sowmya, K.; Mala, D.; Bhat, M.I.; Kumar, N.; Bajaj, R.K.; Kapila, S.; Kapila, R. Bio-accessible milk casein derived tripeptide (LLY) mediates overlapping anti- inflammatory and anti-oxidative effects under cellular (Caco-2) and in vivo milieu. J. Nutr. Biochem. 2018, 62, 167–180. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Sequence 1 | Protein Fragment | Bioactivity | Sample |
|---|---|---|---|
| RELEELNVPGEIVESLSpSSpEESITR | β-casein (1–25) | Immunomodulatory, promote calcium uptake | I-NWS24h; S6h; S24h; S48h |
| FQSpEEQQQTEDELQDK | β-casein (33–48) | Promote calcium uptake | I-NWS6h; S6h |
| YPFP | β-casein (60–63) | Opioid, anti-cancer | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h |
| IPP | β-casein (74–76) κ-casein (108–110) | ACE-inhibitor (IC50 = 5 μmol/L), DPP-IV-inhibitor (IC50 = 169 μmol/L), antioxidant, anti-inflammatory | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| PVVVPPFLQPE | β-casein (81–91) | Anti-microbial | I-NWS6h; I-NWS48h; S6h; S24h; S48h |
| VPP | β-casein (84–86) | ACE-inhibitor (IC50 = 8 μmol/L), antioxidant, anti-inflammatory | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| NLHLPLPLL | β-casein (132–140) | ACE-inhibitor (IC50 = 15 μmol/L) | I-NWS24h; S48h |
| LHLPLPL | β-casein (133–139) | ACE-inhibitor (IC50 = 425 μmol/L) | I-NWS24h; I-NWS48h; S48h |
| LPLP | β-casein (135–138) | ACE-inhibitor (IC50 = 720 μmol/L) | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| PLP | β-casein (136–138) | ACE-inhibitor (IC50 = 430 μmol/L) | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| SQSKVLPVPQKAVPYPQ | β-casein (166–182) | Antioxidant | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| PYPQ | β-casein (179–182) | Antioxidant | I-NWS24h; I-NWS48h; S24h; S48h |
| RDMPIQAF | β-casein (183–190) | ACE-inhibitor (IC50 = 209 μmol/L) | I-NWS6h; I-NWS48h; S24h |
| LLY | β-casein (191–193) | Antioxidant, immunomodulatory, anti-inflammatory | I-NWS48h; S48h |
| YQEPVLGPVRGPFPIIV | β-casein (193–209) | ACE-inhibitor (IC50 = 101 μmol/L), immunomodulatory, anti-microbial | I-NWS6h; I-NWS24h; I-NWS48h; S48h |
| QEPVLGPVRGPFPIIV | β-casein (194–209) | ACE-inhibitor (IC50 = 600 μmol/L) | I-NWS48h; S24h |
| ENLLRF | αS1-casein (18–23) | ACE-inhibitor (IC50 = 82 μmol/L) | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| VPSERYL | αS1-casein (86–92) | ACE-inhibitor (IC50 = 250 μmol/L) | I-NWS24h; I-NWS48h; S48h |
| LRLKKYKVPQL | αS1-casein (99–109) | Anti-microbial | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| AYFYPEL | αS1-casein (143–149) | ACE-inhibitor (IC50 = 7 μmol/L), antioxidant, opioid | I-NWS24h; I-NWS48h |
| PEL | αS1-casein (147–149) | Antioxidant | I-NWS48h |
| VYQHQKAMKPWIQPKTKVIPYVRYL | αS2-casein (183–207) | Anti-microbial | I-NWS24h; I-NWS48h; S24h; S48h |
| TKVIPYVRYL | αS2-casein (198–207) | Anti-microbial | I-NWS6h; I-NWS24h; I-NWS48h; S24h; S48h |
| TKVIP | αS2-casein (198–207) | ACE-inhibitor (IC50 = 400 μmol/L) | I-NWS24h |
