
Metschnikowia pulcherrima in Cold Clarification: Biocontrol Activity and Aroma Enhancement in Verdicchio Wine



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fermentation Trials at the Winery Level
2.1.1. M. pulcherrima Biomass Production
2.1.2. Preparation of Pied de Cuve
2.1.3. Inoculation at Cold Clarification Stage and Fermentation
2.2. Biomass Evolution
2.3. Saccharomyces cerevisiae Typing
2.4. Analytical Procedures
2.5. Sensory Analysis
2.6. Data Analyses
3. Results
3.1. Effect of M. pulcherrima Addition in Cold Clarification
3.2. Pied de Cuve Inoculum and S. cerevisiae Strains Typing
3.3. Biomass Evolution and Saccharomyces cerevisiae Characterization
3.4. Main Oenological Characters of Wine
3.5. Volatile Compounds of Wine
3.6. Sensory Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Escribano-Viana, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine aroma evolution throughout alcoholic fermentation sequentially inoculated with non-Saccharomyces/Saccharomyces yeasts. Food Res. Int. 2018, 112, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Roullier-Gall, C.; Hemmler, D.; Gonsior, M.; Li, Y.; Nikolantonaki, M.; Aron, A.; Coelho, C.; Gougeon, R.D.; Schmitt-Kopplin, P. Sulfites and the wine metabolome. Food Chem. 2017, 237, 106–113. [Google Scholar] [CrossRef]
- Ubeda, C.; Hornedo-Ortega, R.; Cerezo, A.B.; Garcia-Parrilla, M.C.; Troncoso, A.M. Chemical hazards in grapes and wine, climate change and challenges to face. Food Chem. 2020, 314, 126222. [Google Scholar] [CrossRef] [PubMed]
- Giacosa, S.; Segade, S.R.; Cagnasso, E.; Caudana, A.; Rolle, L.; Gerbi, V. SO2 in wines: Rational use and possible alternatives. In Red Wine Technology; Academic Press: Cambridge, MA, USA, 2019; pp. 309–321. [Google Scholar]
- Lisanti, M.T.; Blaiotta, G.; Nioi, C.; Moio, L. Alternative methods to SO2 for microbiological stabilization of wine. Compr. Rev. Food Sci. Food Saf. 2019, 18, 455–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chacon-Rodriguez, L.; Joseph, C.L.; Nazaris, B.; Coulon, J.; Richardson, S.; Dycus, D.A. Innovative use of non-Saccharomyces in bio-protection: T. delbrueckii and M. pulcherrima applied to a machine harvester. Catal. Discov. Into Pract. 2020, 4, 82–90. [Google Scholar] [CrossRef]
- Simonin, S.; Alexandre, H.; Nikolantonaki, M.; Coelho, C.; Tourdot-Maréchal, R. Inoculation of Torulaspora delbrueckii as a bio-protection agent in winemaking. Food Res. Int. 2018, 107, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Windholtz, S.; Redon, P.; Lacampagne, S.; Farris, L.; Lytra, G.; Cameleyre, M.; Barbe, J.C.; Coulon, J.; Masneuf-Pomarede, I. Non-Saccharomyces yeasts as bioprotection in the composition of red wine and in the reduction of sulfur dioxide. LWT 2021, 149, 111781. [Google Scholar] [CrossRef]
- Prior, K.J.; Bauer, F.F.; Divol, B. The utilisation of nitrogenous compounds by commercial non-Saccharomyces yeasts associated with wine. Food Microbiol. 2019, 79, 75–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comitini, F.; Capece, A.; Ciani, M.; Romano, P. New insights on the use of wine yeasts. Curr. Opin. Food Sci. 2017, 13, 44–49. [Google Scholar] [CrossRef]
- Comitini, F.; Agarbati, A.; Canonico, L.; Galli, E.; Ciani, M. Purification and characterization of WA18, a new mycocin produced by Wickerhamomyces anomalus active in wine against Brettanomyces bruxellensis spoilage yeasts. Microorganisms 2020, 9, 56. [Google Scholar] [CrossRef]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef]
- Di Gianvito, P.; Englezos, V.; Rantsiou, K.; Cocolin, L. Bioprotection strategies in winemaking. Int. J. Food Microbiol. 2022, 364, 109532. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; González-Arenzana, L.; Garijo, P.; Fernández, L.; López, R.; Santamaría, P.; Gutiérrez, A.R. Bioprotective effect of a Torulaspora delbrueckii/Lachancea thermotolerans-mixed inoculum in red winemaking. Fermentation 2022, 8, 337. [Google Scholar] [CrossRef]
