
The Effect of Lactiplantibacillus plantarum ZZU203, Cellulase-Producing Bacillus methylotrophicus, and Their Combinations on Alfalfa Silage Quality and Bacterial Community

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Silage Preparation
2.2. Analysis of Fermentation Parameters and Chemical Composition
2.3. Bacterial Community Analyses
2.4. Statistical Analyses
3. Results and Discussion
3.1. Chemical Characteristics of Alfalfa before Ensiling
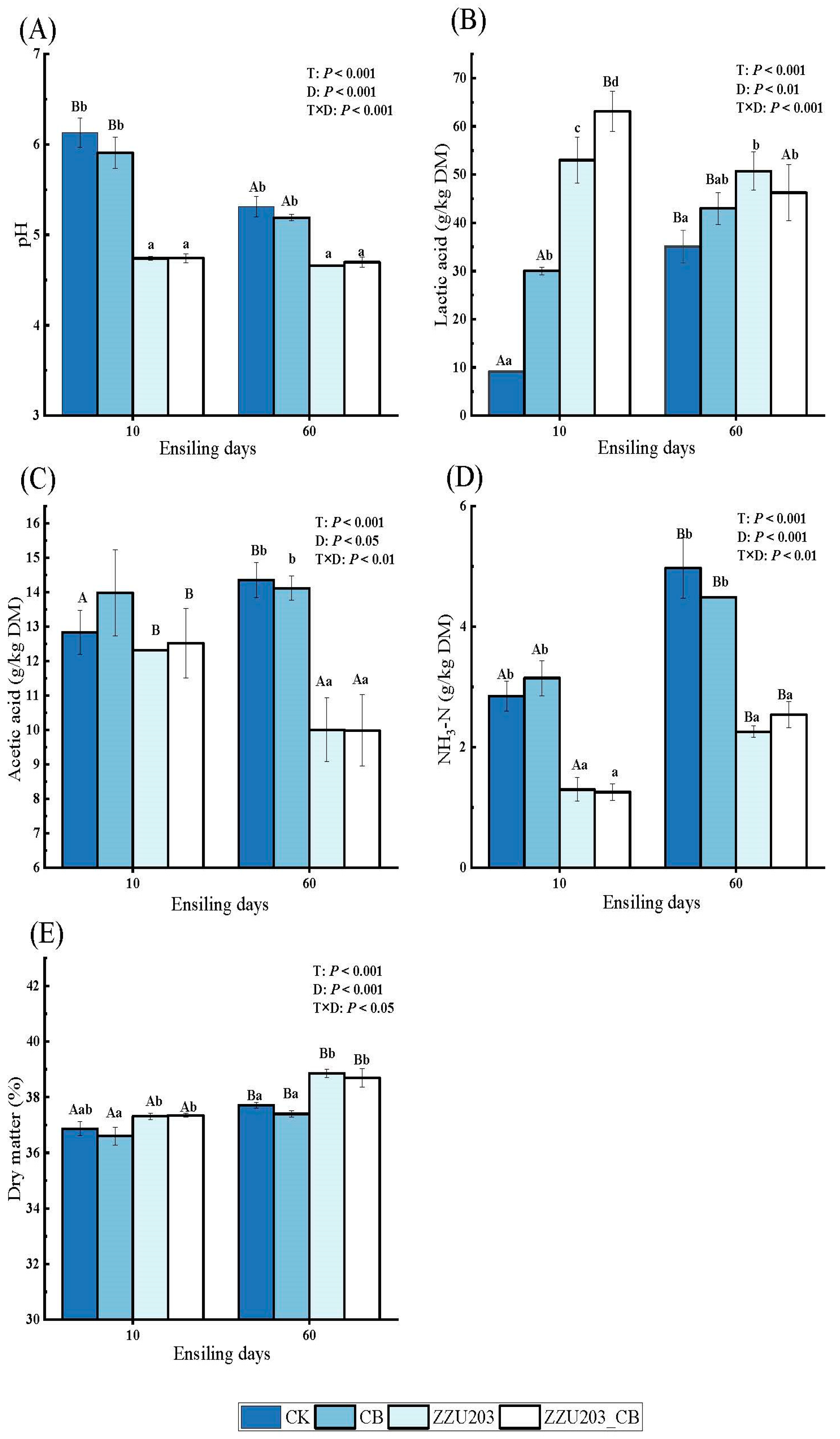
3.2. The Fermentation Parameters of Alfalfa Silage after Ensiling for 60 Days
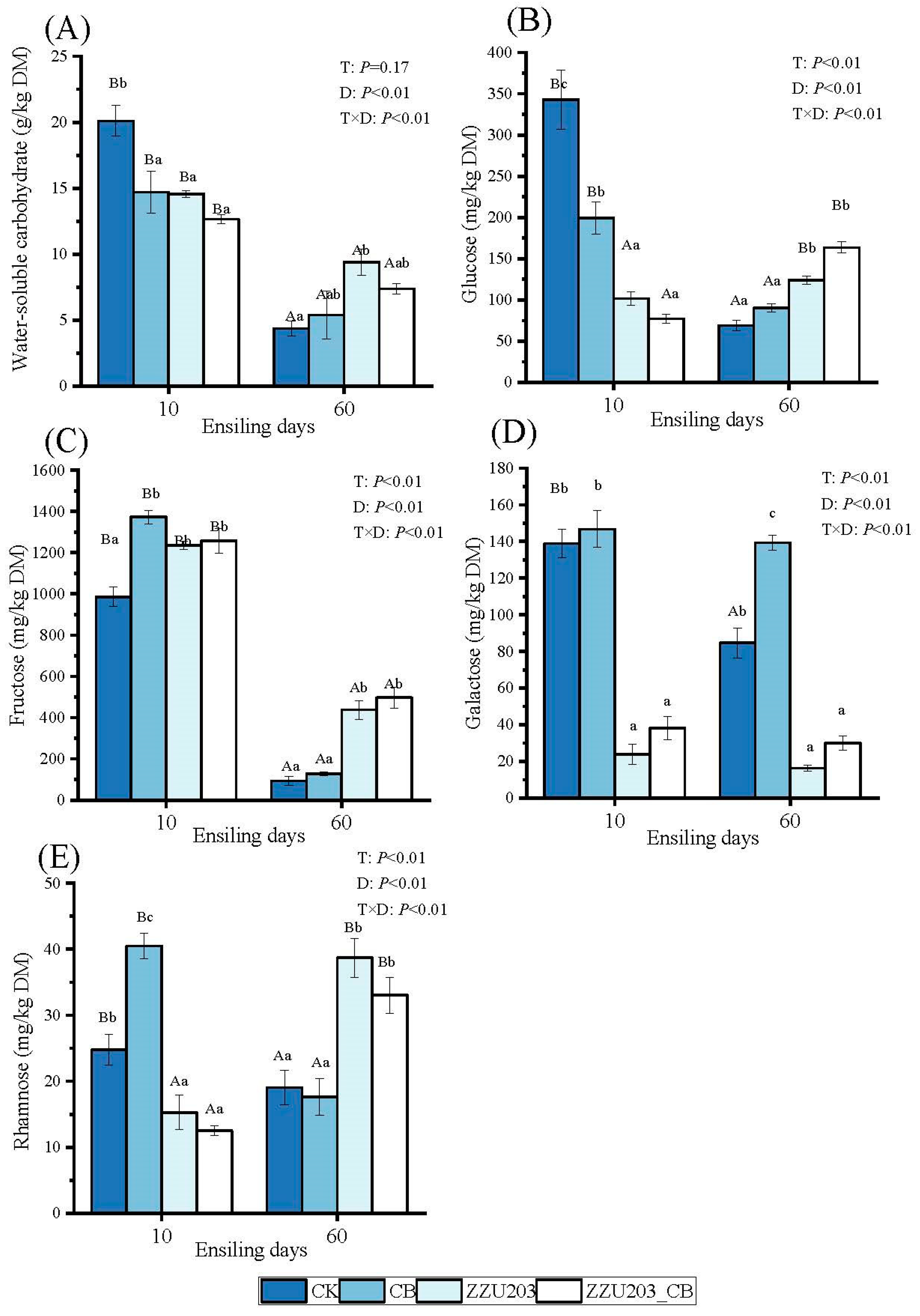
3.3. Nonstructural Carbohydrate Components of Alfalfa Silage
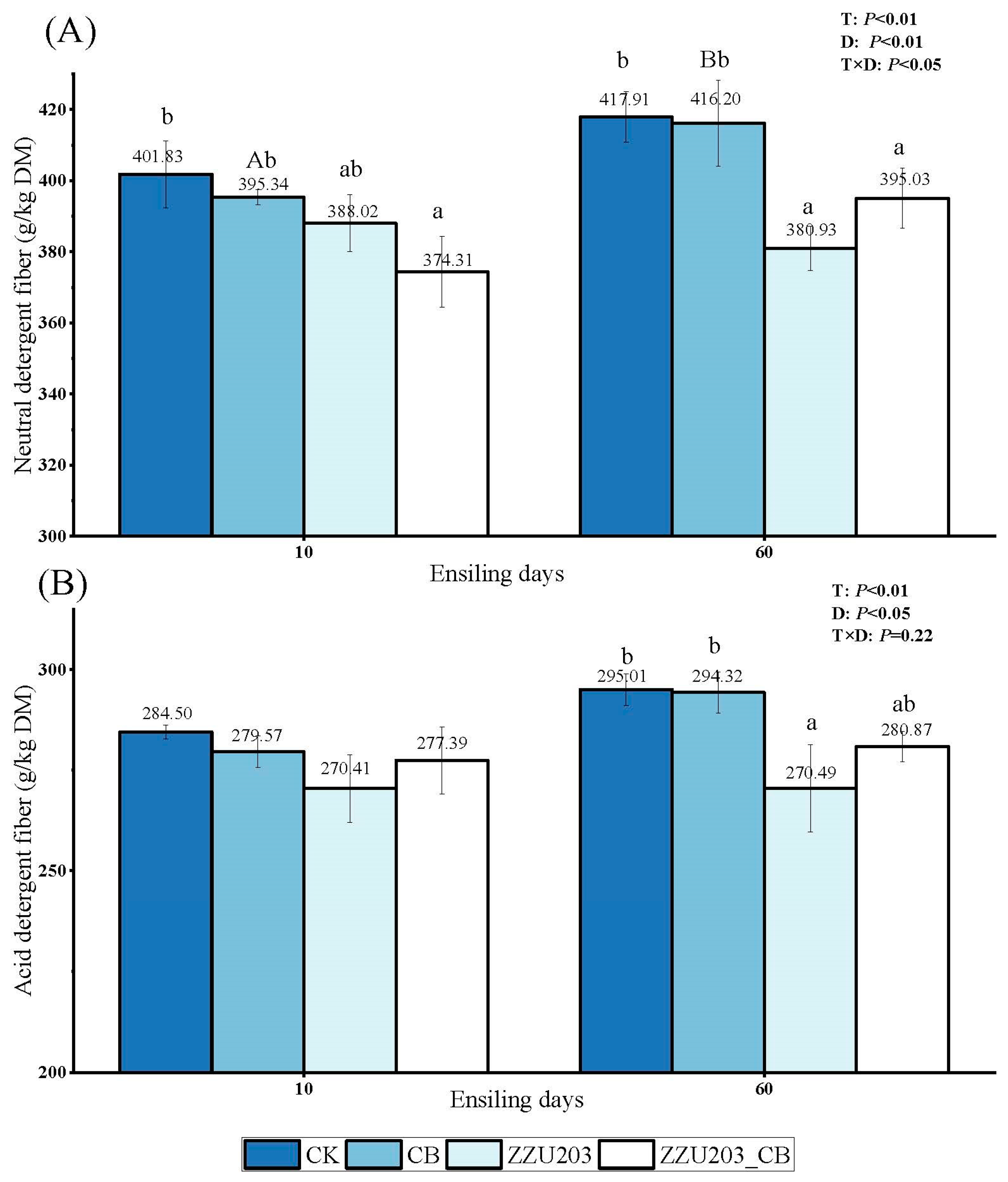
3.4. Structural Carbohydrate Components of Alfalfa Silage
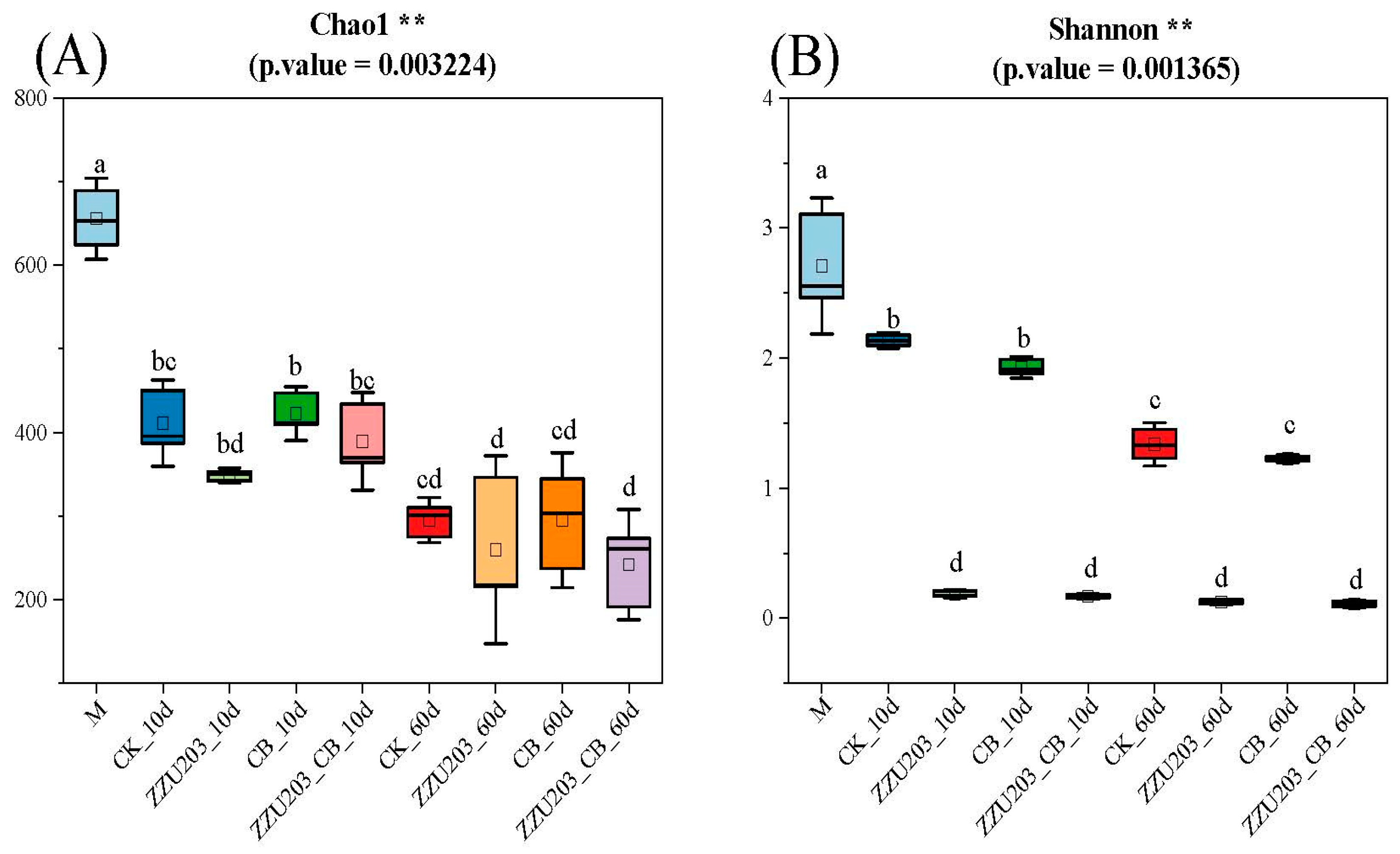
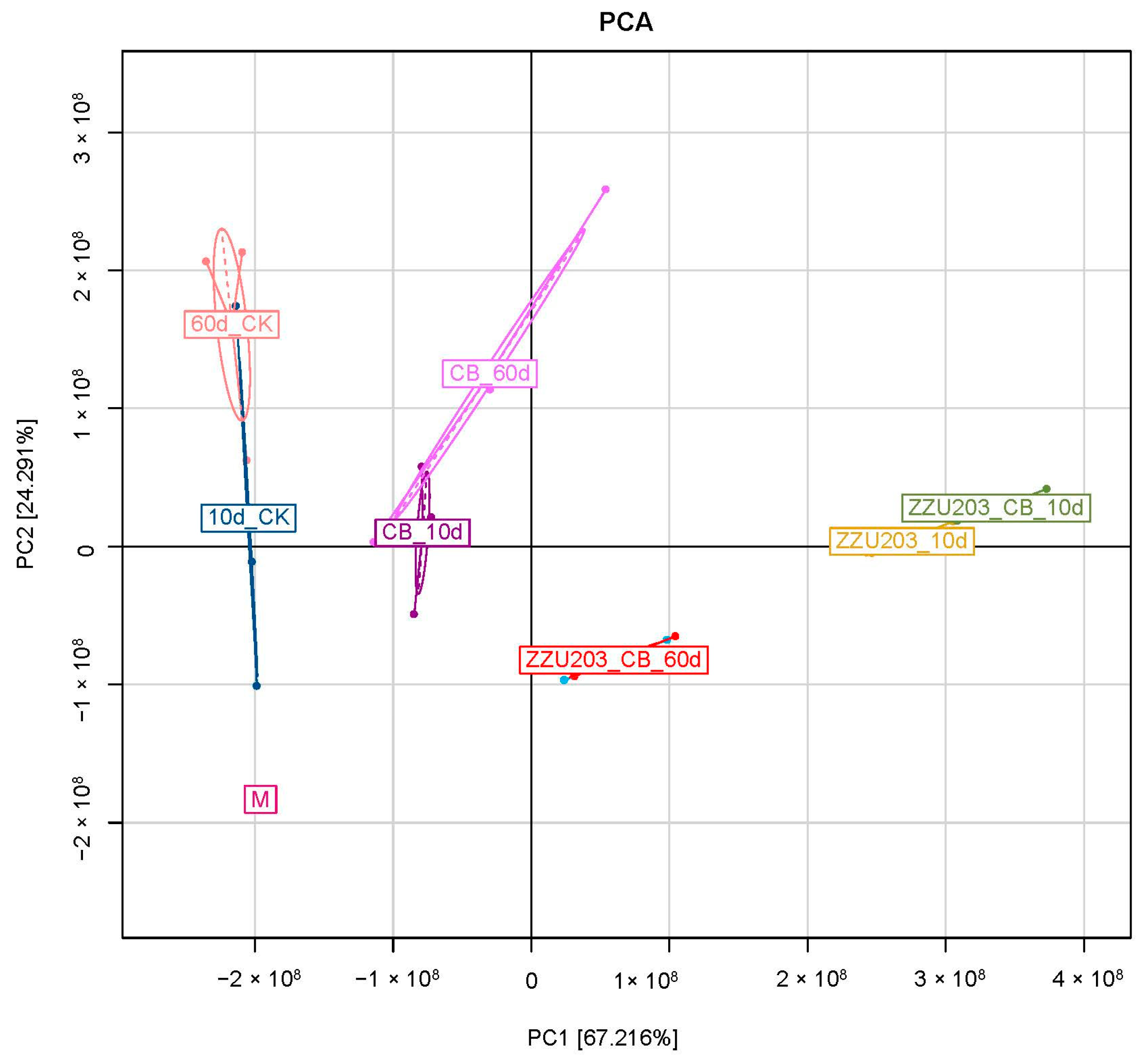
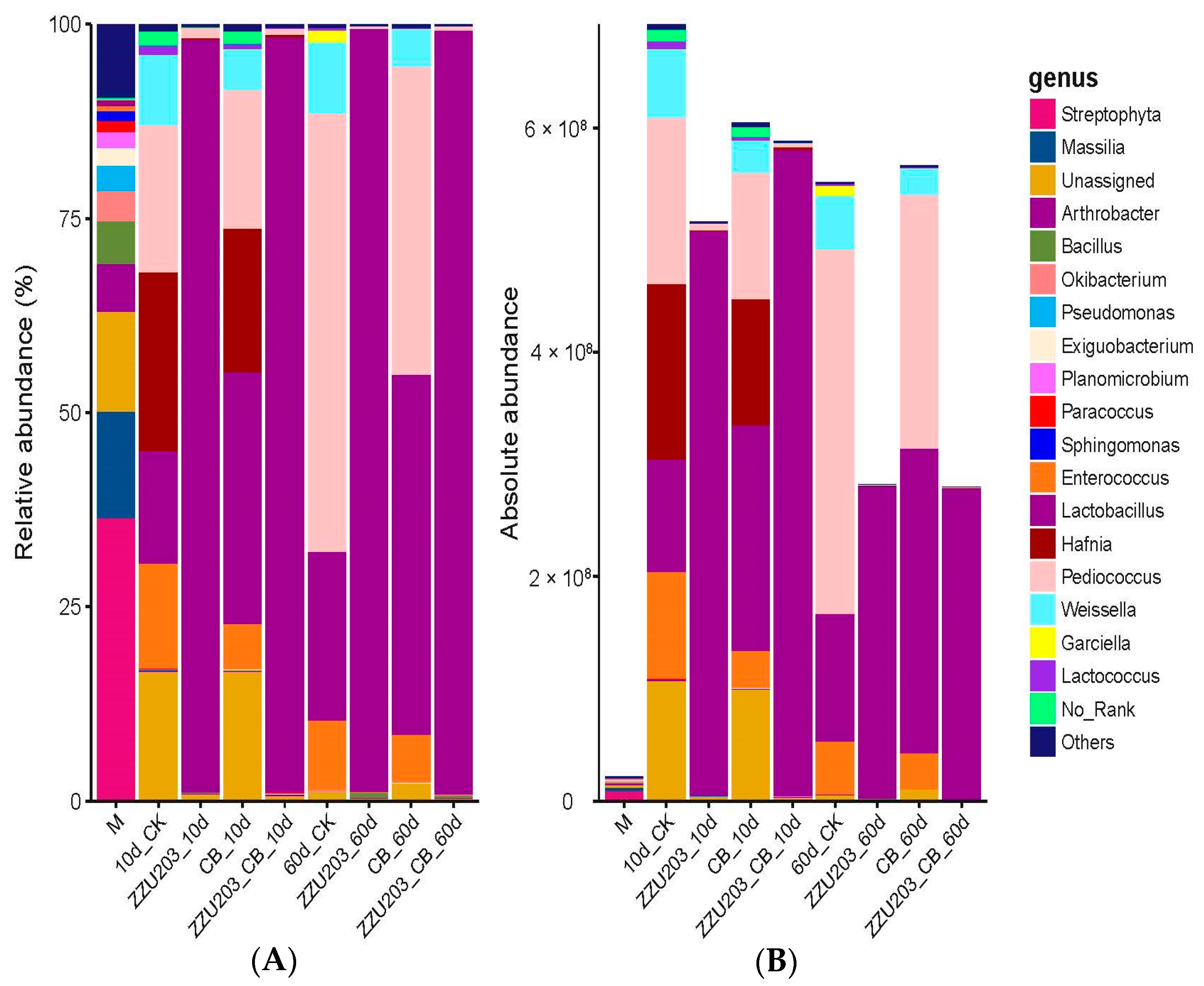
3.5. Bacterial Community after 60 d of Ensiling
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coblentz, W.K.; Muck, R.E. Effects of natural and simulated rainfall on indicators of ensilability and nutritive value for wilting alfalfa forages sampled before preservation as silage. J. Dairy Sci. 2012, 95, 6635–6653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadeau, E.M.G.; Buxton, D.R.; Russell, J.R.; Allison, M.J.; Young, J.W. Enzyme, Bacterial Inoculant, and Formic Acid Effects on Silage Composition of Orchardgrass and Alfalfa1, 2. J. Dairy Sci. 2000, 83, 1487–1502. [Google Scholar] [CrossRef] [PubMed]
- Ke, W.C.; Ding, W.R.; Xu, D.M.; Ding, L.M.; Zhang, P.; Li, F.D.; Guo, X.S. Effects of addition of malic or citric acids on fermentation quality and chemical characteristics of alfalfa silage. J. Dairy Sci. 2017, 100, 8958–8966. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.J.; Wen, A.Y.; Wang, J.; Desta, S.T.; Dong, Z.H.; Shao, T. Effects of four short-chain fatty acids or salts on fermentation characteristics and aerobic stability of alfalfa (Medicago sativa L.) silage. J. Sci. Food Agric. 2018, 98, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Sheperd, A.C.; Maslanka, M.; Quinn, D.; Kung, L. Additives Containing Bacteria and Enzymes for Alfalfa Silage1. J. Dairy Sci. 1995, 78, 565–572. [Google Scholar] [CrossRef]
