

Glucose Conversion for Biobutanol Production from Fresh Chlorella sorokiniana via Direct Enzymatic Hydrolysis
 and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Microalgae Cultivation and Harvest
2.2. Chemical Composition Analysis
2.3. Dilute Acid Pretreatment Followed by Enzymatic Hydrolysis
2.4. Direct Enzymatic Hydrolysis
2.5. Scanning Electron Microscopy Analysis
2.6. Biobutanol Fermentation
2.7. Analysis
2.8. Statistical Analysis
3. Results
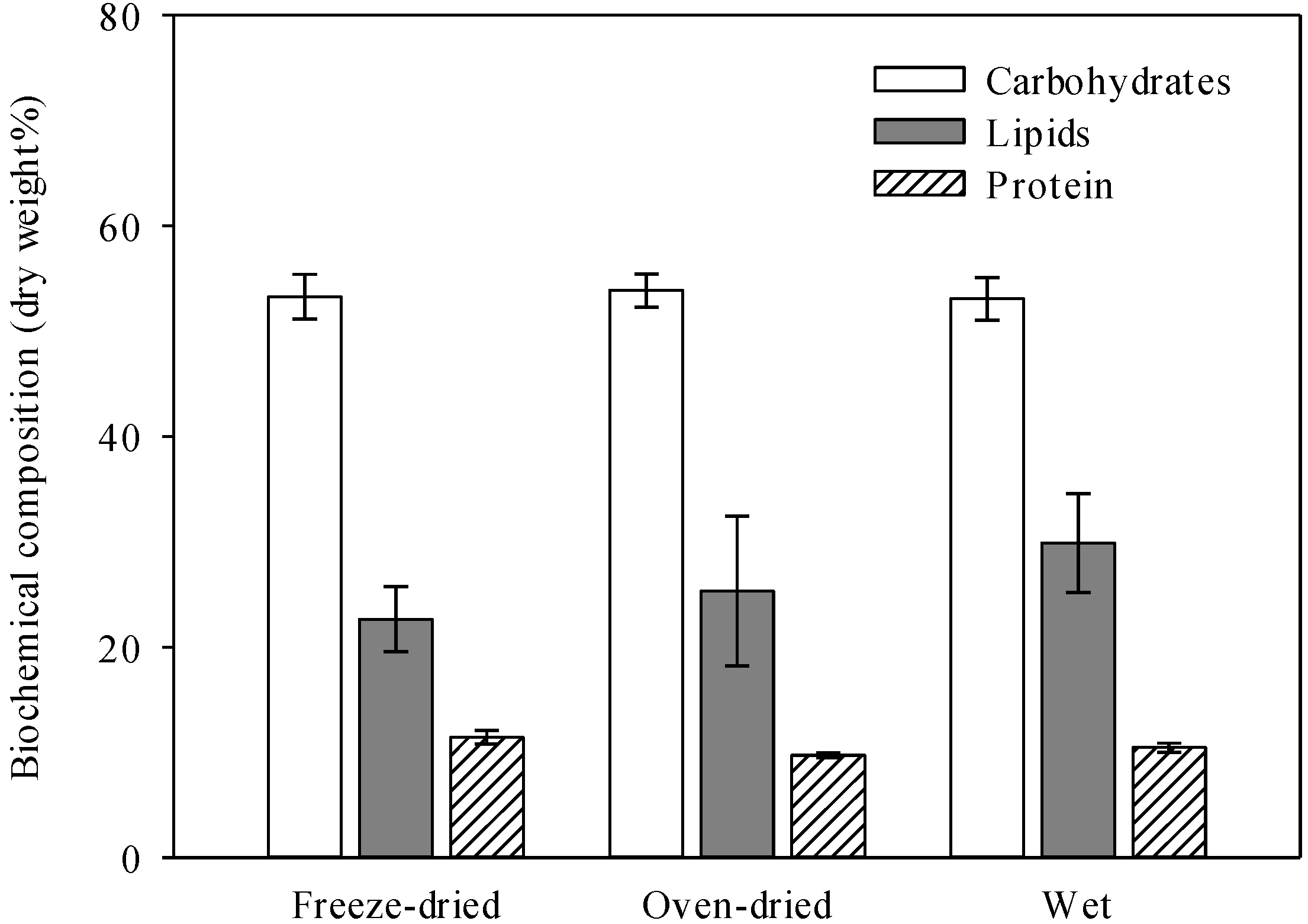
3.1. Wet and Dry Biomasses Used for Composition Analysis
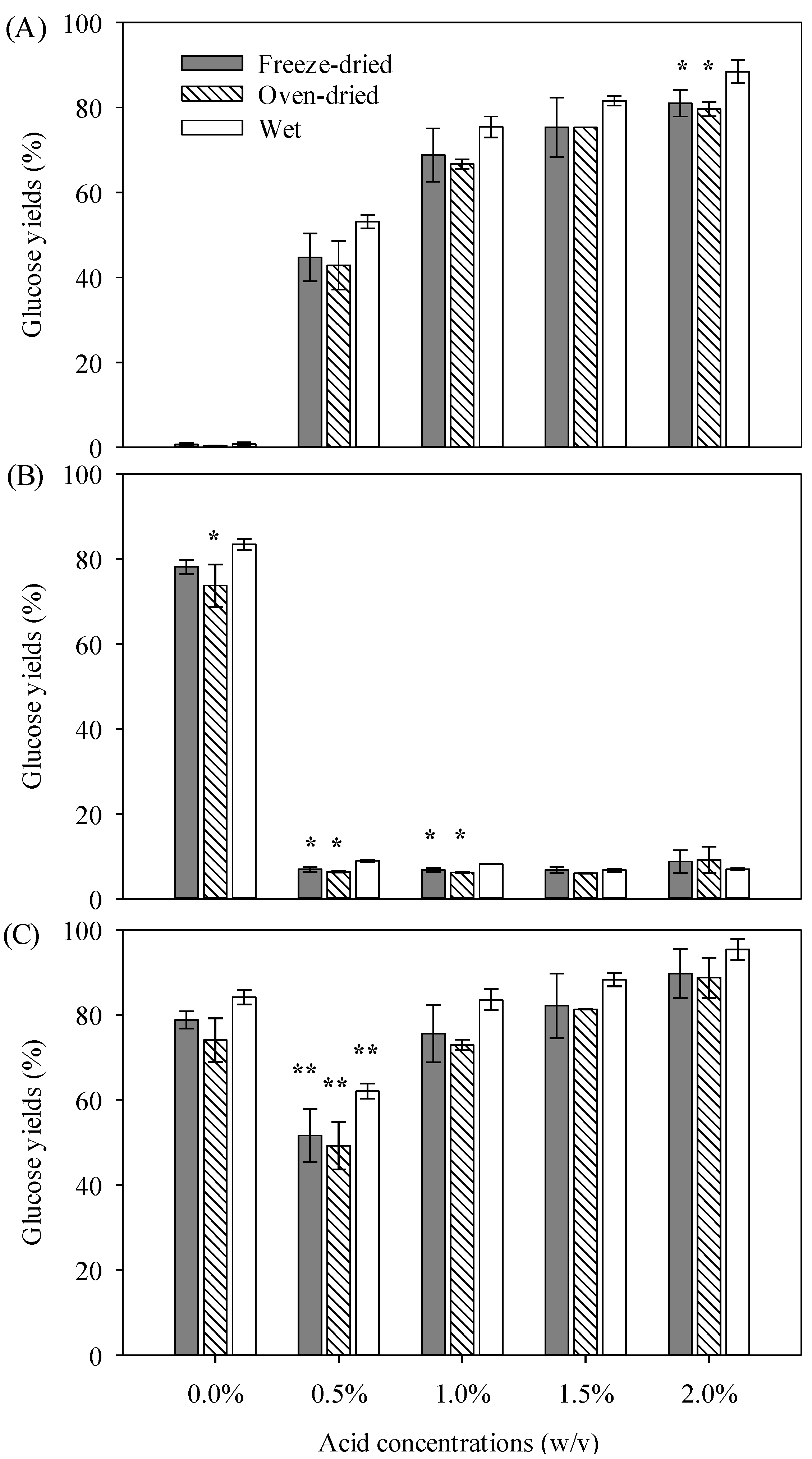
3.2. Acidic Pretreatment Followed by Enzymatic Hydrolysis
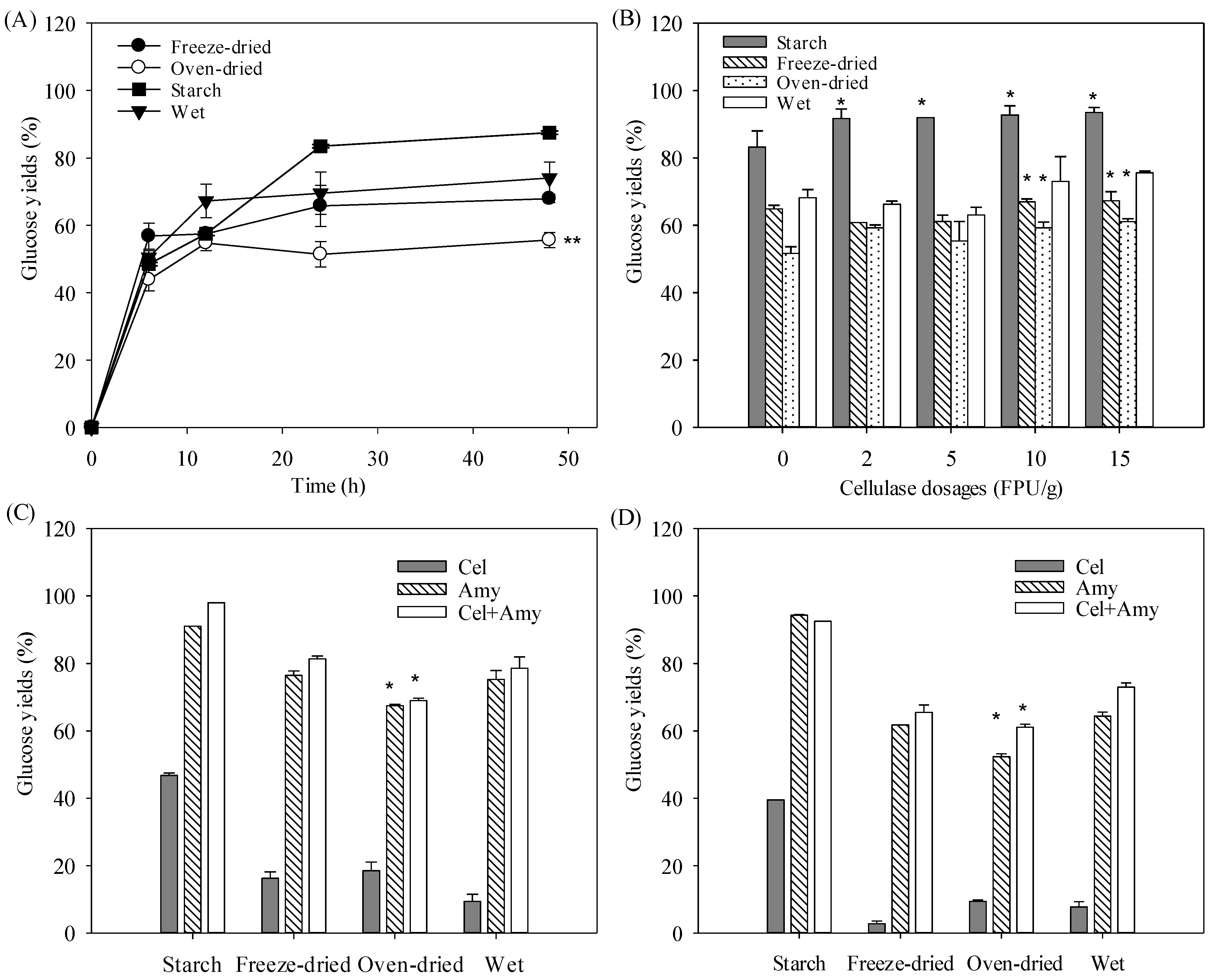
3.3. Direct Enzymatic Hydrolysis
3.4. Cellular Morphology of Dry and Wet Microalgal Biomass
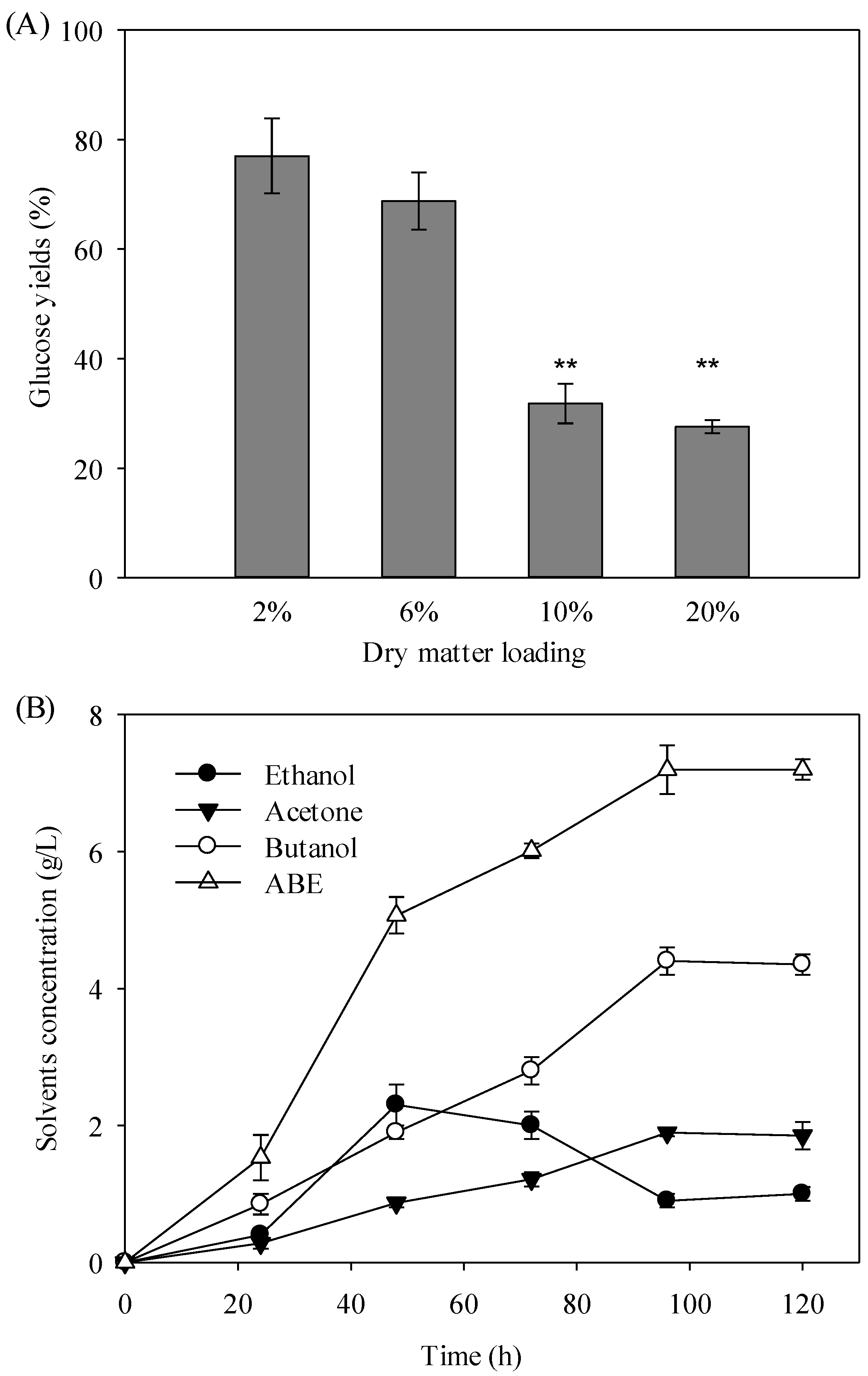
3.5. Biobutanol Fermentation of Wet Microalgal Hydrolysate
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Y.; Xu, H.; Yang, J.; Zhou, Y.; Wang, X.; Dou, S.; Li, L.; Liu, G.; Yang, M. Effect of sulfur limitation strategies on glucose-based carbohydrate production from Chlorella sorokiniana. Renew. Energy 2022, 200, 449–456. [Google Scholar] [CrossRef]
- Domozych, D.S.; Ciancia, M.; Fangel, J.U.; Mikkelsen, M.D.; Ulvskov, P.; Willats, W.G. The cell walls of green algae: A journey through evolution and diversity. Front. Plant Sci. 2012, 3, 82. [Google Scholar] [CrossRef] [PubMed]
- Demuez, M.; Mahdy, A.; Tomas-Pejo, E.; Gonzalez-Fernandez, C.; Ballesteros, M. Enzymatic cell disruption of microalgae biomass in biorefinery processes. Biotechnol. Bioeng. 2015, 112, 1955–1966. [Google Scholar] [CrossRef]
- Harun, R.; Danquah, M.K. Influence of acid pre-treatment on microalgal biomass for bioethanol production. Process Biochem. 2011, 46, 304–309. [Google Scholar] [CrossRef]
- Ho, S.H.; Huang, S.W.; Chen, C.Y.; Hasunuma, T.; Kondo, A.; Chang, J.S. Bioethanol production using carbohydrate-rich microalgae biomass as feedstock. Bioresour. Technol. 2013, 135, 191–198. [Google Scholar] [CrossRef] [PubMed]
