
Metabolome Analysis of the Effects of Sake Lees on Adipocyte Differentiation and Lipid Accumulation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Oil Red O Stain
2.4. Metabolome Analysis
2.5. Analysis of Free Amino Acids in Cell Culture Medium by High-Performance Liquid Chromatography
2.6. Statistical Analysis
3. Results
3.1. Lipid Accumulation
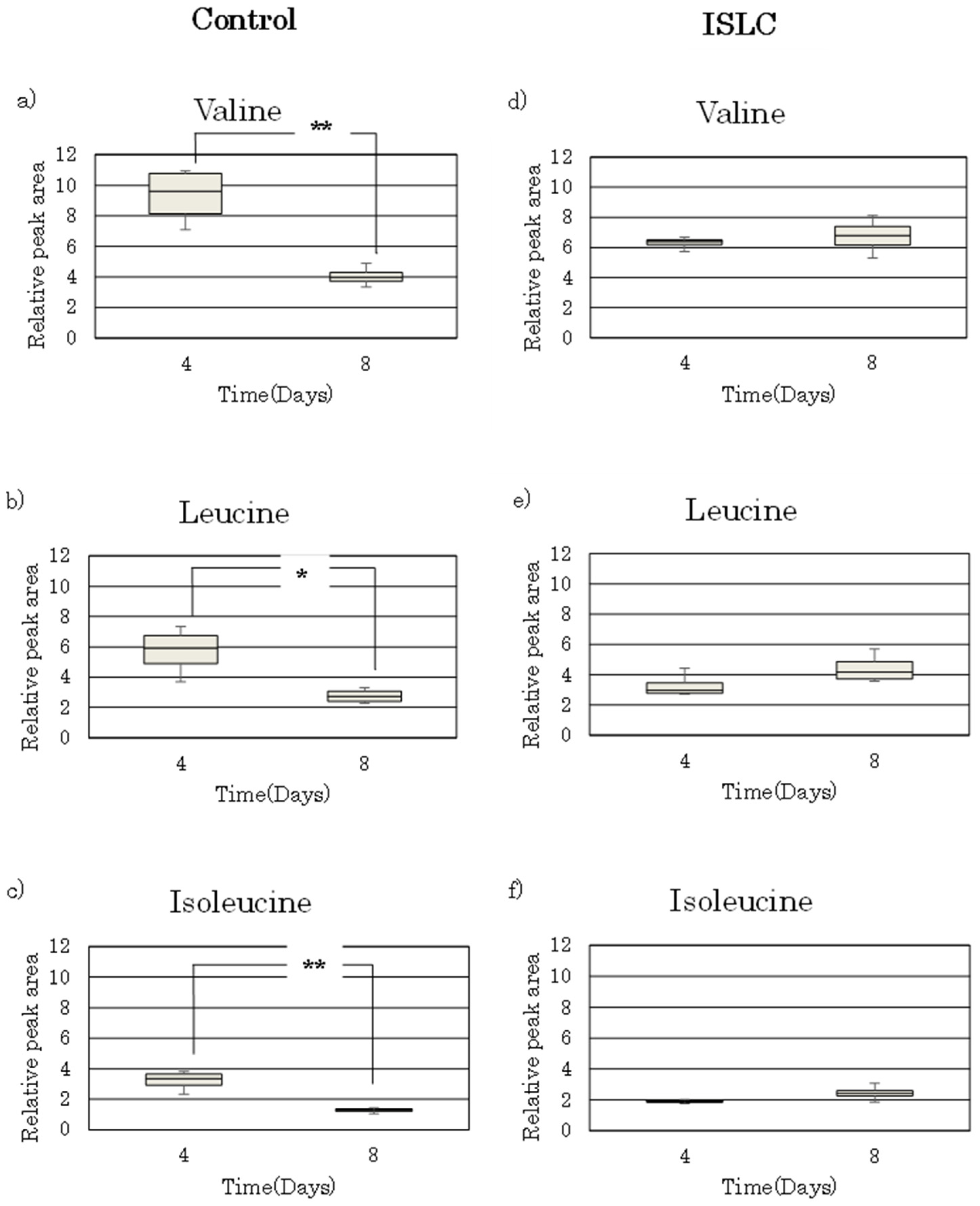
3.2. Metabolome Analysis
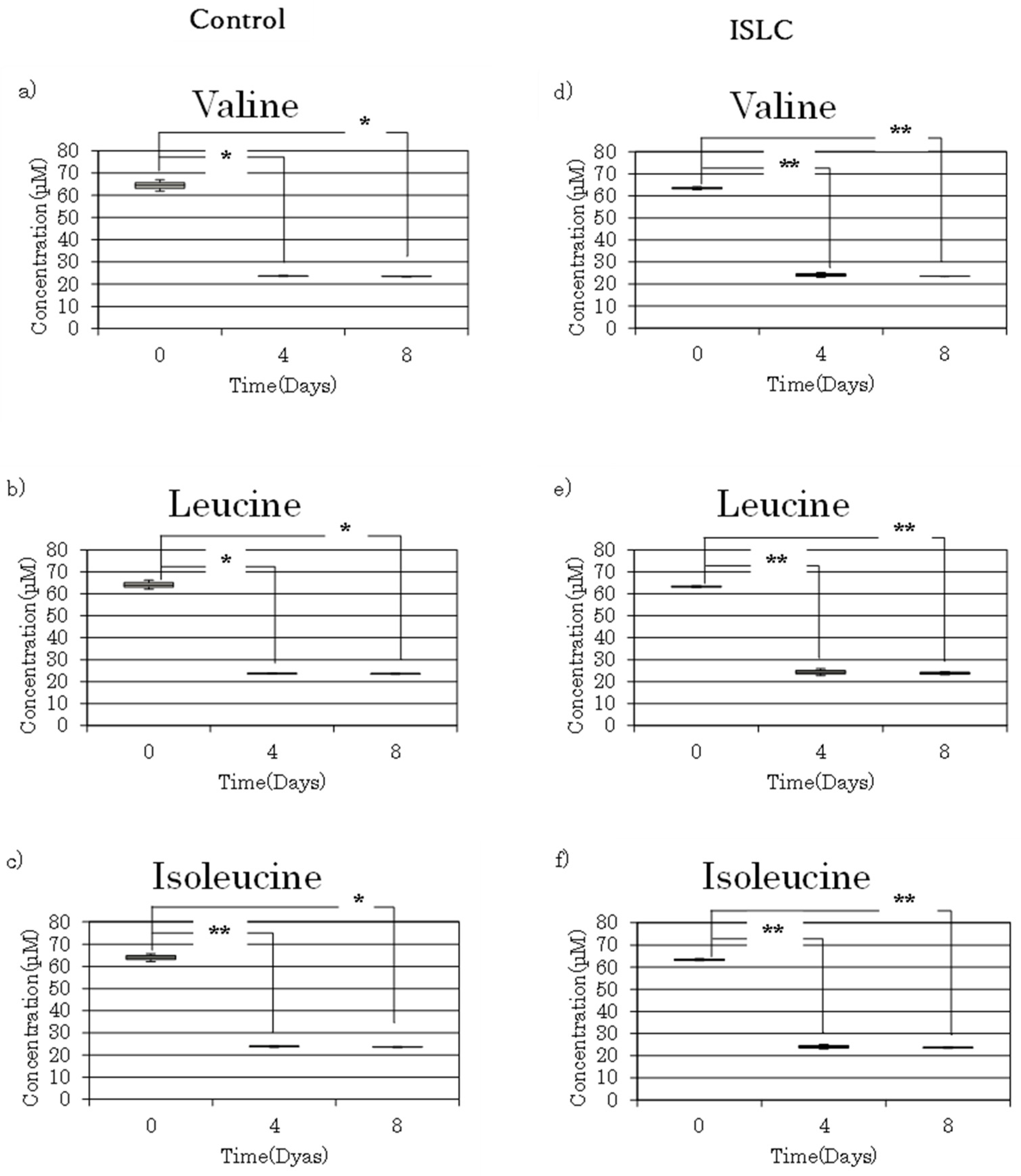
3.3. Analysis of BCAAs in Cell Culture Medium
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ministry of Health, Labor and Welfare. Available online: https://www.mhlw.go.jp/toukei/saikin/hw/life/life21/dl/life18-15.pdf. (accessed on 1 February 2023).
- Ministry of Agriculture, Forestry and Fisheries. Available online: https://www.maff.go.jp/j/wpaper/w_maff/r2/r2_h/trend/part1/pdf/c1_2_00.pdf (accessed on 1 February 2022).
- Ministry of Agriculture, Forestry and Fisheries. Available online: https://www.e-stat.go.jp/stat-search/files?page=1&layout=datalist&toukei=00500300&tstat=000001017950&cycle=8&year=20191&month=0&tclass1=000001032890&tclass2=000001151387 (accessed on 1 February 2022).
