
Single-Cell Optical Nanomotion of Candida albicans in Microwells for Rapid Antifungal Susceptibility Testing

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Cultivation
2.2. Microfluidic Chip Construction
2.3. Computational Simulation of Fluid Flow in the Microfluidic Chip
2.4. Microfluidic Chip Setup
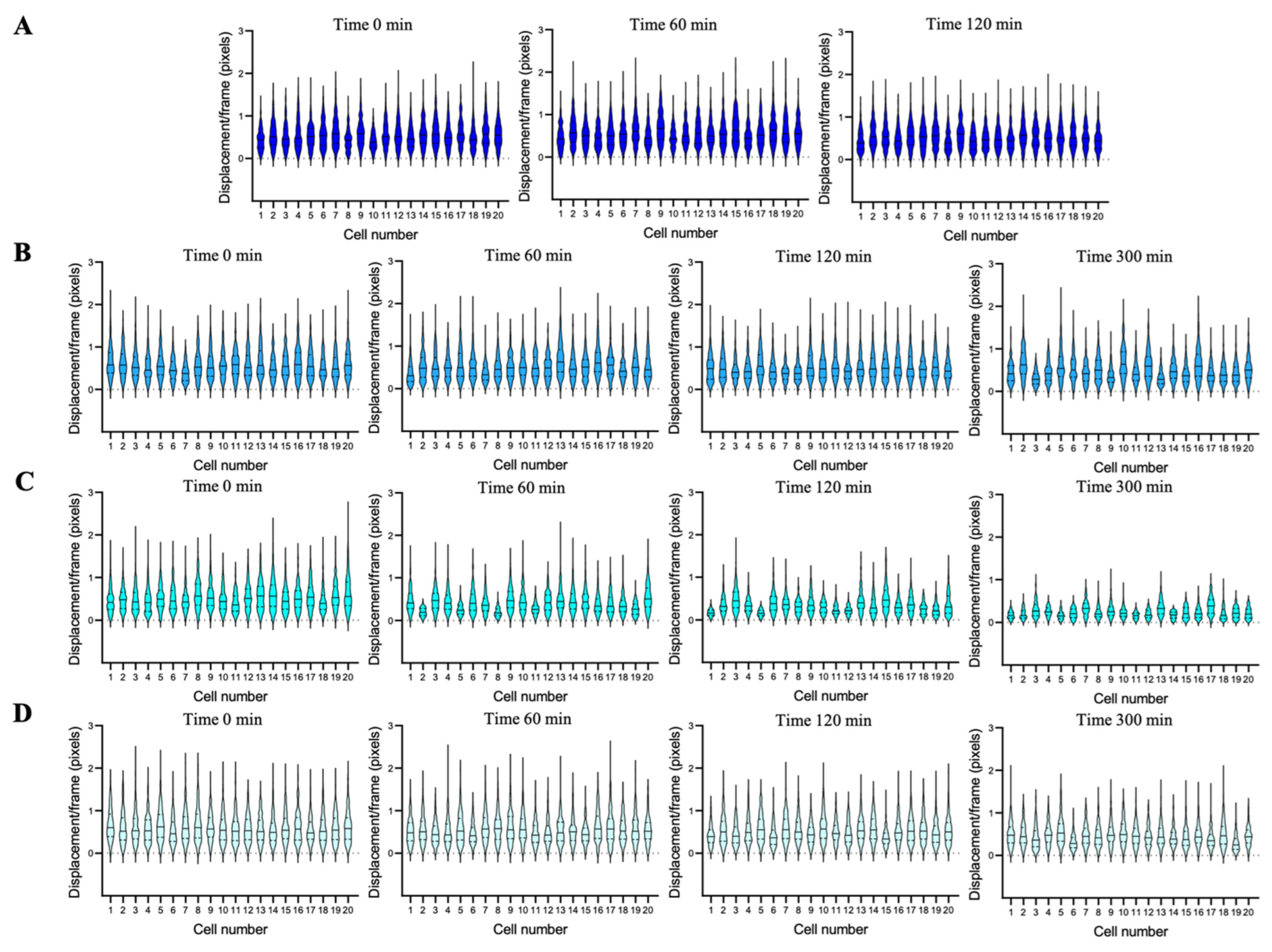
2.5. Nanomotion Measurement and Analysis
2.6. Cell Nanomotion and Viability Evaluation
3. Results
3.1. Construction of the Microfluidic Chip
3.1.1. Selection of the Microwell Shape and Dimensions
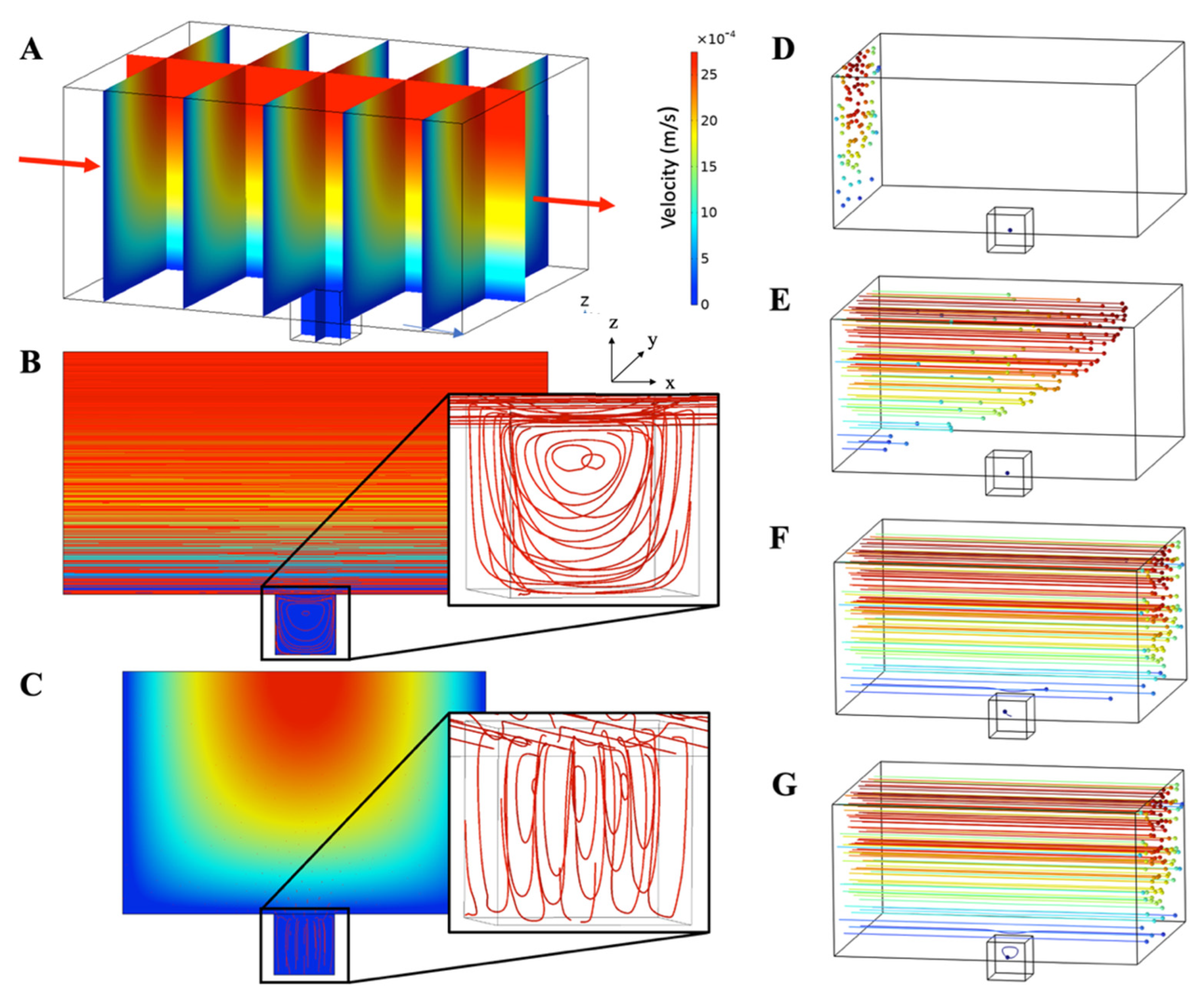
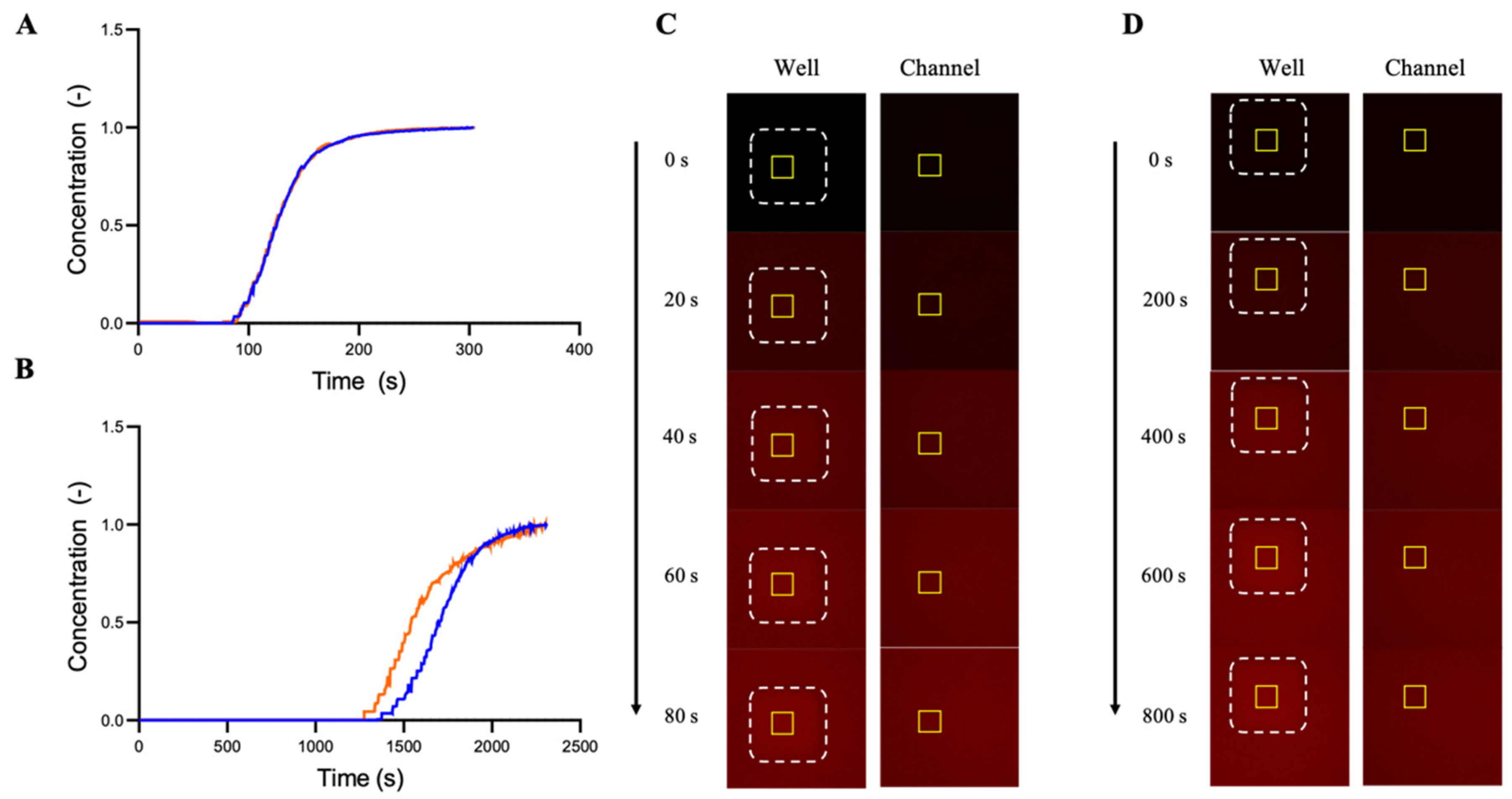
3.1.2. Microfluidic Chip Design and Modelling of Fluid Flow and Mass Transport
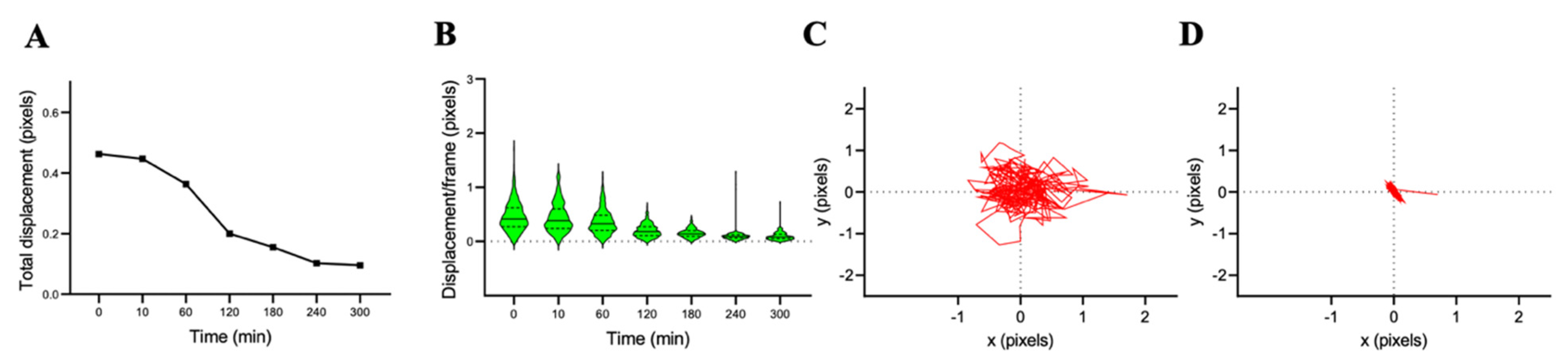
3.2. Cell Nanomotion and Susceptibility for the Antifungals Caspofungin and Fluconazole
3.2.1. C. albicans CAF2-1 Susceptibility for Caspofungin
3.2.2. C. albicans CAF2-1 Susceptibility for Fluconazole
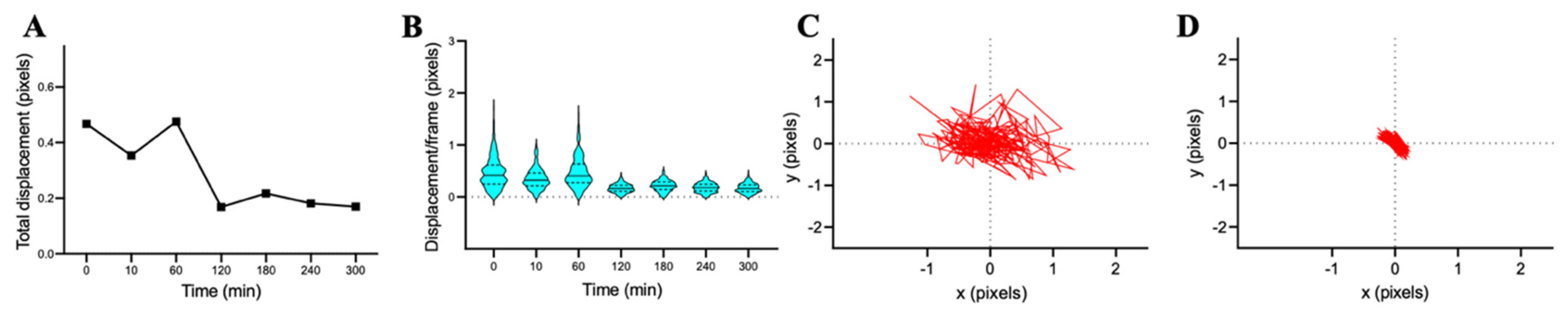
3.2.3. C. albicans DSY1024 Susceptibility for Caspofungin and Fluconazole
ONMD of C. albicans DSY1014 Treated with Fluconazole
ONMD of C. albicans DSY1014 Treated with Caspofungin
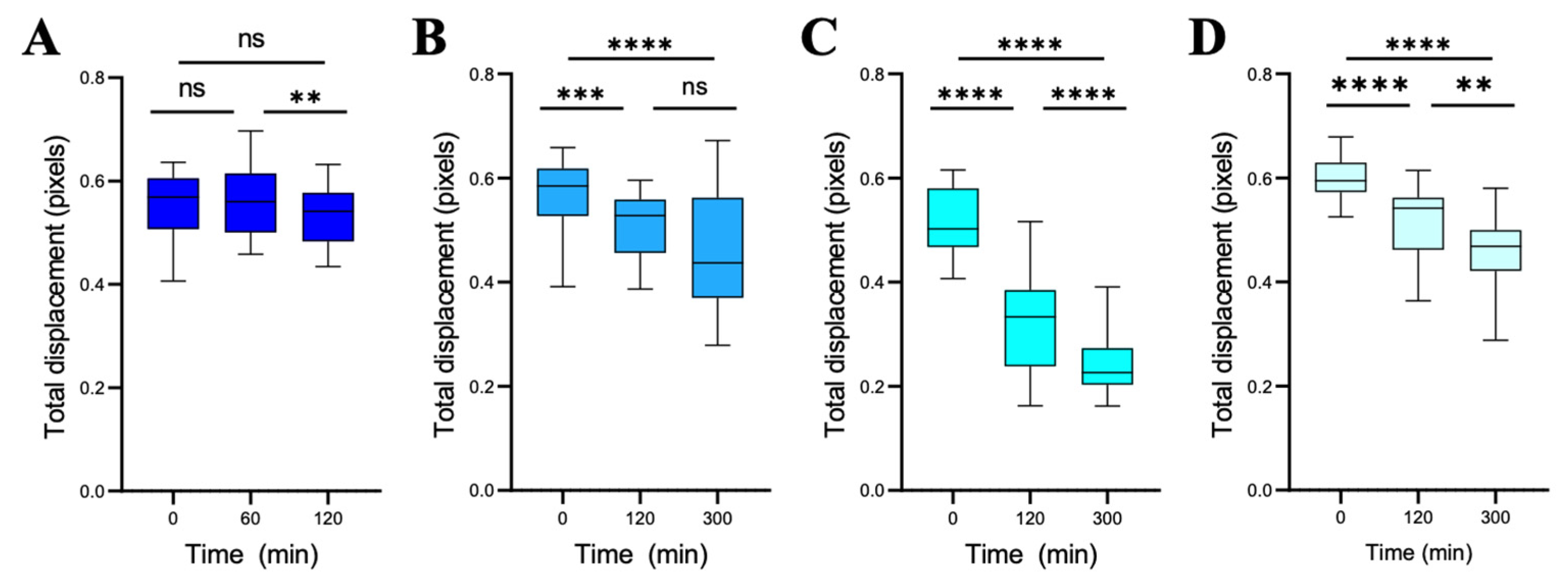
3.2.4. Sensitivity Analysis Based on the Evolution of the Slopes of the Total Displacements
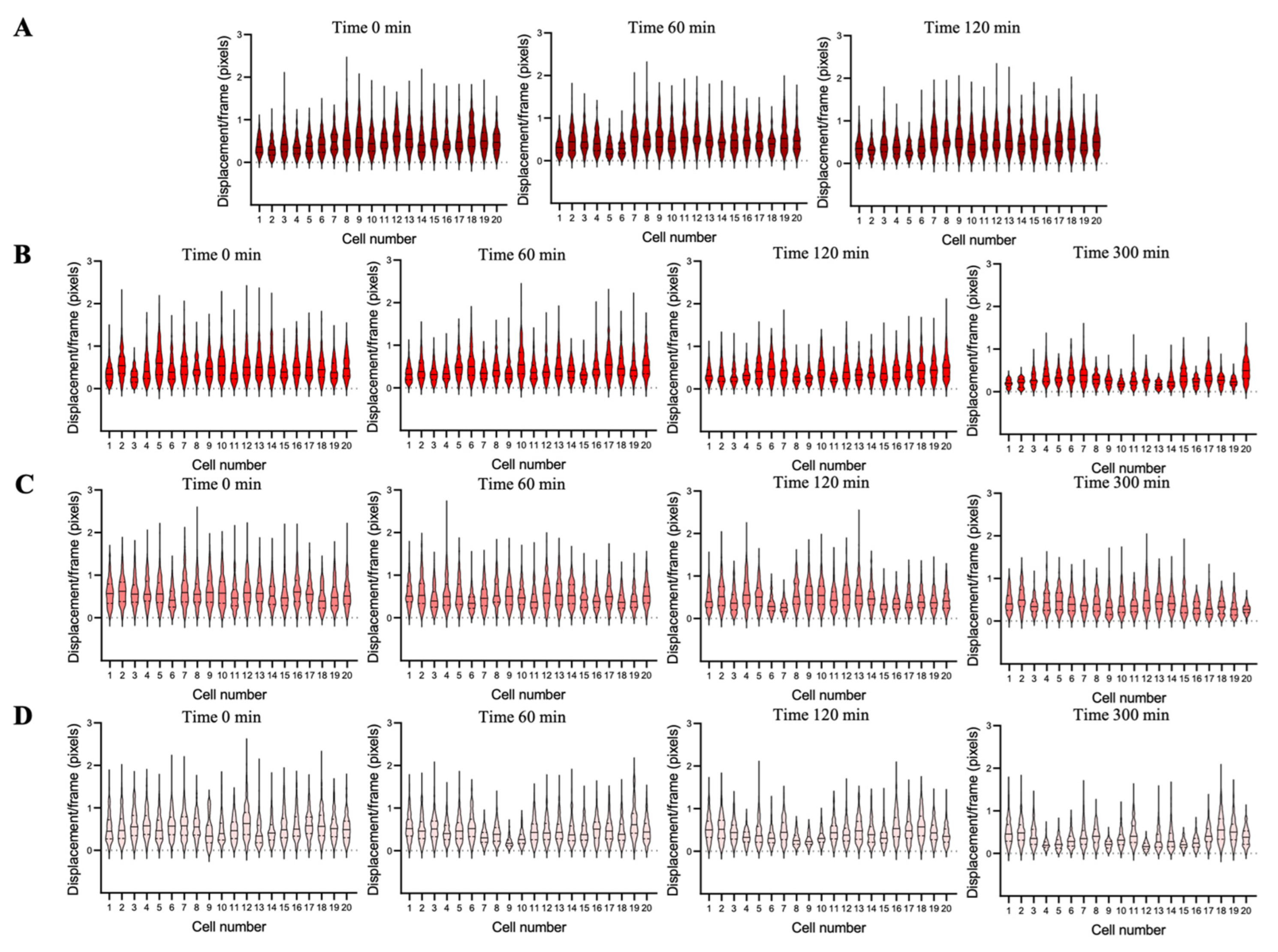
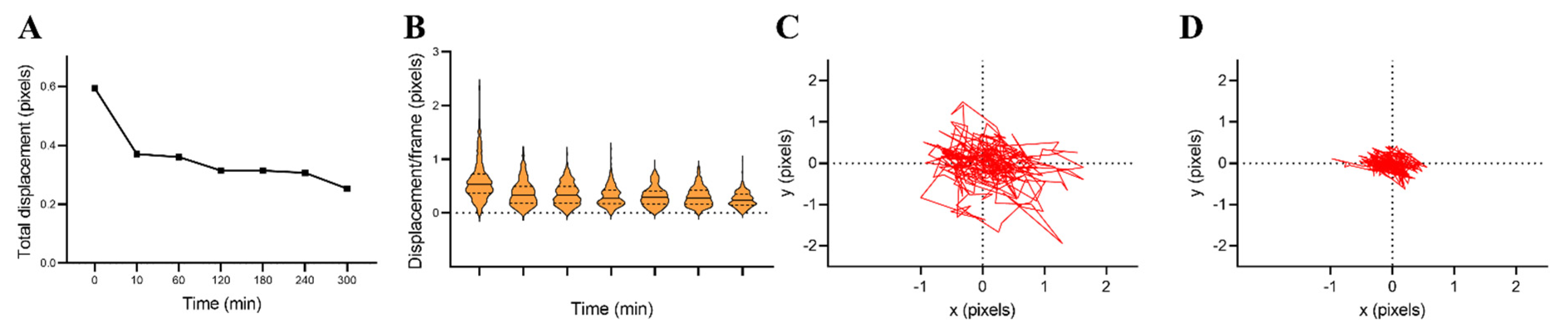
3.3. Cellular Nanomotion of Living Cells Compared to Dead Cells
4. Discussion