- John, R.P.; Anisha, G.S.; Nampoothiri, K.M.; Pandey, A. Micro and macroalgal biomass: A renewable source for bioethanol. Bioresour. Technol. 2011, 102, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guo, W.; Cheng, C.L.; Ho, S.H.; Chang, J.S.; Ren, N. Enhancing bio-butanol production from biomass of Chlorella vulgaris JSC-6 with sequential alkali pretreatment and acid hydrolysis. Bioresour. Technol. 2016, 200, 557–564. [Google Scholar] [CrossRef]
- Naresh Kumar, A.; Chatterjee, S.; Hemalatha, M.; Althuri, A.; Min, B.; Kim, S.H.; Venkata Mohan, S. Deoiled algal biomass derived renewable sugars for bioethanol and biopolymer production in biorefinery framework. Bioresour. Technol. 2020, 296, 122315. [Google Scholar] [CrossRef]
- Ha, G.S.; El-Dalatony, M.M.; Kim, D.H.; Salama, E.S.; Kurade, M.B.; Roh, H.S.; El-Fatah Abomohra, A.; Jeon, B.H. Biocomponent-based microalgal transformations into biofuels during the pretreatment and fermentation process. Bioresour. Technol. 2020, 302, 122809. [Google Scholar] [CrossRef]
- Halim, R.; Harun, R.; Danquah, M.K.; Webley, P.A. Microalgal cell disruption for biofuel development. Appl. Energy 2012, 91, 116–121. [Google Scholar] [CrossRef]
- de Souza, M.F.; Rodrigues, M.A.; Freitas, S.P.; Bon, E.P.d.S. Effect of milling and enzymatic hydrolysis in the production of glucose from starch-rich Chlorella sorokiniana biomass. Algal Res. 2020, 50, 101961. [Google Scholar] [CrossRef]
- Zahoor; Wang, W.; Tan, X.; Guo, Y.; Zhang, B.; Chen, X.; Yu, Q.; Zhuang, X.; Yuan, Z. Mild Urea/KOH pretreatment to enhance enzymatic hydrolysis of corn stover with liquid waste recovery for plant growth. J. Clean. Prod. 2021, 284, 125392. [Google Scholar] [CrossRef]
- Zahoor; Wang, W.; Tan, X.; Imtiaz, M.; Wang, Q.; Miao, C.; Yuan, Z.; Zhuang, X. Rice straw pretreatment with KOH/urea for enhancing sugar yield and ethanol production at low temperature. Ind. Crops Prod. 2021, 170, 113776. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, W.; Zahoor; Tan, X.; Zhuang, X.; Miao, C.; Guo, Y.; Chen, X.; Yu, Q.; Yuan, Z. Recycling of Black Liquor for Treating Sugarcane Bagasse at Low Temperature to Attain High Ethanol Production without Washing Step. ACS Sustain. Chem. Eng. 2020, 8, 17016–17021. [Google Scholar] [CrossRef]