- Li, J.F.; Yuan, X.J.; Desta, S.T.; Dong, Z.H.; Mugabe, W.; Shao, T. Characterization of Enterococcus faecalis JF85 and Enterococcus faecium Y83 isolated from Tibetan yak (Bos grunniens) for ensiling Pennisetum sinese. Bioresour. Technol. 2018, 257, 76–83. [Google Scholar] [CrossRef]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- Ávila, C.L.S.; Carvalho, B.F. Silage fermentation—Updates focusing on the performance of micro-organisms. J. Appl. Microbiol. 2020, 128, 966–984. [Google Scholar] [CrossRef] [Green Version]
- Cao, C.; Bao, W.; Li, W.; Zhao, F.; Kwok, L.-Y.; Zhang, W.; Zhang, H. Changes in physico-chemical characteristics and viable bacterial communities during fermentation of alfalfa silages inoculated with Lactobacillus plantarum. World J. Microbiol. Biotechnol. 2021, 37, 127. [Google Scholar] [CrossRef]
- Ce, L.; Lai, Y.J.; Lu, X.N.; Guo, P.T.; Luo, H.L. Effect of lactic acid bacteria inoculants on alfalfa (Medicago sativa L.) silage quality: Assessment of degradation (in situ) and gas production (in vitro). J. Integr. Agric. 2016, 15, 2834–2841. [Google Scholar] [CrossRef]
- Bumbieris, V.H.; Horst, E.H.; Guimaraes, V.A.P.; Massaro, F.L.; Moraes, G.J.; Meza, D.A.R.; Galbeiro, S. Effect of microbial inoculants on the chemical composition and aerobic stability of Tanzania guinea grass silages. S. Afr. J. Anim. Sci. 2021, 51, 81–87. [Google Scholar] [CrossRef]
- Miguel, M.; Mamuad, L.; Ramos, S.; Ku, M.J.; Jeong, C.D.; Kim, S.H.; Cho, Y.I.; Lee, S.S. Effects of using different roughages in the total mixed ration inoculated with or without coculture of Lactobacillus acidophilus and Bacillus subtilis on in vitro rumen fermentation and microbial population. Anim. Biosci. 2021, 34, 642–651. [Google Scholar] [CrossRef]
- Bai, J.; Xu, D.; Xie, D.; Wang, M.; Li, Z.; Guo, X. Effects of antibacterial peptide-producing Bacillus subtilis and Lactobacillus buchneri on fermentation, aerobic stability, and microbial community of alfalfa silage. Bioresour. Technol. 2020, 315, 123881. [Google Scholar] [CrossRef] [PubMed]
- Khota, W.; Pholsen, S.; Higgs, D.; Cai, Y. Natural lactic acid bacteria population of tropical grasses and their fermentation factor analysis of silage prepared with cellulase and inoculant. J. Dairy Sci. 2016, 99, 9768–9781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.X.; Ni, K.K.; Zhang, Y.C.; Lin, Y.L.; Yang, F.Y. Influence of lactic acid bacteria, cellulase, cellulase-producing Bacillus pumilus and their combinations on alfalfa silage quality. J. Integr. Agric. 2018, 17, 2768–2782. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.S.; Wang, Y.P.; Yang, F.Y.; Wang, Y.; Zhang, H. Screening a Lactobacillus plantarum strain for good adaption in alfalfa ensiling and demonstrating its improvement of alfalfa silage quality. J. Appl. Microbiol. 2020, 129, 233–242. [Google Scholar] [CrossRef]