- Rodgers, R.J.; Tschöp, M.H.; Wilding, J.P. Anti-obesity drugs: Past, present and future. Dis. Models Mech. 2012, 5, 621–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barengolts, E.; Smith, E.D.; Reutrakul, S.; Tonucci, L.; Anothaisintawee, T. The Effect of Probiotic Yogurt on Glycemic Control in Type 2 Diabetes or Obesity: A Meta-Analysis of Nine Randomized Controlled Trials. Nutrients 2019, 11, 671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, S.; Fernández-Navarro, T.; Arboleya, S.; de Los Reyes-Gavilán, C.G.; Salazar, N.; Gueimonde, M. Fermented Dairy Foods: Impact on Intestinal Microbiota and Health-Linked Biomarkers. Front. Microbiol. 2019, 10, 1046. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Wanezaki, K.; Kawato, A.; Imayasu, S. Structure and activity of angiotensin I converting enzyme inhibitory peptides from sake and sake lees. Biosci. Biotechnol. Biochem. 1994, 58, 1767–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izu, H.; Shibata, S.; Fujii, T.; Matsubara, K. Sake cake (sake-kasu) ingestion increases branched-chain amino acids in the plasma, muscles, and brains of senescence-accelerated mice prone 8. Biosci. Biotechnol. Biochem. 2019, 83, 1490–1497. [Google Scholar] [CrossRef] [PubMed]
- Kubo, H.; Hoshi, M.; Matsumoto, T.; Irie, M.; Oura, S.; Tsutsumi, H.; Hata, Y.; Yamamoto, Y.; Saito, K. Sake lees extract improves hepatic lipid accumulation in high fat diet-fed mice. Lipids Health Dis. 2017, 16, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, K.; Moritani, C.; Uraji, M.; Fujita, A.; Kawakami, K.; Hatanaka, T.; Suzaki, E.; Tsuboi, S. Sake lees hydrolysate protects against acetaminophen-induced hepatotoxicity via activation of the Nrf2 antioxidant pathway. J. Clin. Biochem. Nutr. 2017, 61, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Li, E.-M.; Xu, L.-Y. Guide to Metabolomics Analysis: A Bioinformatics Workflow. Metabolites 2022, 12, 357. [Google Scholar] [CrossRef] [PubMed]
- Yoshie, T.; Nishiumi, S.; Izumi, Y.; Sakai, A.; Inoue, J.; Azuma, T.; Yoshida, M. Regulation of the metabolite profile by an APC gene mutation in colorectal cancer. Cancer Sci. 2012, 103, 1010–1021. [Google Scholar] [CrossRef] [PubMed]
- Halama, A.; Horsch, M.; Kastenmuller, G.; Moller, G.; PKumar Prehn, C.; Laumen, H.; Hauner, H.; Hrabe de Angelis, M.; Beckers, J.; Suhre, K.; et al. Metabolic switch during adipogenesis: From branched chain amino acid catabolism to lipid synthesis. Arch. Biochem. Biophys. 2016, 589, 93–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Näslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A Branched-Chain Amino Acid-Related Metabolic Signature that Differentiates Obese and Lean Humans and Contributes to Insulin Resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihalik, S.J.; Michaliszyn, S.F.; de las Heras, J.; Bacha, F.; Lee, S.; Chace, D.H.; DeJesus, V.R.; Vockley, J.; Arslanian, S.A. Metabolomic Profiling of Fatty Acid and Amino Acid Metabolism in Youth with Obesity and Type 2 Diabetes. Diabetes Care 2012, 35, 605–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Guo, K.; LeBlanc, R.E.; Loh, D.; Schwartz, G.J.; Yu, Y.H. Increasing Dietary Leucine Intake Reduces Diet-Induced. Diabetes 2007, 56, 1647–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junko, N.; Takayuki, M.; Mie, A.; Masataka, S.; Hironobu, Y. Isoleucine Prevents the Accumulation of Tissue Triglycerides and Upregulates the Expression of PPARα and Uncoupling Protein in Diet-Induced Obese Mice1-3. J. Nutr. 2010, 140, 496–500. [Google Scholar]