- Menegazzo, M.L.; Fonseca, G.G. Biomass recovery and lipid extraction processes for microalgae biofuels production: A review. Renew. Sustain. Energy Rev. 2019, 107, 87–107. [Google Scholar] [CrossRef]
- Chen, C.Y.; Zhao, X.Q.; Yen, H.W.; Ho, S.H.; Cheng, C.L.; Lee, D.J.; Bai, F.W.; Chang, J.S. Microalgae-based carbohydrates for biofuel production. Biochem. Eng. J. 2013, 78, 1–10. [Google Scholar] [CrossRef]
- Shokrkar, H.; Ebrahimi, S.; Zamani, M. Bioethanol production from acidic and enzymatic hydrolysates of mixed microalgae culture. Fuel 2017, 200, 380–386. [Google Scholar] [CrossRef]
- Wang, Y.; Ho, S.H.; Yen, H.W.; Nagarajan, D.; Ren, N.Q.; Li, S.; Hu, Z.; Lee, D.J.; Kondo, A.; Chang, J.S. Current advances on fermentative biobutanol production using third generation feedstock. Biotechnol. Adv. 2017, 35, 1049–1059. [Google Scholar] [CrossRef]
- de Carvalho Silvello, M.A.; Severo Gonçalves, I.; Patrícia Held Azambuja, S.; Silva Costa, S.; Garcia Pereira Silva, P.; Oliveira Santos, L.; Goldbeck, R. Microalgae-based carbohydrates: A green innovative source of bioenergy. Bioresour. Technol. 2022, 344, 126304. [Google Scholar] [CrossRef]
- Guldhe, A.; Singh, B.; Rawat, I.; Ramluckan, K.; Bux, F. Efficacy of drying and cell disruption techniques on lipid recovery from microalgae for biodiesel production. Fuel 2014, 128, 46–52. [Google Scholar] [CrossRef]
- Zhou, N.; Zhang, Y.; Wu, X.; Gong, X.; Wang, Q. Hydrolysis of Chlorella biomass for fermentable sugars in the presence of HCl and MgCl2. Bioresour. Technol. 2011, 102, 10158–10161. [Google Scholar] [CrossRef]
- Kim, K.H.; Choi, I.S.; Kim, H.M.; Wi, S.G.; Bae, H.-J. Bioethanol production from the nutrient stress-induced microalga Chlorella vulgaris by enzymatic hydrolysis and immobilized yeast fermentation. Bioresour. Technol. 2014, 153, 47–54. [Google Scholar] [CrossRef]
