
Identification of Molecular Markers for Early Detection of Sluggish Fermentation Associated with Heat Shock during Alcoholic Fermentation

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
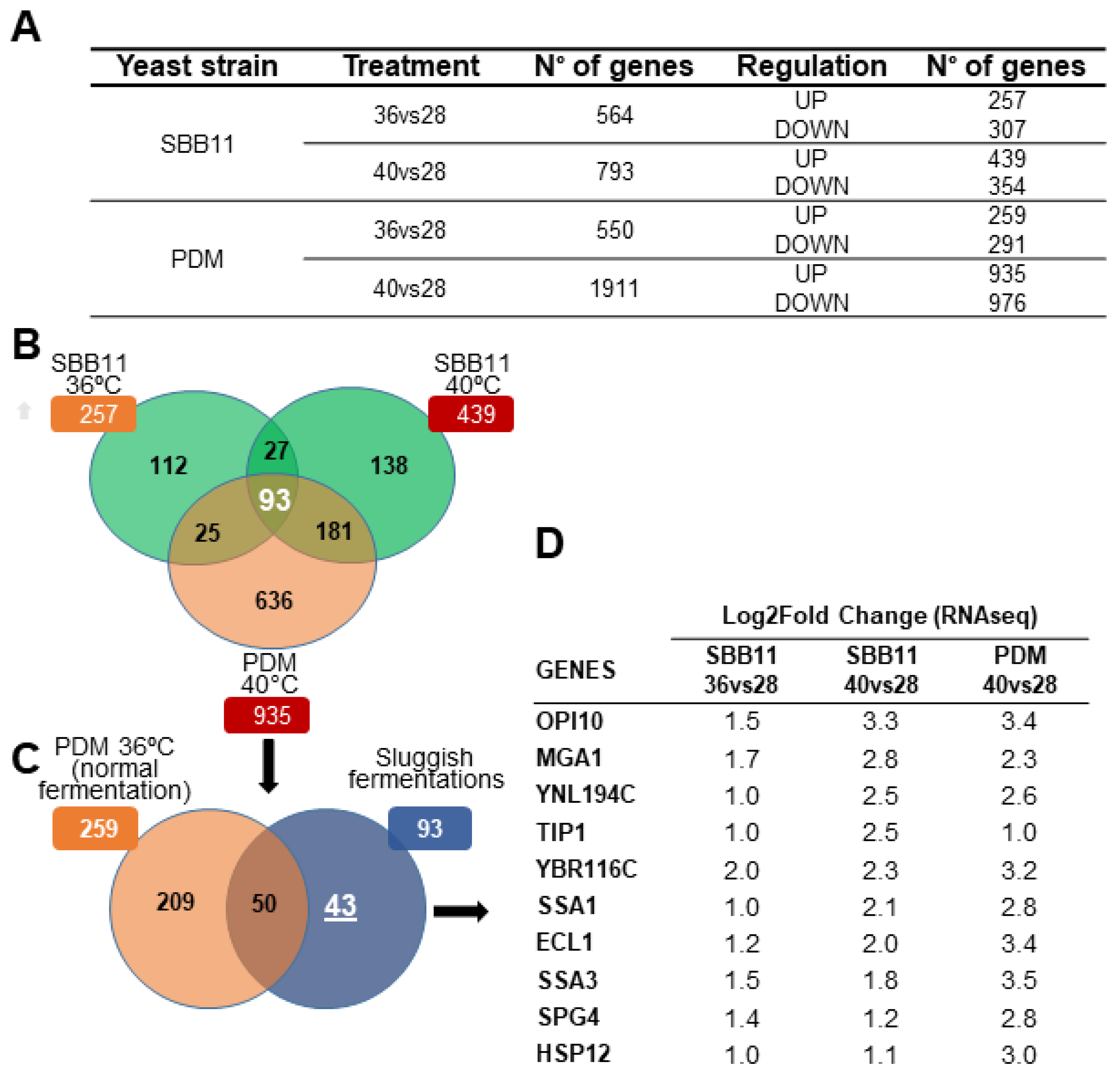
2.1. Transcriptomic Analysis and Candidate Gene Selection
2.2. Yeast Strains and Inoculum Preparation
2.3. Microvinifications
2.4. RNA Extraction and cDNA Synthesis
2.5. Gene Expression Analysis by Quantitative Real-Time PCR (qRT-PCR)
2.6. Statistical Analyses
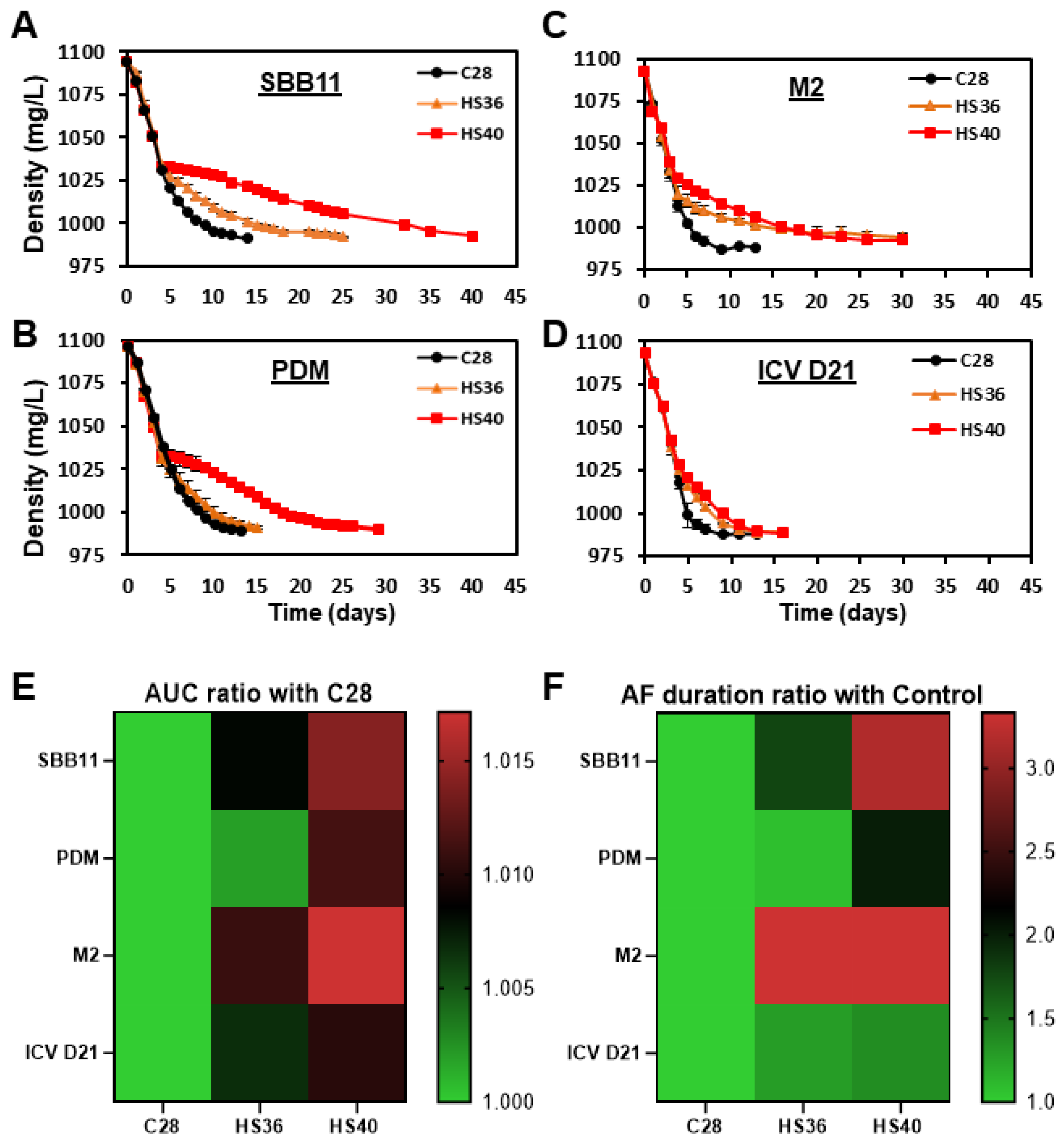
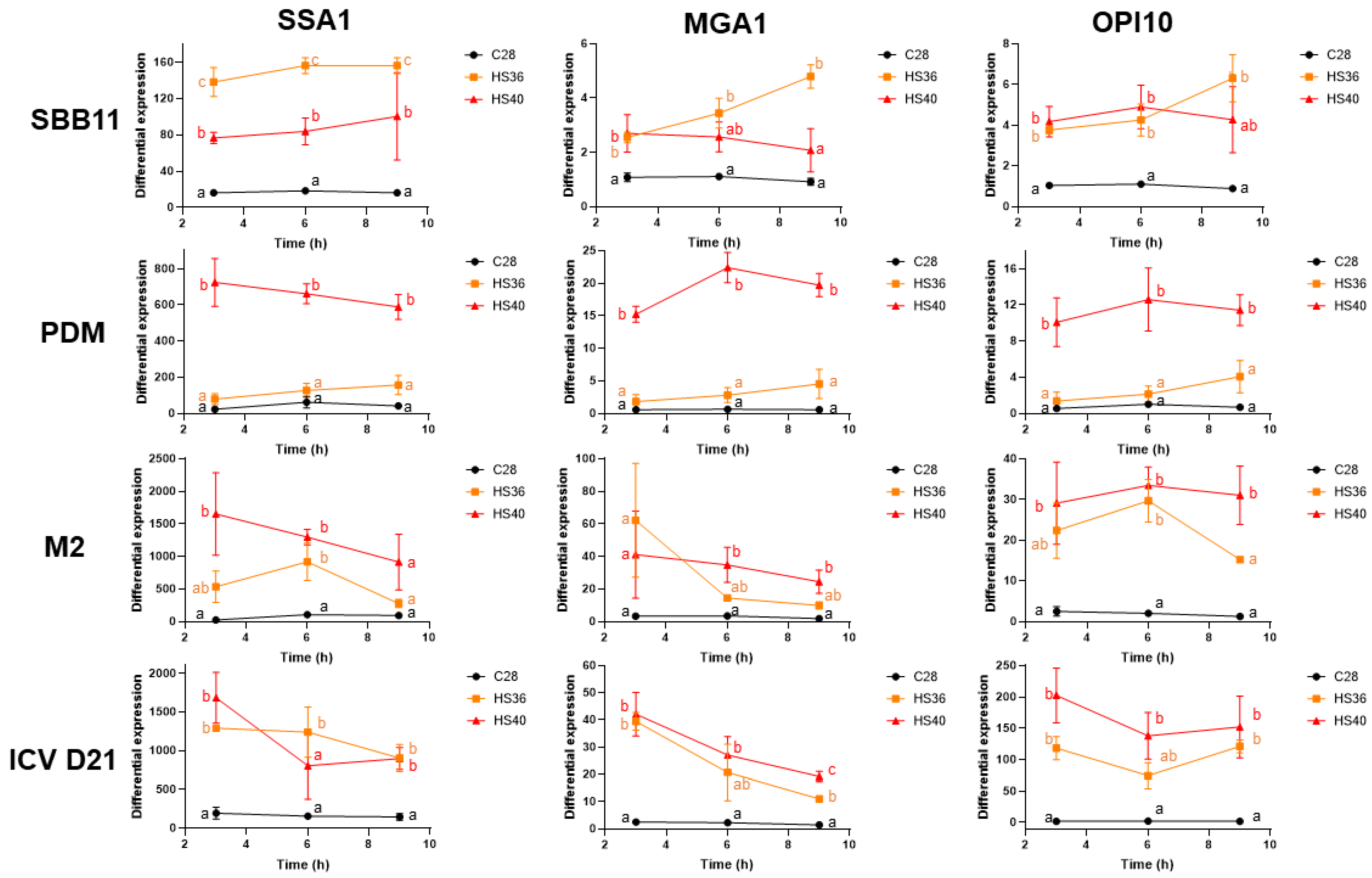
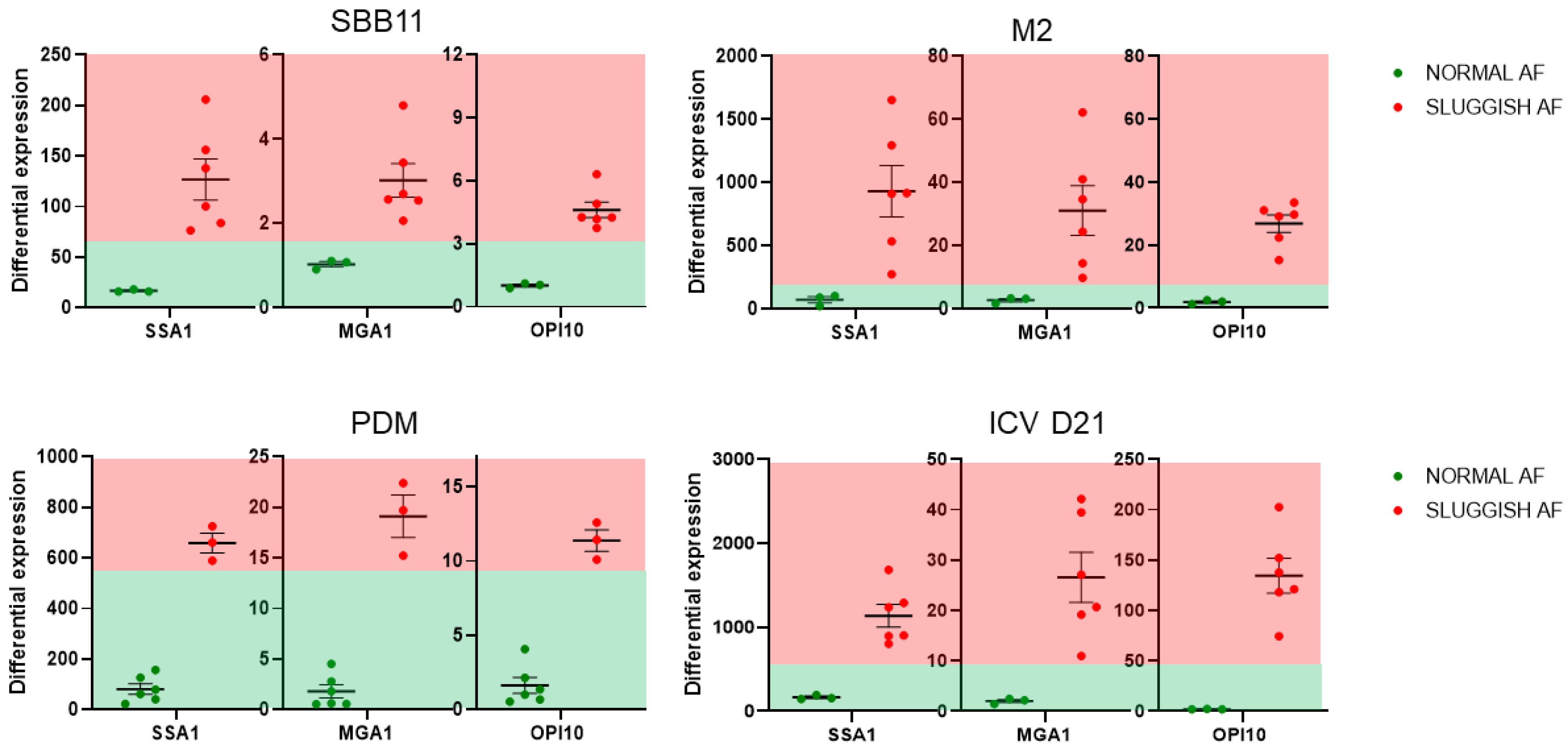
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. Handbook of Enology; Wiley: Hoboken, NJ, USA, 2006. [Google Scholar]
- Walker, G.M.; Stewart, G.G. Saccharomyces cerevisiae in the Production of Fermented Beverages. Beverages 2016, 2, 30. [Google Scholar] [CrossRef]
- Maicas, S. The Role of Yeasts in Fermentation Processes. Microorganisms 2020, 8, 1142. [Google Scholar] [CrossRef] [PubMed]
- Bisson, L.F. Stuck and Sluggish Fermentations. Am. J. Enol. Vitic. 1999, 50, 107–119. [Google Scholar] [CrossRef]
- Maisonnave, P.; Sanchez, I.; Moine, V.; Dequin, S.; Galeote, V. Stuck fermentation: Development of a synthetic stuck wine and study of a restart procedure. Int. J. Food Microbiol. 2013, 163, 239–247. [Google Scholar] [CrossRef]
- Malherbe, S.; Bauer, F.F.; Du Toit, M. Understanding problematic fermentations–A review. S. Afr. J. Enol. Vitic. 2007, 2, 169–186. [Google Scholar]