- Ma, Q.; Zhou, X.; Sun, Y.; Hu, L.; Zhu, J.; Shao, C.; Meng, Q.; Shan, A. Threonine, but Not Lysine and Methionine, Reduces Fat Accumulation by Regulating Lipid Metabolism in Obese Mice. J. Agric. Food Chem. 2020, 68, 4876–4883. [Google Scholar] [CrossRef] [PubMed]
- Opara, E.C.; Petro, A.; Tevrizian, A.; Feinglos, M.N.; Surwit, R.S. L-Glutamine Supplementation of a High Fat Diet Reduces Body Weight and Attenuates Hyperglycemia and Hyperinsulinemia in C57BL/6J Mice. J. Nutr. 1996, 126, 273–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Composition | Content (g/100g) |
|---|---|
| Protein | 57.4 |
| Fat | 5.4 |
| Carbohydrate | 29.6 |
| Moisture | 6.5 |
| Ash | 1.1 |
| Dietary fiber | 25.7 |
| Metabolites | Day 4 | Day 8 | ||
|---|---|---|---|---|
| p Value | Fold Induction | p Value | Fold Induction | |
| Pyruvic acid | 0.0055 | 0.76 | 0.3270 | 1.11 |
| Lactic acid | 0.0002 | 0.67 | 0.1753 | 1.19 |
| Glycolic acid | 0.2290 | 0.91 | 0.0019 | 1.38 |
| Alanine | 0.1153 | 0.63 | 0.0358 | 3.06 |
| 2-keto-isovaleric acid | 0.8862 | 0.96 | 0.0436 | 2.81 |
| Glycine | 0.8704 | 1.07 | 0.0371 | 2.99 |
| 2-Hydroxybutyric acid | 0.2360 | 0.84 | 0.0003 | 1.84 |
| Sarcosine | 0.4409 | 1.09 | 0.0076 | 1.54 |
| 2-Aminoisobutyric acid | 0.0167 | 1.21 | 0.00002 | 1.51 |
| 3-Hydroxybutyric acid | 0.0025 | 0.69 | 0.0507 | 1.36 |
| 3-Aminopropanoic acid | 0.0080 | 0.26 | 0.1166 | 2.78 |
| Acetoacetic acid | 0.1156 | 0.80 | 0.0169 | 1.69 |
| 3-Hydroxyisovaleric acid | 0.1585 | 0.91 | 0.0152 | 1.56 |
| Valine | 0.0192 | 0.68 | 0.0070 | 1.67 |
| Dihydroxyacetone | 0.0024 | 0.56 | 0.1078 | 1.47 |
| 2-Aminoethanol | 0.1878 | 0.87 | 0.0003 | 1.79 |
| Glycerol | 0.0036 | 0.84 | 0.0021 | 1.38 |
| Leucine | 0.0327 | 0.57 | 0.0225 | 1.60 |
| Octanoic acid | 0.0417 | 1.29 | 0.1766 | 1.32 |
| Phosphoric acid | 0.9007 | 1.01 | 0.0067 | 1.64 |
| Isoleucine | 0.0075 | 0.59 | 0.0043 | 1.92 |
| Proline | 0.0151 | 0.52 | 0.0080 | 2.54 |
| Succinic acid | 0.0011 | 0.56 | 0.0452 | 1.53 |
| Glyceric acid | 0.0019 | 0.58 | 0.5820 | 1.13 |
| Uracil | 0.0023 | 0.73 | 0.0786 | 1.29 |
| Serine | 0.0129 | 0.41 | 0.0022 | 2.39 |
| Fumaric acid | 0.5168 | 0.95 | 0.0037 | 1.65 |
| Homoserine | 0.1694 | 1.14 | 0.0272 | 1.95 |
| Nonanoic acid | 0.0633 | 1.14 | 0.0063 | 1.47 |
| Threonine | 0.0232 | 0.63 | 0.0010 | 2.73 |
| Glutaric acid | 0.0992 | 0.77 | 0.0347 | 1.74 |
| Citramalic acid | 0.1531 | 0.89 | 0.0114 | 2.03 |
| Threitol | 0.0383 | 0.79 | 0.0498 | 1.83 |
| Malic acid | 0.0428 | 0.78 | 0.0492 | 1.83 |
| meso-Erythritol | 0.0023 | 0.81 | 0.0374 | 1.84 |
| Aspartic acid | 0.0231 | 0.56 | 0.0001 | 2.04 |
| 3-Aminoglutaric acid | 0.0252 | 0.56 | 0.0001 | 2.03 |