- Miranda, J.R.; Passarinho, P.C.; Gouveia, L. Pre-treatment optimization of Scenedesmus obliquus microalga for bioethanol production. Bioresour. Technol. 2012, 104, 342–348. [Google Scholar] [CrossRef]
- Pôjo, V.; Tavares, T.; Malcata, F.X. Processing methodologies of wet microalga biomass toward oil separation: An overview. Molecules 2021, 26, 641. [Google Scholar] [CrossRef]
- Ngamsirisomsakul, M.; Reungsang, A.; Liao, Q.; Kongkeitkajorn, M.B. Enhanced bio-ethanol production from Chlorella sp. biomass by hydrothermal pretreatment and enzymatic hydrolysis. Renew. Energy 2019, 141, 482–492. [Google Scholar] [CrossRef]
- Wang, X.; Zhou, Y.; Peng, Q.; Han, Y.; Yang, J.; Xu, H.; Li, C.; Li, L.; Dou, S.; Yang, M.; et al. Development of plastic flatbed-based algal culture system deployable on non-arable land. Algal Res. 2022, 66, 102814. [Google Scholar] [CrossRef]
- Wychen, S.V.; Laurens, L.M.L. Determination of Total Carbohydrates in Algal Biomass; NREL: Golden, CO, USA, 2013. [Google Scholar]
- Mishra, S.K.; Suh, W.I.; Farooq, W.; Moon, M.; Shrivastav, A.; Park, M.S.; Yang, J.W. Rapid quantification of microalgal lipids in aqueous medium by a simple colorimetric method. Bioresour. Technol. 2014, 155, 330–333. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, X.; Li, K.; Nan, Y.; Wang, J.; Zhang, J.; Dou, S.; Li, L.; Liu, G.; Yang, M. Comparison of a solvent mixture assisted dilute acid and alkali pretreatment in sugar production from hybrid Pennisetum. Ind. Crops Prod. 2019, 141, 111806. [Google Scholar] [CrossRef]
- Yang, M.; Kuittinen, S.; Vepsalainen, J.; Zhang, J.; Pappinen, A. Enhanced acetone-butanol-ethanol production from lignocellulosic hydrolysates by using starchy slurry as supplement. Bioresour. Technol. 2017, 243, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, A.; Grinsted, E. Methods for the growth and enumeration of anaerobic spore-formers from cheese, with observations on the effect of nisin. J. Dairy Res. 1954, 21, 101–110. [Google Scholar] [CrossRef]