- Vargas-Trinidad, A.S.; Lerena, M.C.; Alonso-del-Real, J.; Esteve-Zarzoso, B.; Mercado, L.A.; Mas, A.; Querol, A.; Combina, M. Effect of transient thermal shocks on alcoholic fermentation performance. Int. J. Food Microbiol. 2020, 312, 108362. [Google Scholar] [CrossRef]
- Gamero-Sandemetrio, E.; Gómez-Pastor, R.; Matallana, E. Antioxidant defense parameters as predictive biomarkers for fermentative capacity of active dried wine yeast. Biotechnol. J. 2014, 9, 1055–1064. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Chiva, R.; Beltran, G.; Mas, A.; Guillamon, J.M. Biomarkers for detecting nitrogen deficiency during alcoholic fermentation in different commercial wine yeast strains. Food Microbiol. 2013, 34, 227–237. [Google Scholar] [CrossRef]
- Lerena, M.C. Global Gene Expression Analysis of S. cerevisiae Strains during Sluggish Fermentation Caused by Heat Shock; CONICET: Mendoza, Argentina, 2022. [Google Scholar]
- Bely, M.; Sablayrolles, J.M.; Barre, P. Automatic detection of assimilable nitrogen deficiencies during alcoholic fermentation in oenological conditions. J. Ferment Bioeng. 1990, 70, 246–252. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.H.; Karlen, Y.; Bakker, O.; van den Hoff, M.J.B.; Moorman, A.F.M. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef]
- Ramakers, C.; Ruijter, J.M.; Lekanne Deprez, R.H.; Moorman, A.F.M. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Warringer, J.; Zörgö, E.; Cubillos, F.A.; Zia, A.; Gjuvsland, A.; Simpson, J.T.; Forsmark, A.; Durbin, R.; Omholt, S.W.; Louis, E.J.; et al. Trait Variation in Yeast Is Defined by Population History. PLoS Genet. 2011, 7, e1002111. [Google Scholar] [CrossRef]