- Simonin, S.; Roullier-Gall, C.; Ballester, J.; Schmitt-Kopplin, P.; Quintanilla-Casas, B.; Vichi, S.; Peyron, D.; Alexandre, H.; Tourdot-Maréchal, R. Bio-Protection as an Alternative to Sulphites: Impact on Chemical and Microbial Characteristics of Red Wines. Front. Microbiol. 2020, 11, 1308. [Google Scholar] [CrossRef] [PubMed]
- Pawlikowska, E.; James, S.A.; Breierova, E.; Antolak, H.; Kregiel, D. Biocontrol capability of local Metschnikowia sp. isolates. Antonie Van Leeuwenhoek 2019, 112, 1425–1445. [Google Scholar] [CrossRef] [Green Version]
- Kuchen, B.; Vazquez, F.; Maturano, Y.P.; Scaglia, G.J.E.; Pera, L.M.; Vallejo, M.D. Toward application of biocontrol to inhibit wine spoilage yeasts: The use of statistical designs for screening and optimisation. OENO One 2021, 55, 75–96. [Google Scholar] [CrossRef]
- Varela, C.; Sengler, F.; Solomon, M.; Curtin, C. Volatile flavour profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 2016, 209, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, B.; Zhang, Z.; Chen, Y.; Tian, S. Antagonistic yeasts: A promising alternative to chemical fungicides for controlling postharvest decay of fruit. J. Fungi. 2020, 6, 158. [Google Scholar] [CrossRef] [PubMed]
- Csutak, O.; Vassu, T.; Sarbu, I.; Stoica, I.; Cornea, P. Antagonistic activity of three newly isolated yeast strains from the surface of fruits. Food Technol. Biotechnol. 2013, 51, 70–77. [Google Scholar]
- Kántor, A.; Kačániová, M. Diversity of bacteria and yeasts on the surface of table grapes. Anim. Sci. Biotechnol. 2015, 48, 149–155. [Google Scholar]
- Oro, L.; Feliziani, E.; Ciani, M.; Romanazzi, G.; Comitini, F. Biocontrol of postharvest brown rot of sweet cherries by Saccharomyces cerevisiae Disva 599, Metschnikowia pulcherrima Disva 267 and Wickerhamomyces anomalus Disva 2 strains. Postharvest Biol. Technol. 2014, 96, 64–68. [Google Scholar] [CrossRef]
- Saravanakumar, D.; Ciavorella, A.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Metschnikowia pulcherrima strain MACH1 outcompetes Botrytis cinerea, Alternaria alternata and Penicillium expansum in apples through iron depletion. Postharvest Biol. Technol. 2008, 49, 121–128. [Google Scholar] [CrossRef]
- Agarbati, A.; Canonico, L.; Pecci, T.; Romanazzi, G.; Ciani, M.; Comitini, F. Biocontrol of non-Saccharomyces yeasts in vineyard against the gray mold disease agent Botrytis cinerea. Microorganisms 2022, 10, 200. [Google Scholar] [CrossRef]
- Agarbati, A.; Canonico, L.; Ciani, M.; Comitini, F. The impact of fungicide treatments on yeast biota of Verdicchio and Montepulciano grape varieties. PLoS ONE 2019, 14, e0217385. [Google Scholar] [CrossRef] [Green Version]
- Legras, J.L.; Karst, F. Optimisation of interdelta analysis for Saccharomyces cerevisiae strain characterisation. FEMS Microbiol. Lett. 2003, 221, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dukes, B.C.; Butzke, C.E. Rapid determination of primary amino acids in grape juice using an o-phthaldialdehyde/N-acetyl-L-cysteine spectrophotometric assay. Am. J. Enol. Vitic. 1998, 49, 125–134. [Google Scholar] [CrossRef]
- Canonico, L.; Comitini, F.; Ciani, M. Influence of vintage and selected starter on Torulaspora delbrueckii/Saccharomyces cerevisiae sequential fermentation. Eur. Food Res. Technol. 2015, 241, 827–833. [Google Scholar] [CrossRef]
- Canonico, L.; Ciani, E.; Galli, E.; Comitini, F.; Ciani, M. Evolution of aromatic profile of Torulaspora delbrueckii mixed fermentation at microbrewery plant. Fermentation 2020, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Ab Rahman, S.F.S.; Singh, E.; Pieterse, C.M.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Navarro, C.; González-Muñoz, M.T.; Jimenez-Lopez, C.; Rodriguez-Gallego, M. Bioprotection. In Encyclopedia of Earth Sciences Series; Finkl, C.W., Ed.; Springer: Berlin, Germany, 2011; pp. 185–189. [Google Scholar]
- Biocontrol-Dictionary Agroecology. Available online: https://dicoagroecologie.fr/en/encyclopedia/biocontrol/ (accessed on 2 February 2023).