- Cai, D.; Zhang, T.; Zheng, J.; Chang, Z.; Wang, Z.; Qin, P.Y.; Tan, T.W. Biobutanol from sweet sorghum bagasse hydrolysate by a hybrid pervaporation process. Bioresour. Technol. 2013, 145, 97–102. [Google Scholar] [CrossRef]
- Dragone, G.; Fernandes, B.D.; Abreu, A.P.; Vicente, A.A.; Teixeira, J.A. Nutrient limitation as a strategy for increasing starch accumulation in microalgae. Appl. Energy 2011, 88, 3331–3335. [Google Scholar] [CrossRef]
- Ran, W.; Wang, H.; Liu, Y.; Qi, M.; Xiang, Q.; Yao, C.; Zhang, Y.; Lan, X. Storage of starch and lipids in microalgae: Biosynthesis and manipulation by nutrients. Bioresour. Technol. 2019, 291, 121894. [Google Scholar] [CrossRef]
- Yang, M.; Kuittinen, S.; Zhang, J.; Keinanen, M.; Pappinen, A. Effect of dilute acid pretreatment on the conversion of barley straw with grains to fermentable sugars. Bioresour. Technol. 2013, 146, 444–450. [Google Scholar] [CrossRef]
- Thu, N.M.; Choi, S.P.; Lee, J.; Lee, J.H.; Sim, S.J. Hydrothermal acid pretreatment of Chlamydomonas reinhardtii biomass for ethanol production. J. Microbiol. Biotechnol. 2009, 19, 161–166. [Google Scholar] [CrossRef]
- Kumar, V.B.; Pulidindi, I.N.; Kinel-Tahan, Y.; Yehoshua, Y.; Gedanken, A. Evaluation of the potential of Chlorella vulgaris for bioethanol production. Energy Fuels 2016, 30, 3161–3166. [Google Scholar] [CrossRef]
- Harun, R.; Jason, W.S.Y.; Cherrington, T.; Danquah, M.K. Exploring alkaline pre-treatment of microalgal biomass for bioethanol production. Appl. Energy 2011, 88, 3464–3467. [Google Scholar] [CrossRef]
- Onay, M. Bioethanol production via different saccharification strategies from H. tetrachotoma ME03 grown at various concentrations of municipal wastewater in a flat-photobioreactor. Fuel 2019, 239, 1315–1323. [Google Scholar] [CrossRef]
- Ho, S.H.; Li, P.J.; Liu, C.C.; Chang, J.S. Bioprocess development on microalgae-based CO2 fixation and bioethanol production using Scenedesmus obliquus CNW-N. Bioresour. Technol. 2013, 145, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.P.; Nguyen, M.T.; Sim, S.J. Enzymatic pretreatment of Chlamydomonas reinhardtii biomass for ethanol production. Bioresour. Technol. 2010, 101, 5330–5336. [Google Scholar] [CrossRef]