- Rajabi, R.; Tahmasbi, R.; Dayani, O.; Khezri, A. Chemical composition of alfalfa silage with waste date and its feeding effect on ruminal fermentation characteristics and microbial protein synthesis in sheep. J. Anim. Physiol. Anim. Nutr. 2017, 101, 466–474. [Google Scholar] [CrossRef]
- Eikmeyer, F.G.; Köfinger, P.; Poschenel, A.; Jünemann, S.; Zakrzewski, M.; Heinl, S.; Mayrhuber, E.; Grabherr, R.; Pühler, A.; Schwab, H.; et al. Metagenome analyses reveal the influence of the inoculant Lactobacillus buchneri CD034 on the microbial community involved in grass ensiling. J. Biotechnol. 2013, 167, 334–343. [Google Scholar] [CrossRef]
- Guo, L.N.; Yao, D.D.; Li, D.X.; Lin, Y.L.; Bureenok, S.; Ni, K.K.; Yang, F.Y. Effects of Lactic Acid Bacteria Isolated from Rumen Fluid and Feces of Dairy Cows on Fermentation Quality, Microbial Community, and in vitro Digestibility of Alfalfa Silage. Front. Microbiol. 2020, 10, 2998. [Google Scholar] [CrossRef]
- Yang, F.Y.; Zhao, S.S.; Wang, Y.; Fan, X.M.; Wang, Y.P.; Feng, C.S. Assessment of Bacterial Community Composition and Dynamics in Alfalfa Silages with and Without Lactobacillus plantarum Inoculation Using Absolute Quantification 16S rRNA Sequencing. Front. Microbiol. 2021, 11, 629894. [Google Scholar] [CrossRef]
- Johnson, H.E.; Merry, R.J.; Davies, D.R.; Kell, D.B.; Theodorou, M.K.; Griffith, G.W. Vacuum packing: A model system for laboratory–scale silage fermentations. J. Appl. Microbiol. 2005, 98, 106–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoedtke, S.; Zeyner, A. Comparative evaluation of laboratory-scale silages using standard glass jar silages or vacuum-packed model silages. J. Sci. Food Agric. 2011, 91, 841–849. [Google Scholar] [CrossRef]
- Broderick, G.; Kang, J.H. Automated Simultaneous Determination of Ammonia and Total Amino Acids in Ruminal Fluid and In Vitro Media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Thiex, N.; Novotny, L.; Crawford, A. Determination of Ash in Animal Feed: AOAC Official Method 942.05 Revisited. J. AOAC Int. 2012, 95, 1392–1397. [Google Scholar] [CrossRef] [PubMed]