- Morata, A.; Loira, I.; Escott, C.; del Fresno, J.M.; Bañuelos, M.A.; Suárez-Lepe, J.A. Applications of Metschnikowia pulcherrima in wine biotechnology. Fermentation 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Türkel, S.; Ener, B. Isolation and characterization of new Metschnikowia pulcherrima strains as producers of the antimicrobial pigment pulcherrimin. Z. Für Nat. C 2009, 64, 405–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zott, K.; Thibon, C.; Bely, M.; Lonvaud-Funel, A.; Dubourdieu, D.; Masneuf-Pomarede, I. The grape must non-Saccharomyces microbial community: Impact on volatile thiol release. Int. J. Food Microbiol. 2011, 151, 210–215. [Google Scholar] [CrossRef]
- Ruiz, J.; Belda, I.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Rauht, D.; Santos, A.; Benito, S. Analytical impact of Metschnikowia pulcherrima in the volatile profile of Verdejo white wines. App. Microbiol. Biotechnol. 2018, 102, 8501–8509. [Google Scholar] [CrossRef] [Green Version]
- Vicente, J.; Ruiz, J.; Belda, I.; Benito-Vázquez, I.; Marquina, D.; Calderón, F.; Santos, A.; Benito, S. The genus Metschnikowia Muñoz-in enology. Microorganisms 2020, 8, 1038. [Google Scholar] [CrossRef] [PubMed]
- Redondo, J.M.; Puertas, B.; Cantos-Villar, E.; Jiménez-Hierro, M.J.; Carbú, M.; Garrido, C.; Puiz-Moreno, M.J.; Moreno-Rojas, J.M. Impact of sequential inoculation with the non-Saccharomyces T. delbrueckii and M. pulcherrima combined with Saccharomyces cerevisiae strains on chemicals and sensory profile of rosé wines. J. Agr. Food Chem. 2021, 69, 1598–1609. [Google Scholar] [CrossRef] [PubMed]
- Duarte, F.L.; Egipto, R.; Baleiras-Couto, M.M. Mixed fermentation with Metschnikowia pulcherrima using different grape varieties. Fermentation 2019, 5, 59. [Google Scholar] [CrossRef] [Green Version]
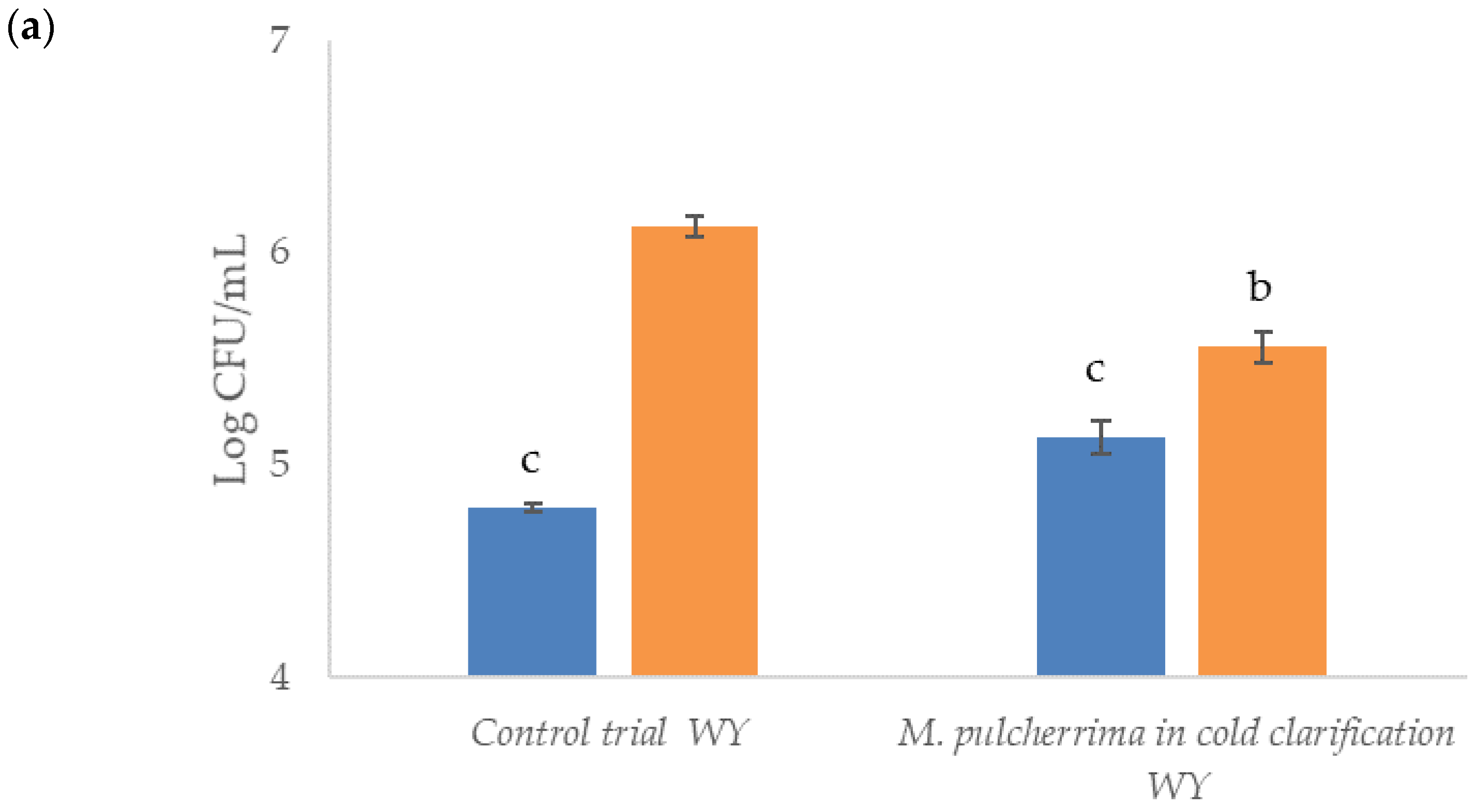
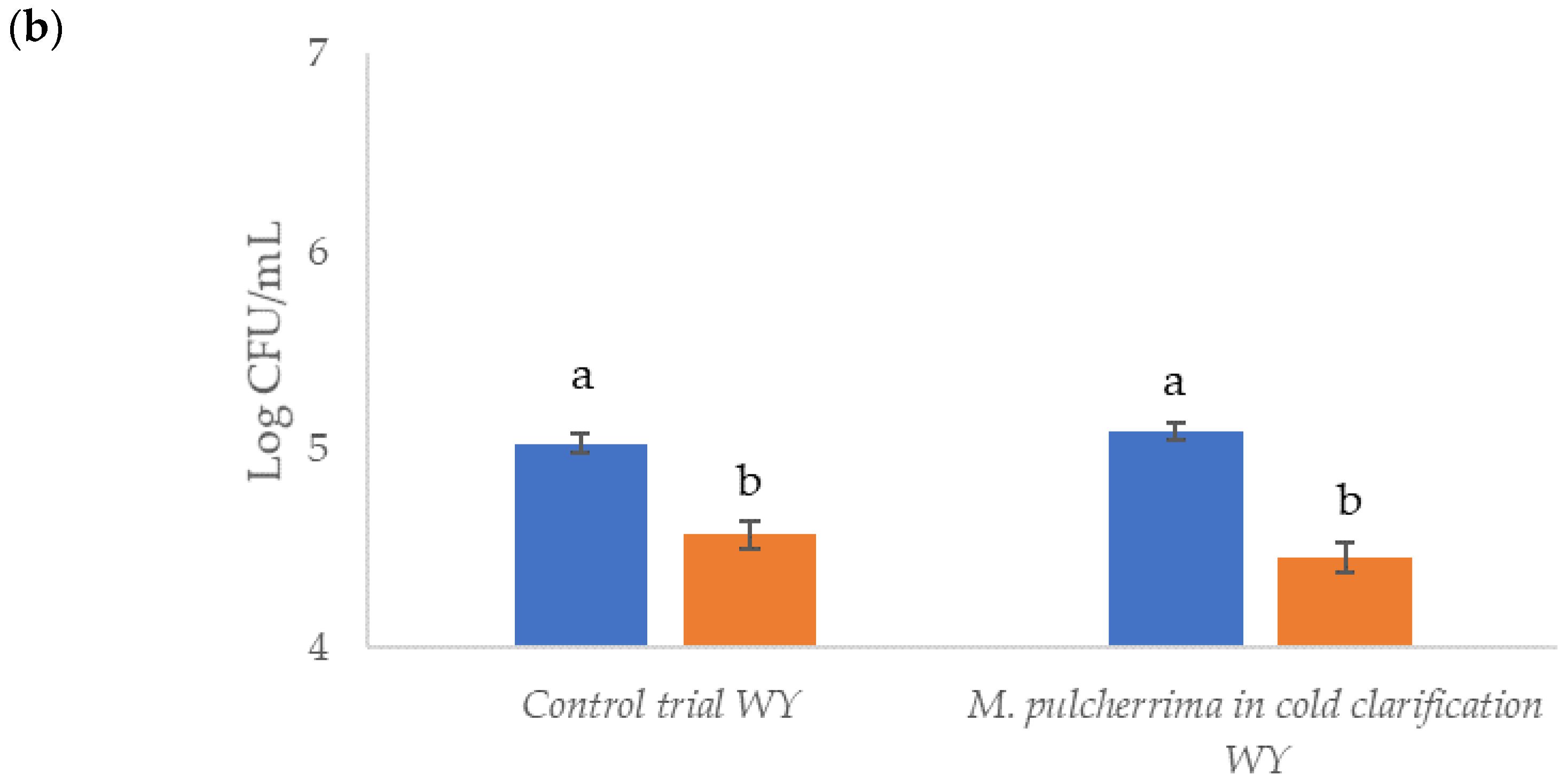
 before clarification;
before clarification;  after clarification. Data (n = 3) are the means ± standard deviation. Data with different superscript letters (a,b,c) are significantly different (Duncan tests; p < 0.05).