5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lopes, J.P.; Lionakis, M.S. Pathogenesis and virulence of Candida albicans. Virulence 2021, 13, 89–121. [Google Scholar] [CrossRef]
- Dadar, M.; Tiwari, R.; Karthik, K.; Chakraborty, S.; Shahali, Y.; Dhama, K. Candida albicans—Biology, molecular characterization, pathogenicity, and advances in diagnosis and control—An update. Microb. Pathog. 2018, 117, 128–138. [Google Scholar] [CrossRef]
- Poulain, D. Candida albicans, plasticity and pathogenesis. Crit. Rev. Microbiol. 2015, 41, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Pilmis, B.; Yang, Z.; Lanternier, F.; Lortholary, O. Systemic Candidiasis. In Infectious Diseases; Elsevier: Amsterdam, The Netherlands, 2017; pp. 439–445.e1. [Google Scholar] [CrossRef]
- Durand, C.; Maubon, D.; Cornet, M.; Wang, Y.; Aldebert, D.; Garnaud, C. Can We Improve Antifungal Susceptibility Testing? Front. Cell. Infect. Microbiol. 2021, 11, 720609. [Google Scholar] [CrossRef]
- Knabl, L.; Lass-Flörl, C. Antifungal susceptibility testing in Candida species: Current methods and promising new tools for shortening the turnaround time. Expert Rev. Anti-infective Ther. 2020, 18, 779–787. [Google Scholar] [CrossRef]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi; Approved Standard—2nd ed. CLSI document M38-A2; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- Van Belkum, A.; Burnham, C.