- Arthur Thomas, T. An automated procedure for the determination of soluble carbohydrates in herbage. J. Sci. Food Agric. 1977, 28, 639–642. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Guo, J.; Xie, Y.; Yu, Z.; Meng, G.; Wu, Z. Effect of Lactobacillus plantarum expressing multifunctional glycoside hydrolases on the characteristics of alfalfa silage. Appl. Microbiol. Biotechnol. 2019, 103, 7983–7995. [Google Scholar] [CrossRef]
- Zhao, S.S.; Yang, F.Y.; Wang, Y.; Fan, X.M.; Feng, C.S.; Wang, Y.P. Dynamics of Fermentation Parameters and Bacterial Community in High-Moisture Alfalfa Silage with or without Lactic Acid Bacteria. Microorganisms 2021, 9, 1225. [Google Scholar] [CrossRef]
- Yang, F.Y.; Wang, Y.P.; Zhao, S.S.; Wang, Y. Lactobacillus plantarum Inoculants Delay Spoilage of High Moisture Alfalfa Silages by Regulating Bacterial Community Composition. Front. Microbiol. 2020, 11, 1989. [Google Scholar] [CrossRef]
- Smets, W.; Leff, J.W.; Bradford, M.A.; McCulley, R.L.; Lebeer, S.; Fierer, N. A method for simultaneous measurement of soil bacterial abundances and community composition via 16S rRNA gene sequencing. Soil Biol. Biochem. 2016, 96, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Tkacz, A.; Hortala, M.; Poole, P.S. Absolute quantitation of microbiota abundance in environmental samples. Microbiome 2018, 6, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoddard, S.F.; Smith, B.J.; Hein, R.; Roller, B.R.K.; Schmidt, T.M. rrnDB: Improved tools for interpreting rRNA gene abundance in bacteria and archaea and a new foundation for future development. Nucleic Acids Res. 2015, 43, D593–D598. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Yang, Y.; Chen, S.; Jason Shi, Z.; Zhao, M.; Zhu, Z.; Yang, S.; Qu, Y.; Ma, Q.; He, Z.; et al. Microbial functional trait of rRNA operon copy numbers increases with organic levels in anaerobic digesters. ISME J. 2017, 11, 2874–2878. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Li, X.; Guan, H.; Huang, L.; Ma, X.; Peng, Y.; Li, Z.; Nie, G.; Zhou, J.; Yang, W.; et al. Microbial community and fermentation characteristic of Italian ryegrass silage prepared with corn stover and lactic acid bacteria. Bioresour. Technol. 2019, 279, 166–173. [Google Scholar] [CrossRef]