- García-Ríos, E.; Gutiérrez, A.; Salvadó, Z.Z.; Arroyo-López, F.N.; Guillamon, J.M. The Fitness Advantage of Commercial Wine Yeasts in Relation to the Nitrogen Concentration, Temperature, and Ethanol Content under Microvinification Conditions. Appl. Env. Microbiol. 2014, 80, 704–713. [Google Scholar] [CrossRef]
- Ganucci, D.; Guerrini, S.; Mangani, S.; Vincenzini, M.; Granchi, L. Quantifying the effects of ethanol and temperature on the fitness advantage of predominant Saccharomyces cerevisiae strains occurring in spontaneous wine fermentations. Front. Microbiol. 2018, 9, 1563. [Google Scholar] [CrossRef]
- Riles, L.; Fay, J.C. Genetic basis of variation in heat and ethanol tolerance in Saccharomyces cerevisiae. G3: Genes Genomes Genet. 2019, 9, 179–188. [Google Scholar] [CrossRef]
- Coulter, A.D.; Henschke, P.A.; Simos, C.A.; Pretorius, I.S. When the heat is on, yeast fermentation runs out of puff. Aust. N. Z. Wine Ind. 2008, 23, 29–33. [Google Scholar]
- Schmid, F.; Schadt, J.; Jiranek, V.; Block, D.E. Formation of temperature gradients in large- and small-scale red wine fermentations during cap management. Aust. J. Grape Wine Res. 2009, 15, 249–255. [Google Scholar] [CrossRef]
- Valentine, G.D.S.; Walker, M.E.; Gardner, J.M.; Schmid, F.; Jiranek, V. Brief temperature extremes during wine fermentation: Effect on yeast viability and fermentation progress. Aust. J. Grape Wine Res. 2019, 25, 62–69. [Google Scholar] [CrossRef]