- Harun, R.; Danquah, M.K. Enzymatic hydrolysis of microalgal biomass for bioethanol production. Chem. Eng. J. 2011, 168, 1079–1084. [Google Scholar] [CrossRef]
- de Farias Silva, C.E.; Meneghello, D.; Bertucco, A. A systematic study regarding hydrolysis and ethanol fermentation from microalgal biomass. Biocatal. Agric. Biotechnol. 2018, 14, 172–182. [Google Scholar] [CrossRef]
- Alam, M.A.; Yuan, T.; Xiong, W.; Zhang, B.; Lv, Y.; Xu, J. Process optimization for the production of high-concentration ethanol with Scenedesmus raciborskii biomass. Bioresour. Technol. 2019, 294, 122219. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Sun, J.; Huang, Y.; Feng, J.; Zhou, J.; Cen, K. Dynamic microstructures and fractal characterization of cell wall disruption for microwave irradiation-assisted lipid extraction from wet microalgae. Bioresour. Technol. 2013, 150, 67–72. [Google Scholar] [CrossRef]
- Hernández, D.; Riaño, B.; Coca, M.; García-González, M.C. Saccharification of carbohydrates in microalgal biomass by physical, chemical and enzymatic pre-treatments as a previous step for bioethanol production. Chem. Eng. J. 2015, 262, 939–945. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, P.; Wang, Y.; Liu, S.; Wang, Q.; Wang, Y. Fermentable Sugar Production from Wet Microalgae Residual after Biodiesel Production Assisted by Radio Frequency Heating. Renew. Energy. 2020, 155, 827–836. [Google Scholar] [CrossRef]
- van der Wal, H.; Sperber, B.L.; Houweling-Tan, B.; Bakker, R.R.; Brandenburg, W.; Lopez-Contreras, A.M. Production of acetone, butanol, and ethanol from biomass of the green seaweed Ulva lactuca. Bioresour. Technol. 2013, 128, 431–437. [Google Scholar] [CrossRef]
- Ellis, J.T.; Hengge, N.N.; Sims, R.C.; Miller, C.D. Acetone, butanol, and ethanol production from wastewater algae. Bioresour. Technol. 