-A.D.; Rossen, J.W.A.; Mallard, F.; Rochas, O.; Dunne, W.M., Jr. Innovative and rapid antimicrobial susceptibility testing systems. Nat. Rev. Microbiol. 2020, 18, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Posteraro, B.; Torelli, R.; De Carolis, E.; Posteraro, P.; Sanguinetti, M. Antifungal susceptibility testing: Current role from the clinical laboratory perspective. Mediterr. J. Hematol. Infect. Dis. 2014, 6, e2014030. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.A. Antifungal Drug Resistance: Mechanisms, Epidemiology, and Consequences for Treatment. Am. J. Med. 2012, 125, S3–S13. [Google Scholar] [CrossRef] [PubMed]
- Hayden, R.T.; Clinton, L.K.; Hewitt, C.; Koyamatsu, T.; Sun, Y.; Jamison, G.; Perkins, R.; Tang, L.; Pounds, S.; Bankowski, M.J. Rapid Antimicrobial Susceptibility Testing Using Forward Laser Light Scatter Technology. J. Clin. Microbiol. 2016, 54, 2701–2706. [Google Scholar] [CrossRef] [Green Version]
- Johnson, W.L.; France, D.C.; Rentz, N.S.; Cordell, W.T.; Walls, F.L. Sensing bacterial vibrations and early response to antibiotics with phase noise of a resonant crystal. Sci. Rep. 2017, 7, 12138. [Google Scholar] [CrossRef] [Green Version]
- Cowger, T.A.; Yang, Y.; Rink, D.E.; Todd, T.; Chen, H.; Shen, Y.; Yan, Y.; Xie, J. Protein-Adsorbed Magnetic-Nanoparticle-Mediated Assay for Rapid Detection of Bacterial Antibiotic Resistance. Bioconjugate Chem. 2017, 28, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Kadiyala, U.; VanEpps, J.S.; Yau, S.