before clarification; after clarification. Data (n = 3) are the means ± standard deviation. Data with different superscript letters (a,b,c) are significantly different (Duncan tests; p < 0.05).
after clarification. Data (n = 3) are the means ± standard deviation. Data with different superscript letters (a,b,c) are significantly different (Duncan tests; p < 0.05).
before clarification; after clarification. Data (n = 3) are the means ± standard deviation. Data with different superscript letters (a,b,c) are significantly different (Duncan tests; p < 0.05).

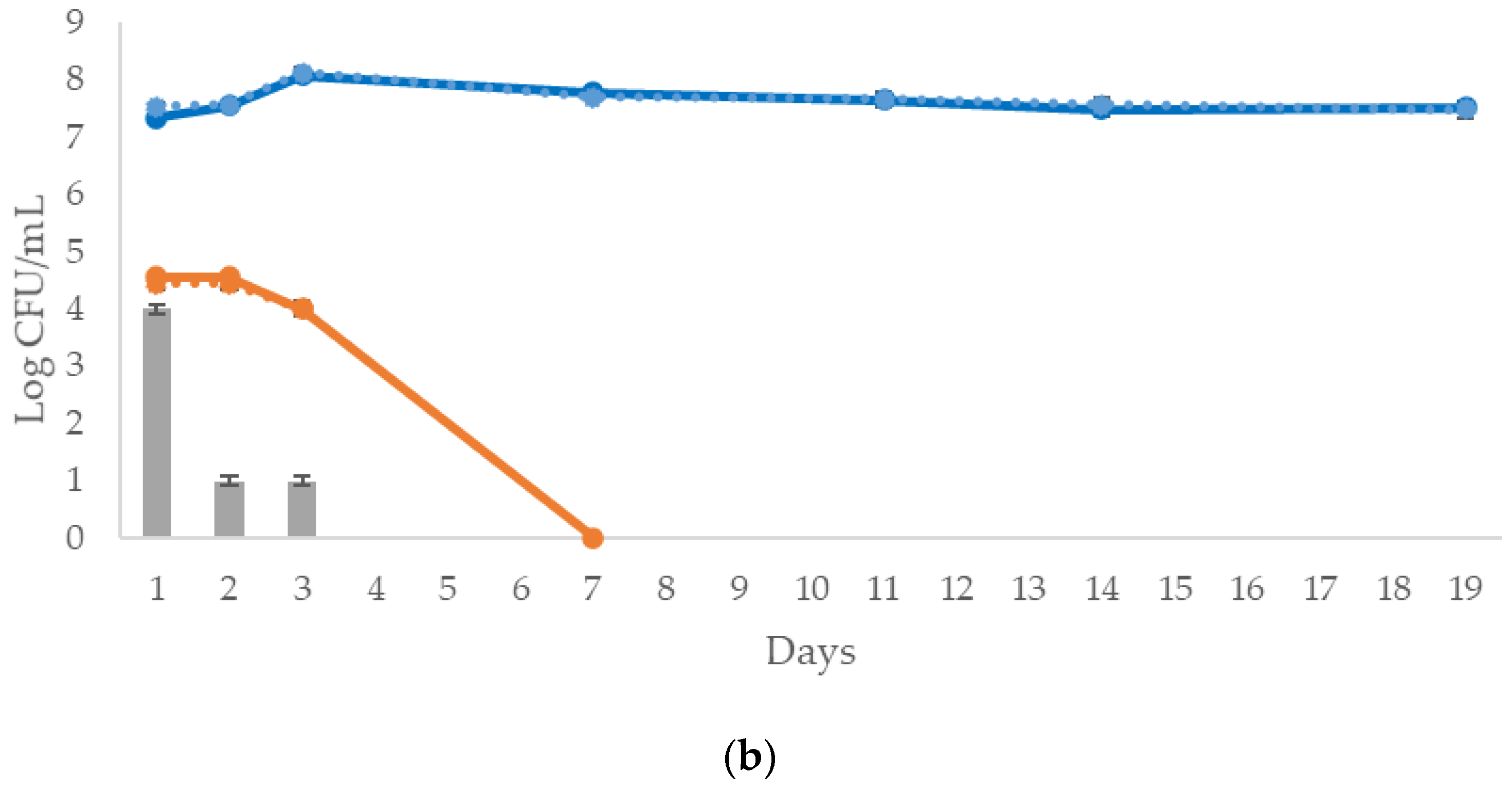
 population in both (a) 2021 vintage and (b) 2022 vintage wines.