| VESTVATL | κ-casein (139–146) | Anti-microbial | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| STVATL | κ-casein (141–146) | Anti-microbial | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| PPEIN | κ-casein (156–160) | ACE-inhibitor (IC50 = 510 μmol/L) | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| VQVTSTAV | κ-casein (162–169) | Anti-microbial | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| AVF | β-lactoglobulin (80–82) | Anti-inflammatory | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| YLL | β-lactoglobulin (102–104) | ACE-inhibitor (IC50 = 519 μmol/L), antioxidant | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S48h |
| LLF | β-lactoglobulin (103–105) | ACE-inhibitor (IC50 = 80 μmol/L) | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| LKGYGGVSLPE | α-lactalbumin (15–25) | DPP-IV-inhibitory (IC50 = 486 μmol/L) | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| YGG | α-lactalbumin (18–20) | Immunomodulator | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| YGL | α-lactalbumin (50–52) | ACE-inhibitor (IC50 = 409 μmol/L) | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| YGLF | α-lactalbumin (50–53) | ACE-inhibitor (IC50 = 733 μmol/L) | I-NWS6h; I-NWS24h; I-NWS48h; S6h |
| DKVGINYW | α-lactalbumin (97–104) | ACE-inhibitor (IC50 = 25 μmol/L) | I-NWS6h; I-NWS24h; S6h |
| DKVGINY | α-lactalbumin (97–103) | ACE-inhibitor (IC50 = 100 μmol/L) | I-NWS6h; I-NWS24h; I-NWS48h; S24h; S48h |
| WLA | α-lactalbumin (104–106) | Anti-inflammatory | I-NWS6h; I-NWS24h; I-NWS48h; S24h; S48h |
| LAHKAL | α-lactalbumin (105–110) | ACE-inhibitor (IC50 = 621 μmol/L) | I-NWS6h; I-NWS24h; I-NWS48h; S6h; S24h; S48h |
| Sequence | I-NWS 6 h | I-NWS 24 h | I-NWS 48 h | S 6 h | S 24 h | S 48 h |
|---|---|---|---|---|---|---|
| VPP | 0.59 ± 0.05 a | 3.83 ± 0.11 b | 6.24 ± 0.52 c | 0.70 ± 0.07 a | 4.52 ± 0.31 d | 4.68 ± 0.39 d |
| IPP | 1.65 ± 0.12 a | 16.80 ± 1.01 b | 16.21 ± 1.23 b | 1.96 ± 0.18 a | 24.04 ± 1.99 c | 23.87 ± 2.01 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Helal, A.; Nasuti, C.; Sola, L.; Sassi, G.; Tagliazucchi, D.; Solieri, L. Impact of Spontaneous Fermentation and Inoculum with Natural Whey Starter on Peptidomic Profile and Biological Activities of Cheese Whey: A Comparative Study. Fermentation 2023, 9, 270. https://doi.org/10.3390/fermentation9030270
Helal A, Nasuti C, Sola L, Sassi G, Tagliazucchi D, Solieri L. Impact of Spontaneous Fermentation and Inoculum with Natural Whey Starter on Peptidomic Profile and Biological Activities of Cheese Whey: A Comparative Study. Fermentation. 2023; 9(3):270. https://doi.org/10.3390/fermentation9030270
Chicago/Turabian StyleHelal, Ahmed, Chiara Nasuti, Laura Sola, Giada Sassi, Davide Tagliazucchi, and Lisa Solieri. 2023. "Impact of Spontaneous Fermentation and Inoculum with Natural Whey Starter on Peptidomic Profile and Biological Activities of Cheese Whey: A Comparative Study" Fermentation 9, no. 3: 270. https://doi.org/10.3390/fermentation9030270
APA StyleHelal, A., Nasuti, C., Sola, L., Sassi, G., Tagliazucchi, D., & Solieri, L. (2023). Impact of Spontaneous Fermentation and Inoculum with Natural Whey Starter on Peptidomic Profile and Biological Activities of Cheese Whey: A Comparative Study. Fermentation, 9(3), 270. https://doi.org/10.3390/fermentation9030270