-T. Culture-free bacterial detection and identification from blood with rapid, phenotypic, antibiotic susceptibility testing. Sci. Rep. 2018, 8, 3416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kittel, M.; Findeisen, P.; Ghebremedhin, B.; Miethke, T.; Grundt, A.; Ahmad-Nejad, P.; Neumaier, M. Rapid susceptibility testing of multi-drug resistant Escherichia coli and Klebsiella by glucose metabolization monitoring. Clin. Chem. Lab. Med. 2019, 57, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.-Y.; Wang, J.-C.; Chuang, H.-S. Rapid Bead-Based Antimicrobial Susceptibility Testing by Optical Diffusometry. PLoS ONE 2016, 11, e0148864. [Google Scholar] [CrossRef] [PubMed]
- Iriya, R.; Syal, K.; Jing, W.; Mo, M.; Yu, H.; Haydel, S.E.; Wang, S.; Tao, N. Real-time detection of antibiotic activity by measuring nanometer-scale bacterial deformation. J. Biomed. Opt. 2017, 22, 126002–126009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparbier, K.; Schubert, S.; Kostrzewa, M. MBT-ASTRA: A suitable tool for fast antibiotic susceptibility testing? Methods 2016, 104, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Ibarlucea, B.; Rim, T.; Baek, C.K.; de Visser, J.A.G.M.; Baraban, L.; Cuniberti, G. Nanowire sensors monitor bacterial growth kinetics and response to antibiotics. Lab Chip 2017, 17, 4283–4293. [Google Scholar] [CrossRef] [PubMed]