population in both (a) 2021 vintage and (b) 2022 vintage wines.  S. cerevisiae in the control trial; S. cerevisiae in M. pulcherrima cold clarification;
S. cerevisiae in the control trial; S. cerevisiae in M. pulcherrima cold clarification;  WY in the control trial;
WY in the control trial;  WY in M. pulcherrima cold clarification;
WY in M. pulcherrima cold clarification;  M. pulcherrima biomass in the trial with M. pulcherrima cold clarification.
population in both (a) 2021 vintage and (b) 2022 vintage wines. S. cerevisiae in the control trial; S. cerevisiae in M. pulcherrima cold clarification; WY in the control trial; WY in M. pulcherrima cold clarification; M. pulcherrima biomass in the trial with M. pulcherrima cold clarification.
M. pulcherrima biomass in the trial with M. pulcherrima cold clarification.
population in both (a) 2021 vintage and (b) 2022 vintage wines. S. cerevisiae in the control trial; S. cerevisiae in M. pulcherrima cold clarification; WY in the control trial; WY in M. pulcherrima cold clarification; M. pulcherrima biomass in the trial with M. pulcherrima cold clarification.


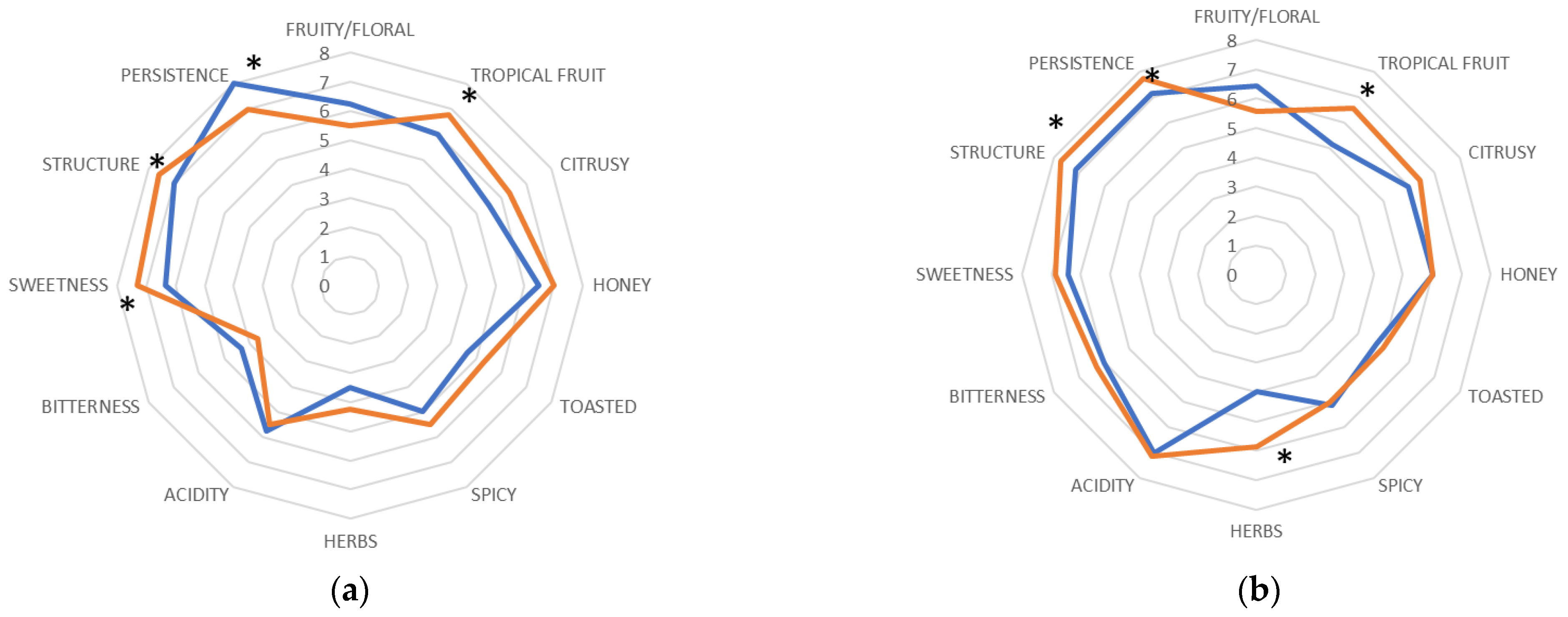
 control trial;
control trial;  M. pulcherrima in cold clarification. *, significantly different (Fischer ANOVA; p-value < 0.05).