| 4-Hydroxyproline | 0.0102 | 0.61 | 0.0128 | 2.51 |
| Methionine | 0.0153 | 0.59 | 0.0027 | 2.31 |
| 4-Aminobutyric acid | 0.0670 | 1.15 | 0.0056 | 2.75 |
| 5-Oxoproline | 0.1823 | 0.88 | 0.0206 | 2.30 |
| Cysteine | 0.0100 | 0.53 | 0.0011 | 2.18 |
| Creatinine | 0.1297 | 0.78 | 0.0026 | 1.61 |
| 2-ketoglutaric acid | 0.7964 | 1.04 | 0.0071 | 1.99 |
| Glutamic acid | 0.0148 | 0.55 | 0.0041 | 1.75 |
| 5-Aminovaleric acid | 0.3171 | 1.18 | 0.0010 | 2.03 |
| Xylose | 0.0136 | 0.85 | 0.0268 | 2.05 |
| Phenylalanine | 0.0030 | 0.63 | 0.0077 | 2.33 |
| Lyxose | 0.8255 | 0.98 | 0.0004 | 1.46 |
| Arabinose | 0.0074 | 0.80 | 0.0164 | 1.84 |
| Ribulose | 0.6849 | 0.93 | 0.0118 | 1.39 |
| Lauric acid | 0.0909 | 1.08 | 0.0062 | 1.41 |
| Ribose | 0.5429 | 1.05 | 0.0007 | 1.48 |
| Homocysteine | 0.0041 | 0.63 | 0.0285 | 2.02 |
| Asparagine | 0.0864 | 0.65 | 0.0045 | 2.04 |
| Xylitol | 0.0292 | 0.72 | 0.0095 | 1.85 |
| Arabitol | 0.2740 | 1.04 | 0.0001 | 1.42 |
| 2-Deoxy-glucose | 0.5964 | 1.03 | 0.00004 | 1.42 |
| Orotic acid | 0.0001 | 0.72 | 0.0005 | 1.74 |
| Isocitric acid | 0.0440 | 1.40 | 0.0623 | 2.95 |
| 2-Aminopimelic acid | 0.0256 | 1.50 | 0.0350 | 3.09 |
| Citric acid | 0.0306 | 1.50 | 0.0343 | 3.16 |
| Ornithine | 0.7128 | 0.88 | 0.0249 | 1.84 |
| Glycyl-Glycine | 0.1939 | 0.90 | 0.0045 | 1.54 |
| Cadaverine | 0.0463 | 0.84 | 0.0039 | 1.47 |
| Mannose | 0.0048 | 0.73 | 0.0174 | 1.78 |
| Glucose | 0.0015 | 0.47 | 0.3026 | 2.10 |
| Galacturonic acid | 0.1774 | 0.87 | 0.0058 | 1.84 |
| Glucuronic acid | 0.1764 | 0.87 | 0.0069 | 1.84 |
| Tryptamine | 0.0584 | 0.82 | 0.0219 | 1.85 |
| Glucaric acid | 0.0272 | 0.72 | 0.0077 | 1.71 |
| Palmitoleic acid | 0.0024 | 2.13 | 0.0508 | 1.41 |
| Inositol | 0.2936 | 0.91 | 0.0014 | 1.43 |
| Margaric acid | 0.0337 | 1.14 | 0.0023 | 1.38 |
| Kynurenine | 0.0583 | 0.73 | 0.0002 | 1.54 |
| 2,3-bisphosphoglyceric acid | 0.0936 | 0.77 | 0.0005 | 1.58 |
| Cystamine | 0.0408 | 0.78 | 0.0091 | 1.43 |
| Elaidic acid | 0.9684 | 0.98 | 0.0328 | 1.29 |
| Stearic acid | 0.9072 | 1.01 | 0.0042 | 1.37 |
| Maltose | 0.0141 | 0.78 | 0.1247 | 1.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Motono, Y.; Nishiumi, S.; Yoshida, M.; Takaoka, M. Metabolome Analysis of the Effects of Sake Lees on Adipocyte Differentiation and Lipid Accumulation. Fermentation 2023, 9, 300. https://doi.org/10.3390/fermentation9030300
Motono Y, Nishiumi S, Yoshida M, Takaoka M. Metabolome Analysis of the Effects of Sake Lees on Adipocyte Differentiation and Lipid Accumulation. Fermentation. 2023; 9(3):300. https://doi.org/10.3390/fermentation9030300
Chicago/Turabian StyleMotono, Yuki, Shin Nishiumi, Masaru Yoshida, and Motoko Takaoka. 2023. "Metabolome Analysis of the Effects of Sake Lees on Adipocyte Differentiation and Lipid Accumulation" Fermentation 9, no. 3: 300. https://doi.org/10.3390/fermentation9030300
APA StyleMotono, Y., Nishiumi, S., Yoshida, M., & Takaoka, M. (2023). Metabolome Analysis of the Effects of Sake Lees on Adipocyte Differentiation and Lipid Accumulation. Fermentation, 9(3), 300. https://doi.org/10.3390/fermentation9030300