- Kuss, S.; Couto, R.A.D.S.; Evans, R.M.; Lavender, H.; Tang, C.C.; Compton, R.G. Versatile Electrochemical Sensing Platform for Bacteria. Anal. Chem. 2019, 91, 4317–4322. [Google Scholar] [CrossRef] [PubMed]
- Idelevich, E.A.; Hoy, M.; Görlich, D.; Knaack, D.; Grünastel, B.; Peters, G.; Borowski, M.; Becker, K. Rapid Phenotypic Detection of Microbial Resistance in Gram-Positive Bacteria by a Real-Time Laser Scattering Method. Front. Microbiol. 2017, 8, 1064. [Google Scholar] [CrossRef]
- e Silva, D.F.; Silva-Dias, A.; Gomes, R.; Martins-Oliveira, I.; Ramos, M.; Rodrigues, A.; Cantón, R.; Pina-Vaz, C. Evaluation of rapid colistin susceptibility directly from positive blood cultures using a flow cytometry assay. Int. J. Antimicrob. Agents 2019, 54, 820–823. [Google Scholar] [CrossRef]
- Tannert, A.; Grohs, R.; Popp, J.; Neugebauer, U. Phenotypic antibiotic susceptibility testing of pathogenic bacteria using photonic readout methods: Recent achievements and impact. Appl. Microbiol. Biotechnol. 2018, 103, 549–566. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Li, H.; Torab, P.; Mach, K.E.; Craft, D.W.; Thomas, N.J.; Puleo, C.M.; Liao, J.C.; Wang, T.-H.; Wong, P.K. Nanotube assisted microwave electroporation for single cell pathogen identification and antimicrobial susceptibility testing. Nanomed. Nanotechnol. Biol. Med. 2019, 17, 246–253. [Google Scholar] [CrossRef]