- Marullo, P.; Mansour, C.; Dufour, M.; Albertin, W.; Sicard, D.; Bely, M.; Dubourdieu, D. Genetic improvement of thermo-tolerance in wine Saccharomyces cerevisiae strains by a backcross approach. FEMS Yeast Res. 2009, 9, 1148–1160. [Google Scholar] [CrossRef]
- Gasch, A.P.; Spellman, P.T.; Kao, C.M.; Carmel-Harel, O.; Eisen, M.B.; Storz, G.; Botstein, D.; Brown, P.O. Genomic Expression Programs in the Response of Yeast Cells to Environmental Changes. Mol. Biol. Cell 2000, 12, 4241–4257. [Google Scholar] [CrossRef] [PubMed]
- Morano, K.A.; Grant, C.M.; Moye-Rowley, W.S. The response to heat shock and oxidative stress in Saccharomyces cerevisiae. Genetics 2012, 190, 1157–1195. [Google Scholar] [CrossRef] [PubMed]
- Strassburg, K.; Walther, D.; Takahashi, H.; Kanaya, S.; Kopka, J. Dynamic Transcriptional and Metabolic Responses in Yeast Adapting to Temperature Stress. OMICS 2010, 14, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Kampinga, H.H.; Craig, E.A. The Hsp70 chaperone machinery: J-proteins as drivers of functional specificity. Nat. Rev. Mol. Cell Biol. 2010, 11, 579–592. [Google Scholar] [CrossRef]
- Lee, D.H.; Sherman, M.Y.; Goldberg, A.L. The requirements of yeast Hsp70 of SSA family for the ubiquitin-dependent degradation of short-lived and abnormal proteins. Biochem. Biophys Res. Commun. 2016, 475, 100–106. [Google Scholar] [CrossRef]
- Lotz, S.K.; Knighton, L.E.; Nitika Jones, G.W.; Truman, A.W. Not quite the SSAme: Unique roles for the yeast cytosolic Hsp70s. Curr. Genet. 2019, 65, 1127–1134. [Google Scholar] [CrossRef]
- Kaushik, S.; Cuervo, A.M. Chaperone-mediated autophagy: A unique way to enter the lysosome world. Trends Cell Biol. 2012, 22, 407–417. [Google Scholar] [CrossRef]