- Tao, L.; Zhou, H.; Zhang, N.; Si, B.; Tu, Y.; Ma, T.; Diao, Q. Effects of different source additives and wilt conditions on the pH value, aerobic stability, and carbohydrate and protein fractions of alfalfa silage. Anim. Sci. J. 2017, 88, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, C.; Zhou, W.; Yang, F.-y.; Chen, X.-y.; Zhang, Q. Effects of Wilting and Lactobacillus plantarum Addition on the Fermentation Quality and Microbial Community of Moringa oleifera Leaf Silage. Front. Microbiol. 2018, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.L.; Niu, D.Z.; Jiang, D.; Zuo, S.S.; Xu, C.C. Dynamics of microbial community during ensiling direct-cut alfalfa with and without LAB inoculant and sugar. J. Appl. Microbiol. 2017, 122, 1456–1470. [Google Scholar] [CrossRef]
- Kung, L.; Shaver, R. Interpretation and use of silage fermentation analysis reports. Focus Forage 2001, 3, 1–5. [Google Scholar]
- Dong, M.; Li, Q.; Xu, F.; Wang, S.; Chen, J.; Li, W. Effects of microbial inoculants on the fermentation characteristics and microbial communities of sweet sorghum bagasse silage. Sci. Rep. 2020, 10, 837. [Google Scholar] [CrossRef] [Green Version]
- Dunière, L.; Sindou, J.; Chaucheyras-Durand, F.; Chevallier, I.; Sergentet, D. Silage processing and strategies to prevent persistence of undesirable microorganisms. Anim. Feed Sci. Technol. 2013, 182, 1–15. [Google Scholar] [CrossRef]
- Cai, Y.; Benno, Y.; Ogawa, M.; Ohmomo, S.; Kumai, S.; Nakase, T. Influence of Lactobacillus spp. from an Inoculant and of Weissella and Leuconostoc spp. from Forage Crops on Silage Fermentation. Appl. Environ. Microbiol. 1998, 64, 2982–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, T.; Ohba, N.; Shimojo, M.; Masuda, Y. Changes in Mono-and Disaccharides Compositions of Guineagrass (Panicum maximum Jacq.) Silage During Early Stages of Ensiling. J. Fac. Agric. Kyushu Univ. 2003, 47, 333–339. [Google Scholar] [CrossRef]
- Liu, Q.H.; Li, J.F.; Zhao, J.; Wu, J.X.; Shao, T. Enhancement of lignocellulosic degradation in high-moisture alfalfa via anaerobic bioprocess of engineered Lactococcus lactis with the function of secreting cellulase. Biotechnol. Biofuels 2019, 12, 88. [Google Scholar] [CrossRef] [Green Version]