- Novelli-Rousseau, A.; Espagnon, I.; Filiputti, D.; Gal, O.; Douet, A.; Mallard, F.; Josso, Q. Culture-free Antibiotic-susceptibility Determination from Single-bacterium Raman Spectra. Sci. Rep. 2018, 8, 3957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitruzzello, G.; Thorpe, S.; Johnson, S.; Evans, A.; Gadêlha, H.; Krauss, T.F. Multiparameter antibiotic resistance detection based on hydrodynamic trapping of individual E. coli. Lab Chip 2019, 19, 1417–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Jing, W.; Iriya, R.; Yang, Y.; Syal, K.; Mo, M.; Grys, T.E.; Haydel, S.E.; Wang, S.; Tao, N. Phenotypic Antimicrobial Susceptibility Testing with Deep Learning Video Microscopy. Anal. Chem. 2018, 90, 6314–6322. [Google Scholar] [CrossRef]
- Correa-Martínez, C.L.; Idelevich, E.A.; Sparbier, K.; Kostrzewa, M.; Becker, K. Rapid Detection of Extended-Spectrum β-Lactamases (ESBL) and AmpC β-Lactamases in Enterobacterales: Development of a Screening Panel Using the MALDI-TOF MS-Based Direct-on-Target Microdroplet Growth Assay. Front. Microbiol. 2019, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köck, R.; Wüllenweber, J.; Horn, D.; Lanckohr, C.; Becker, K.; Idelevich, E.A. Implementation of short incubation MALDI-TOF MS identification from positive blood cultures in routine diagnostics and effects on empiric antimicrobial therapy. Antimicrob. Resist. Infect. Control. 2017, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Kohler, A.; Venturelli, L.; Longo, G.; Dietler, G.; Kasas, S. Nanomotion detection based on atomic force microscopy cantilevers. Cell Surf. 2019, 5, 100021. [Google Scholar] [CrossRef]