- Shiber, A.; Ravid, T. Chaperoning Proteins for Destruction: Diverse Roles of Hsp70 Chaperones and their Co-Chaperones in Targeting Misfolded Proteins to the Proteasome. Biomolecules 2014, 4, 704–724. [Google Scholar] [CrossRef]
- Gasch, A.P. The environmental stress response: A common yeast response to diverse environmental stresses. In Yeast Stress Responses. Topics in Current Genetics; Hohmann, S., Mager, W.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; Volume 1. [Google Scholar]
- Yu, T.; Takahashi, U.; Iwahashi, H. Transcriptome Analysis of the Influence of High-Pressure Carbon Dioxide on Saccharomyces cerevisiae under Sub-Lethal Condition. J. Fungi. 2022, 8, 1011. [Google Scholar] [CrossRef]
- Peffer, S.; Gonçalves, D.; Morano, K.A. Regulation of the Hsf1-dependent transcriptome via conserved bipartite contacts with Hsp70 promotes survival in yeast. J. Biol. Chem. 2019, 294, 12191–12202. [Google Scholar] [CrossRef]
- Zheng, X.; Krakowiak, J.; Patel, N.; Beyzavi, A.; Ezike, J.; Khalil, A.S.; Pincus, D. Dynamic control of Hsf1 during heat shock by a chaperone switch and phosphorylation. eLife 2016, 5, e1863. [Google Scholar] [CrossRef]
- Sunyer-Figueres, M.; Vázquez, J.; Mas, A.; Torija, M.J.; Beltran, G. Transcriptomic Insights into the Effect of Melatonin in Saccharomyces cerevisiae in the Presence and Absence of Oxidative Stress. Antioxidants 2020, 9, 947. [Google Scholar] [CrossRef]