- Ogunade, I.M.; Jiang, Y.; Pech Cervantes, A.A.; Kim, D.H.; Oliveira, A.S.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Bacterial diversity and composition of alfalfa silage as analyzed by Illumina MiSeq sequencing: Effects of Escherichia coli O157:H7 and silage additives. J. Dairy Sci. 2018, 101, 2048–2059. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela Saldinger, S. Bacterial Dynamics of Wheat Silage. Front. Microbiol. 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.; Wang, C.; He, L.; Zhou, W.; Yang, F.; Zhang, Q. The bacterial community and fermentation quality of mulberry (Morus alba) leaf silage with or without Lactobacillus casei and sucrose. Bioresour. Technol. 2019, 293, 122059. [Google Scholar] [CrossRef]
- Guo, L.N.; Wang, X.; Lin, Y.; Yang, X.; Ni, K.; Yang, F. Microorganisms that are critical for the fermentation quality of paper mulberry silage. Food Energy Secur. 2021, 10, e304. [Google Scholar] [CrossRef]
- Jiang, S.-Q.; Yu, Y.-N.; Gao, R.-W.; Wang, H.; Zhang, J.; Li, R.; Long, X.-H.; Shen, Q.-R.; Chen, W.; Cai, F. High-throughput absolute quantification sequencing reveals the effect of different fertilizer applications on bacterial community in a tomato cultivated coastal saline soil. Sci. Total. Environ. 2019, 687, 601–609. [Google Scholar] [CrossRef]
- Ni, K.K.; Wang, F.F.; Zhu, B.G.; Yang, J.X.; Zhou, G.A.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef]
- Polley, H.W.; Wilsey, B.J.; Derner, J.D. Dominant species constrain effects of species diversity on temporal variability in biomass production of tallgrass prairie. Oikos 2007, 116, 2044–2052. [Google Scholar] [CrossRef] [Green Version]
- Méndez-García, C.; Peláez, A.I.; Mesa, V.; Sánchez, J.; Golyshina, O.V.; Ferrer, M. Microbial diversity and metabolic networks in acid mine drainage habitats. Front. Microbiol. 2015, 6, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ni, K.K.; Minh, T.T.; Tu, T.T.M.; Tsuruta, T.; Pang, H.; Nishino, N. Comparative microbiota assessment of wilted Italian ryegrass, whole crop corn, and wilted alfalfa silage using denaturing gradient gel electrophoresis and next-generation sequencing. Appl. Microbiol. Biotechnol. 2017, 101, 1385–1394. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.K.; Zhao, J.Y.; Zhu, B.G.; Su, R.N.; Pan, Y.; Ma, J.K.; Zhou, G.A.; Tao, Y.; Liu, X.R.; Zhong, J. Assessing the fermentation quality and microbial community of the mixed silage of forage soybean with crop corn or sorghum. Bioresour. Technol. 2018, 265, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wang, Y.; Zhao, S.; Feng, C.; Fan, X. Dynamics of the Fermentation Products, Residual Non-structural Carbohydrates, and Bacterial Communities of Wilted and Non-wilted Alfalfa Silage with and Without Lactobacillus plantarum Inoculation. Front. Microbiol. 