- Longo, G.; Alonso-Sarduy, L.; Rio, L.M.; Bizzini, A.; Trampuz, A.; Notz, J.; Dietler, G.; Kasas, S. Rapid detection of bacterial resistance to antibiotics using AFM cantilevers as nanomechanical sensors. Nat. Nanotechnol. 2013, 8, 522–526. [Google Scholar] [CrossRef]
- Kasas, S.; Ruggeri, F.S.; Benadiba, C.; Maillard, C.; Stupar, P.; Tournu, H.; Dietler, G.; Longo, G. Detecting nanoscale vibrations as signature of life. Proc. Natl. Acad. Sci. USA 2014, 112, 378–381. [Google Scholar] [CrossRef] [Green Version]
- Villalba, M.I.; Stupar, P.; Chomicki, W.; Bertacchi, M.; Dietler, G.; Arnal, L.; Vela, M.E.; Yantorno, O.; Kasas, S. Nanomotion Detection Method for Testing Antibiotic Resistance and Susceptibility of Slow-Growing Bacteria. Small 2017, 14. [Google Scholar] [CrossRef]
- Venturelli, L.; Kohler, A.; Stupar, P.; Villalba, M.I.; Kalauzi, A.; Radotic, K.; Bertacchi, M.; Dinarelli, S.; Girasole, M.; Pešić, M.; et al. A perspective view on the nanomotion detection of living organisms and its features. J. Mol. Recognit. 2020, 33, e2849. [Google Scholar] [CrossRef]
- Kasas, S.; Malovichko, A.; Villalba, M.; Vela, M.; Yantorno, O.; Willaert, R. Nanomotion Detection-Based Rapid Antibiotic Susceptibility Testing. Antibiotics 2021, 10, 287. [Google Scholar] [CrossRef]