control trial; M. pulcherrima in cold clarification. *, significantly different (Fischer ANOVA; p-value < 0.05).
M. pulcherrima in cold clarification. *, significantly different (Fischer ANOVA; p-value < 0.05).
control trial; M. pulcherrima in cold clarification. *, significantly different (Fischer ANOVA; p-value < 0.05).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2021 | 2022 | |||||
|---|---|---|---|---|---|---|
| Pied de cuve | Byotype | % | n. Strains | Byotype | % | Strains |
| I | 12.5 | 2 | II | 67.0 | 12 | |
| II | 37.5 | 6 | III | 11.0 | 2 | |
| III | 12.5 | 2 | VII | 11.0 | 2 | |
| IV | 12.5 | 2 | VIII | 11.0 | 2 | |
| V | 25.0 | 4 | - | - | - | |
| Fermentation control | VI | 10.0 | 1 | II | 40.0 | 4 |
| I | 20.0 | 2 | III | - | - | |
| II | 30.0 | 3 | VIII | 20.0 | 2 | |
| III | - | - | I | 20.0 | 2 | |
| IV | 40.0 | 4 | IX | 20.0 | 2 | |
| M. pulcherrima in cold clarification | VI | 42.0 | 5 | I | 50.0 | 4 |
| I | 17.0 | 2 | II | 50.0 | 4 | |
| II | 17.0 | 2 | III | - | - | |
| III | 8.0 | 1 | - | - | - | |
| IV | 17.0 | 2 | - | - | ||
| M. pulcherrima in Cold Clarification (2021) | Control Trial (2021) | M. pulcherrima in Cold Clarification (2022) | Control Trial (2022) | |
|---|---|---|---|---|
| Ethanol %v/v | 15.05 ± 0.02 a | 14.49 ± 0.01 b | 14.81 ± 0.15 a | 15.08 ± 0.02 a |
| Total acidity (g/L tartaric acid) | 6.26 ± 0.09 a | 6.52 ± 0.01 a | 7.56 ± 0.12 a | 7.84 ± 0.14 a |
| Sugar residue (g/L) | 3.7 ± 0.02 b | 14.0 ± 0.03 a | 6.6 ± 0.7 a | 1.5 ± 0.1 b |
| pH | 3.28 ± 0.02 a | 3.25 ± 0.00 a | 3.08 ± 0.00 a | 3.09 ± 0.00 a |
| Volatile acidity (g/L acetic acid) | 0.64 ± 0.01 a | 0.56 ± 0.02 b | 0.34 ± 0.02 a | 0.36 ± 0.03 a |
| Total SO2 (mg/L) | 25 ± 0.9 a | 26 ± 0.8 a | 21 ± 0.9 a | 21 ± 0.9 a |
| Malic acid (g/L) | 0.55 ± 0.03 a | 0.49 ± 0.02 a | 0.54 ± 0.05 a | 0.55 ± 0.05 a |
| Ethanol yield (g/g) | 0.45 ± 0.07 a | 0.45 ± 0.06 a | 0.44 ± 0.09 a | 0.44 ± 0.09 a |
| M. pulcherrima in Cold Clarification 2021 | OAV (Odor Activity Value) | Control Trial 2021 | OAV (Odor Activity Value) | M. pulcherrima in Cold Clarification 2022 | OAV (Odor Activity Value) | Control Trial 2022 | OAV (Odor Activity Value) | |
|---|---|---|---|---|---|---|---|---|
| Esters (mg/L) | ||||||||
| Ethyl butyrate | 0.13 ± 0.014 a | 0.325 | 0.13 ± 0.00 a | 0.325 | 0.187 ± 0.007 a | 0.467 | 0.148 ± 0.006 b | 0.445 |
| Ethyl acetate | 35.75 ± 0.41 a | 2.97 | 35.55 ± 0.36 a | 2.96 | 30.12 ± 0.18 b | 2.51 | 31.91 ± 0.35 a | 2.65 |
| Phenyl ethyl acetate | 0.89 ± 0.042 a | 12.19 | 0.88 ± 0.034 a | 12.05 | 0.821 ± 0.032 a | 11.24 | 0.773 ± 0.045 a | 10.58 |