2022, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhao, J.; Dong, Z.; Li, J.; Kaka, N.A.; Shao, T. Sequencing and microbiota transplantation to determine the role of microbiota on the fermentation type of oat silage. Bioresour. Technol. 2020, 309, 123371. [Google Scholar] [CrossRef]
- Yang, L.; Yuan, X.; Li, J.; Dong, Z.; Shao, T. Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculant. Bioresour. Technol. 2019, 275, 280–287. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Li, D.X.; Wang, X.K.; Lin, Y.L.; Zhang, Q.; Chen, X.Y.; Yang, F.Y. Fermentation dynamics and diversity of bacterial community in four typical woody forages. Ann. Microbiol. 2019, 69, 233–240. [Google Scholar] [CrossRef]
- Miranda-Tello, E.; Fardeau, M.-L.; Sepúlveda, J.; Fernández, L.; Cayol, J.-L.; Thomas, P.; Ollivier, B. Garciella nitratireducens gen. nov., sp. nov., an anaerobic, thermophilic, nitrate- and thiosulfate-reducing bacterium isolated from an oilfield separator in the Gulf of Mexico. Int. J. Syst. Evol. Microbiol. 2003, 53, 1509–1514. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Alfalfa |
|---|---|
| pH | 6.52 |
| Dry matter (%) | 39.20 |
| Neutral detergent fiber (g/kg DM) | 389.79 |
| Acid detergent fiber (g/kg DM) | 274.98 |
| Water soluble carbohydrate (g/kg DM) | 76.11 |
| Rhamnse (mg/kg DM) | 69.70 |
| Galactose (mg/kg DM) | 1152.59 |
| Glucose (mg/kg DM) | 9144.42 |
| Fructose (mg/kg DM) | 14,019.40 |
| Genus | Treatments | RQS (%) | AQS (Copies/ng DNA) | ||
|---|---|---|---|---|---|
| Ensiling Days (d) | |||||
| 10 | 60 | 10 | 60 | ||
| Lactobacillus | CK | 14.51 aA | 21.76 aB | 1.0 × 108 a | 1.14 × 108 a |
| CB | 32.42 bA | 46.37 bB | 2.01 × 108 bA | 2.71 × 108 Bb | |
| ZZU203 | 96.88 c | 98.27 c | 5.04 × 108 cB | 2.77 × 108 bA | |
| ZZU203_CB | 97.18 c | 98.31 c | 5.76 × 108 dB | 2.78 × 108 bA | |
| Pediococcus | CK | 18.85 bA | 56.38 cB | 1.48 × 108 cA | 3.24 × 108 cB |
| CB | 17.85 bA | 39.63 bB | 1.12 × 108 bA | 2.26 × 108 bB | |
| ZZU203 | 1.26 a | <1.00 a | 3.35 × 106 aB | 5.93 × 105 aA | |
| ZZU203_CB | <1.00 a | <1.00 a | 3.34 × 106 a | 1.05 × 106 a | |
| Hafnia | CK | 23.08 bB | <1.00 A | 1.56 × 108 cB | 2.18 × 105 bA |
| CB | 18.44 bB | <1.00 A | 1.12 × 108 bB | 3.25 × 105 bA | |
| ZZU203 | <1.00 a | <1.00 | 7.53 × 105 a | 8.96 × 104 a | |