- Proft, M.; Gibbons, F.D.; Copeland, M.; Roth, F.P.; Struhl, K. Genome wide Identification of Sko1 Target Promoters Reveals a Regulatory Network That Operates in Response to Osmotic Stress in Saccharomyces cerevisiae. Eukaryot. Cell 2005, 4, 1343. [Google Scholar] [CrossRef]
- Kim, I.S.; Choi, W.; Son, J.; Lee, J.H.; Lee, H.; Lee, J.; Shin, S.C.; Kim, H.W. Screening and Genetic Network Analysis of Genes Involved in Freezing and Thawing Resistance in DaMDHAR-Expressing Saccharomyces cerevisiae Using Gene Expression Profiling. Genes 2021, 12, 219. [Google Scholar] [CrossRef]
- Hancock, L.C.; Behta, R.P.; Lopes, J.M. Genomic Analysis of the Opi− Phenotype. Genetics 2006, 173, 621–634. [Google Scholar] [CrossRef]
- Tkach, J.M.; Yimit, A.; Lee, A.Y.; Riffle, M.; Costanzo, M.; Jaschob, D.; Hendry, J.A.; Ou, J.; Moffat, J.; Boone, C.; et al. Dissecting DNA damage response pathways by analysing protein localization and abundance changes during DNA replication stress. Nat. Cell Biol. 2012, 14, 966–976. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Fabregat, A.; Sidiropoulos, K.; Viteri, G.; Marin-Garcia, P.; Ping, P.; Stein, L.; D’Eustachio, P.; Hermjakob, H. Reactome diagram viewer: Data structures and strategies to boost performance. Bioinformatics 2018, 34, 1208–1214. [Google Scholar] [CrossRef]
- Longin, C.; Julliat, F.; Serpaggi, V.; Maupeu, J.; Bourbon, G.; Rousseaux, S.; Guilloux-Benatier, M.; Alexandre, H. Evaluation of three Brettanomyces qPCR commercial kits: Results from an interlaboratory study. OENO One 2016, 50, 223–230. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Name | Oligonucleotide Sequence (5′-3′end) * |