2012, 111, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Efremenko, E.N.; Nikolskaya, A.B.; Lyagin, I.V.; Senko, O.V.; Makhlis, T.A.; Stepanov, N.A.; Maslova, O.V.; Mamedova, F.; Varfolomeev, S.D. Production of biofuels from pretreated microalgae biomass by anaerobic fermentation with immobilized Clostridium acetobutylicum cells. Bioresour. Technol. 2012, 114, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Ezeji, T.C.; Qureshi, N.; Blaschek, H.P. Production of acetone butanol (AB) from liquefied corn starch, a commercial substrate, using Clostridium beijerinckii coupled with product recovery by gas stripping. J. Ind. Microbiol. Biotechnol. 2007, 34, 771–777. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | Source | Activity | Optimum Temperature (°C) | Optimum pH |
|---|---|---|---|---|
| α-amylase | Aspergillus oryzae | 40,000 µ/g | 55–100 | 5.2–6.0 |
| Amyloglucosidase | Aspergillus niger | 100,000 µ/mL | 58–62 | 4.2–4.5 |
| Cellulase | A blend of aggressive cellulases, high level of β-glucosidases and hemicellulase | 176 FPU/mL | 45–50 | 4.5–5.0 |
| Microalgae | Substrates | Pretreatment/Hydrolysis Methods | Glucose Yields (%) | References |
|---|---|---|---|---|
| Chlorella vulgaris FSP-E | Lyophilized | 1% (v/v) H2SO4 at 120 °C for 20 min | 93.6 | [5] |
| Chlorella sp. TIB-A01 | Dried at 60 °C | 2% HCl and 2.5% MgCl2 at 180 °C for 10 min | 64.21 | [21] |
| Scenedesmus obliquus | Oven-dried at 80 °C | 2 N H2SO4 at 120 °C for 30 min | 95.6 | [23] |
| Chlorella vulgaris | Dried in a sunbath for 24 h | 1 M HCl at 120 °C for 60 min | 23 | [37] |
| Chlorococcum infusionum | Oven dried at 60 °C overnight and milled | 0.75% (w/v) NaOH at 120 °C for 30 min | 350 mg/g Biomass | [38] |
| Hindakia tetrachotoma ME03 | Lyophilized | 1M HCl at 121 °C for 30 min | 90.3 | [39] |
| Scenedesmus obliquus CNW-N | Wet | 2% (v/v) H2SO4 at 120 °C for 20 min | 96–98 | [40] |
| Chlorella sorokiniana UTEX 1230 | Wet | 2% (w/v) H2SO4 at 120 °C for 60 min | 88.5 | This study |
| Microalgae | Substrates | Pretreatments | Enzymes | Glucose Yields (%) | References |
|---|---|---|---|---|---|
| Chlorella vulgaris FSP-E | Lyophilized | Sonication and autoclave | Endoglucanase, β-glucosidase and amylase | 90.4 | [5] |