| ZZU203_CB | <1.00 a | <1.00 | 1.90 × 106 a | 7.47 × 104 a | |
| Enterococcus | CK | 13.48 cB | 8.95 bA | 9.50 × 107 cB | 4.72 × 107 bA |
| CB | 5.79 b | 6.12 b | 3.21 × 107 b | 3.29 × 107 b | |
| ZZU203 | <1.00 a | <1.00 a | 1.16 × 105 a | 6.79 × 104 a | |
| ZZU203_CB | <1.00 a | <1.00 a | 1.17 × 105 a | 5.32 × 104 a | |
| Weissella | CK | 9.08 c | 9.13 c | 6.12 × 107 c | 4.83 × 107 c |
| CB | 5.28 b | 4.85 b | 2.92 × 107 b | 2.43 × 107 b | |
| ZZU203 | <1.00 a | <1.00 a | 1.31 × 105 a | 7.15 × 104 a | |
| ZZU203_CB | <1.00 a | <1.00 a | 1.64 × 105 a | 5.65 × 104 a | |
| Lactococcus | CK | 1.2 bB | <1.00 A | 7.43 × 106 cB | 1.41 × 106 bA |
| CB | <1.00 a | <1.00 | 3.31 × 106 b | 4.26 × 105 b | |
| ZZU203 | <1.00 a | <1.00 | 1.28 × 104 a | 1.59 × 103 a | |
| ZZU203_CB | <1.00 a | <1.00 | 5.36 × 104 a | 9.90 × 102 a | |
| Garciella | CK | <1.00 A | 1.55 bB | 5.36 × 104 cA | 8.63 × 106 bB |
| CB | <1.00 | <1.00 a | 1.63 × 104 b | 3.16 × 104 a | |
| ZZU203 | <1.00 | <1.00 a | 1.91 × 104 b | 7.56 × 103 a | |
| ZZU203_CB | <1.00 | <1.00 a | 7.63 × 103 a | 7.34 × 103 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhao, S.; Wang, Y.; Yang, F.; Wang, Y.; Fan, X.; Feng, C. The Effect of Lactiplantibacillus plantarum ZZU203, Cellulase-Producing Bacillus methylotrophicus, and Their Combinations on Alfalfa Silage Quality and Bacterial Community. Fermentation 2023, 9, 287. https://doi.org/10.3390/fermentation9030287
Zhang X, Zhao S, Wang Y, Yang F, Wang Y, Fan X, Feng C. The Effect of Lactiplantibacillus plantarum ZZU203, Cellulase-Producing Bacillus methylotrophicus, and Their Combinations on Alfalfa Silage Quality and Bacterial Community. Fermentation. 2023; 9(3):287. https://doi.org/10.3390/fermentation9030287
Chicago/Turabian StyleZhang, Xueying, Shanshan Zhao, Yanping Wang, Fengyuan Yang, Yuan Wang, Xiaomiao Fan, and Changsong Feng. 2023. "The Effect of Lactiplantibacillus plantarum ZZU203, Cellulase-Producing Bacillus methylotrophicus, and Their Combinations on Alfalfa Silage Quality and Bacterial Community" Fermentation 9, no. 3: 287. https://doi.org/10.3390/fermentation9030287
APA StyleZhang, X., Zhao, S., Wang, Y., Yang, F., Wang, Y., Fan, X., & Feng, C. (2023). The Effect of Lactiplantibacillus plantarum ZZU203, Cellulase-Producing Bacillus methylotrophicus, and Their Combinations on Alfalfa Silage Quality and Bacterial Community. Fermentation, 9(3), 287. https://doi.org/10.3390/fermentation9030287