|---|---|---|
| UBC6 | UBC6-F | TACTTGGAATCCTGGCTGGT |
| UBC6-R | GATCCTGTCGTGGCTTCATC | |
| SSA1 | SSA1-F | GAAGTCCGAGATCTTTTCCACTT |
| SSA1-R | CCTCTTGGAGCTGGTGGAAT | |
| MGA1 | MGA1-F | ATCTCATCCTTCCCCAGACC |
| MGA1-R | ATTCAAGATACCGGCGTTGG | |
| OPI10 | OPI10-F | CCGCTGATCCGTTTACTGAC |
| OPI10-R | TTCCTTGTTCTCGAGGCTCA | |
| YNL194C | YNL194C-F | GATACTAGCAGGTGGCAGGA |
| YNL194C-R | TTAAAGCCCGAAGTGGATGC | |
| TIP1 | TIP1-F | ATCGCTGCTGCTCTTGCCT |
| TIP1-R | AGCGGCAGAGGATGTAGCTT | |
| YBR116C | YBR116C-F | GTGTTGCGTCAAGGGCTGAA |
| YBR116C-R | GGCAGCAAGTGACCATCAACC | |
| ECL1 | ECL1-F | TGCTCCGAAGATTGTAAGCTG |
| ECL1-R | CGGTGGAGTGAGATTATGCG | |
| SSA3 | SSA3-F | AGGTAGGCTCTCGAAGGATG |
| SSA3-R | GTTCTGCCTCCCTTTCATCG | |
| SPG4 | SPG4-F | GGAGACAGTAAAACGCAGGT |
| SPG4-R | ACATCGGAACTGTCCTGTGA | |
| HSP12 | HSP12-F | CAAGGTCGCTGGTAAGGTTC |
| HSP12-R | ACCTTCAGCGTTATCCTTGC |
| Yeast/Treatment | AUC ± SD (Arbitrary Units) | ||
|---|---|---|---|
| C28 | HS36 | HS40 | |
| SBB11 | 11,310.38 ± 3.34 (a) | 11,403.32 ± 15.54 (b) | 11,472.93 ± 1.7 (c) |
| PDM | 11,334.02 ± 3.71 (a) | 11,355.47 ± 33.3 (a) | 11,463.07 ± 30.47 (b) |
| M2 | 11,167.47 ± 25.76 (a) | 11,289.18 ± 28.9 (b) | 11,358.97 ± 5.85 (c) |
| ICV D21 | 11,183.18 ± 25.36 (a) | 11,257.82 ± 18.56 (b) | 11,297.42 ± 5.21 (c) |
| AF duration ± SD (days) | |||
| SBB11 | 12.33 ± 0.57 (a) | 22 ± 1 (b) | 39.66 ± 1.52 (c) |
| PDM | 13.33 ± 0.57 (a) | 14.33 ± 0.57 (a) | 27 ± 1.73 (b) |
| M2 | 9.67 ± 1.15 (a) | 30.67 ± 0.58 (b) | 30.33 ± 0.58 (b) |