| Chlorella sorokiniana UTEX 1663 | Freeze-dried Wet | Milling | Cellulase and amylase | 80.14 11.56 | [11] |
| Chlorella Vulgaris UTEX 26 | Freeze- dried | Autoclave, sonication, and milling | Pectinase | 79 | [22] |
| Chlorella vulgaris | Dried | Milled | Amylases, cellulase/hemicellulase, and pectinases | 92 | [43] |
| Scenedesmus raciborskii ZKMT | Freeze- dried | Ground and sieving | Cellulase, α-amylase and amyloglucosidase | 91.47 | [44] |
| Chlorella variabilis NC64A | Wet | Virus infection | α-amylase and amyloglucosidase | 43.4 | [45] |
| Chlorellasorokiniana UTEX 1230 | Wet | Autoclave | α-amylase and amyloglucosidase | 83.4 | This study |
| Chlorella sorokiniana UTEX 1230 | Wet | No | α-amylase and amyloglucosidase | 78.7 | This study |
| Dry Matter Loading (%) | Glucose Concentrations (g/L) |
|---|---|
| 2 | 8.0 ± 0.7 |
| 6 | 23.0 ± 1.4 |
| 10 | 16.5 ± 1.9 |
| 20 | 28.6 ± 1.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Cai, D.; Liu, X.; Zhu, L.; Zhang, C.; Peng, Q.; Han, Y.; Liu, G.; Yang, M. Glucose Conversion for Biobutanol Production from Fresh Chlorella sorokiniana via Direct Enzymatic Hydrolysis. Fermentation 2023, 9, 284. https://doi.org/10.3390/fermentation9030284
Yang J, Cai D, Liu X, Zhu L, Zhang C, Peng Q, Han Y, Liu G, Yang M. Glucose Conversion for Biobutanol Production from Fresh Chlorella sorokiniana via Direct Enzymatic Hydrolysis. Fermentation. 2023; 9(3):284. https://doi.org/10.3390/fermentation9030284
Chicago/Turabian StyleYang, Jinzhi, Di Cai, Xudong Liu, Liqi Zhu, Changwei Zhang, Qing Peng, Yanxia Han, Guozhen Liu, and Ming Yang. 2023. "Glucose Conversion for Biobutanol Production from Fresh Chlorella sorokiniana via Direct Enzymatic Hydrolysis" Fermentation 9, no. 3: 284. https://doi.org/10.3390/fermentation9030284
APA StyleYang, J., Cai, D., Liu, X., Zhu, L., Zhang, C., Peng, Q., Han, Y., Liu, G., & Yang, M. (2023). Glucose Conversion for Biobutanol Production from Fresh Chlorella sorokiniana via Direct Enzymatic Hydrolysis. Fermentation, 9(3), 284. https://doi.org/10.3390/fermentation9030284