| Ethyl hexanoate | 1.80 ± 0.121 a | 22.5 | 0.63 ± 0.142 b | 7.87 | 1.470 ± 0.050 a | 18.37 | 0.612 ± 0.043 b | 7.65 |
| Ethyl octanoate | 0.00 ± 0.00 a | 0 | 0.00 ± 0.00 a | 0 | 0.002 ± 0.000 a | 0.003 | 0.003 ± 0.001 a | 0.005 |
| Isoamyl acetate | 1.27 ± 0.026 b | 7.93 | 2.22 ± 0.147 a | 13.87 | 1.484 ± 0.112 a | 9.27 | 1.404 ± 0.024 a | 8.77 |
| Alcohols (mg/L) | ||||||||
| n- propanol | 30.88 ± 0.36 a | 0.100 | 26.08 ± 0.13 b | 0.08 | 36.44 ± 4.92 a | 0.119 | 31.64 ± 0.05 a | 0.103 |
| Isobutanol | 20.48 ± 0.29 a | 0.512 | 18.30 ± 0.10 b | 0.45 | 14.09 ± 0.34 a | 2.83 | 11.41 ± 0.24 b | 0.285 |
| Amyl alcohol | 13.00 ± 0.55 a | 0.203 | 11.69 ± 0.41 a | 0.18 | 10.03 ± 0.29 a | 0.156 | 8.64 ± 0.48 a | 0.135 |
| Isoamyl alcohol | 113.94 ± 0.05 a | 1.89 | 114.16 ± 0.24 a | 1.90 | 89.94 ± 0.12 a | 1.49 | 80.95 ± 0.52 b | 1.34 |
| β-Phenyl Ethanol | 55.9 ± 0.130 b | 3.99 | 64.5 ± 0.152 a | 4.60 | 14.01 ± 0.022 a | 1 | 13.32 ± 0.045 a | 0.951 |
| Carbonyl Compounds (mg/L) | ||||||||
| Acetaldehyde | 63.16 ± 0.28 a | 126.32 | 25.35 ± 0.37 b | 70 | 22.39 ± 0.59 a | 44.78 | 16.57 ± 0.1 b | 33.14 |
| Monoterpenes (mg/L) | ||||||||
| Linalool | 0.015 ± 0.001 a | 0.60 | 0.012 ± 0.002 a | 0.48 | 0.010 ± 0.00 a | 0.40 | 0.009 ± 0.00 a | 0.36 |
| Geraniol | 0.010 ± 0.002 a | 0.33 | 0.002 ± 0.000 b | 0.06 | 0.016 ± 0.00 a | 0.53 | 0.005 ± 0.001 b | 0.17 |
| Nerol | 0.023 ± 0.001 a | 1.53 | 0.005 ± 0.000 b | 0.33 | 0.001 ± 0.000 a | 0.06 | 0.001 ± 0.00 a | 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agarbati, A.; Canonico, L.; Ciani, M.; Comitini, F. Metschnikowia pulcherrima in Cold Clarification: Biocontrol Activity and Aroma Enhancement in Verdicchio Wine. Fermentation 2023, 9, 302. https://doi.org/10.3390/fermentation9030302
Agarbati A, Canonico L, Ciani M, Comitini F. Metschnikowia pulcherrima in Cold Clarification: Biocontrol Activity and Aroma Enhancement in Verdicchio Wine. Fermentation. 2023; 9(3):302. https://doi.org/10.3390/fermentation9030302
Chicago/Turabian StyleAgarbati, Alice, Laura Canonico, Maurizio Ciani, and Francesca Comitini. 2023. "Metschnikowia pulcherrima in Cold Clarification: Biocontrol Activity and Aroma Enhancement in Verdicchio Wine" Fermentation 9, no. 3: 302. https://doi.org/10.3390/fermentation9030302
APA StyleAgarbati, A., Canonico, L., Ciani, M., & Comitini, F. (2023). Metschnikowia pulcherrima in Cold Clarification: Biocontrol Activity and Aroma Enhancement in Verdicchio Wine. Fermentation, 9(3), 302. https://doi.org/10.3390/fermentation9030302