| ICV D21 | 9.67 ± 1.15 (a) | 13.33 ± 0.58 (b) | 13.67 ± 1.15 (b) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lerena, M.C.; Vargas-Trinidad, A.S.; Alonso-del-Real, J.; Rojo, M.C.; González, M.L.; Mercado, L.A.; Lijavetzky, D.C.; Querol, A.; Combina, M. Identification of Molecular Markers for Early Detection of Sluggish Fermentation Associated with Heat Shock during Alcoholic Fermentation. Fermentation 2023, 9, 313. https://doi.org/10.3390/fermentation9030313
Lerena MC, Vargas-Trinidad AS, Alonso-del-Real J, Rojo MC, González ML, Mercado LA, Lijavetzky DC, Querol A, Combina M. Identification of Molecular Markers for Early Detection of Sluggish Fermentation Associated with Heat Shock during Alcoholic Fermentation. Fermentation. 2023; 9(3):313. https://doi.org/10.3390/fermentation9030313
Chicago/Turabian StyleLerena, María Cecilia, Andrea Susana Vargas-Trinidad, Javier Alonso-del-Real, Maria Cecilia Rojo, Magalí Lucía González, Laura Analía Mercado, Diego Claudio Lijavetzky, Amparo Querol, and Mariana Combina. 2023. "Identification of Molecular Markers for Early Detection of Sluggish Fermentation Associated with Heat Shock during Alcoholic Fermentation" Fermentation 9, no. 3: 313. https://doi.org/10.3390/fermentation9030313
APA StyleLerena, M. C., Vargas-Trinidad, A. S., Alonso-del-Real, J., Rojo, M. C., González, M. L., Mercado, L. A., Lijavetzky, D. C., Querol, A., & Combina, M. (2023). Identification of Molecular Markers for Early Detection of Sluggish Fermentation Associated with Heat Shock during Alcoholic Fermentation. Fermentation, 9(3), 313. https://doi.org/10.3390/fermentation9030313