- Syal, K.; Iriya, R.; Yang, Y.; Yu, H.; Wang, S.; Haydel, S.E.; Chen, H.-Y.; Tao, N. Antimicrobial Susceptibility Test with Plasmonic Imaging and Tracking of Single Bacterial Motions on Nanometer Scale. ACS Nano 2015, 10, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Syal, K.; Shen, S.; Yang, Y.; Wang, S.; Haydel, S.E.; Tao, N. Rapid Antibiotic Susceptibility Testing of Uropathogenic E. coli by Tracking Submicron Scale Motion of Single Bacterial Cells. ACS Sens. 2017, 2, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Bermingham, C.R.; Murillo, I.; Payot, A.D.; Balram, K.C.; Kloucek, M.B.; Hanna, S.; Antognozzi, M. Imaging of sub-cellular fluctuations provides a rapid way to observe bacterial viability and response to antibiotics. bioRxiv 2018, 460139. [Google Scholar] [CrossRef] [Green Version]
- Leonard, H.; Halachmi, S.; Ben-Dov, N.; Nativ, O.; Segal, E. Unraveling Antimicrobial Susceptibility of Bacterial Networks on Micropillar Architectures Using Intrinsic Phase-Shift Spectroscopy. ACS Nano 2017, 11, 6167–6177. [Google Scholar] [CrossRef] [PubMed]
- Willaert, R.G.; Boer, P.V.; Malovichko, A.; Alioscha-Perez, M.; Radotić, K.; Bartolić, D.; Kalauzi, A.; Villalba, M.I.; Sanglard, D.; Dietler, G.; et al. Single yeast cell nanomotions correlate with cellular activity. Sci. Adv. 2020, 6, eaba3139. [Google Scholar] [CrossRef] [PubMed]
- Radonicic, V.; Yvanoff, C.; Villalba, M.I.; Kasas, S.; Willaert, R.G. The Dynamics of Single-Cell Nanomotion Behaviour of Saccharomyces cerevisiae in a Microfluidic Chip for Rapid Antifungal Susceptibility Testing. Fermentation 2022, 8, 195. [Google Scholar] [CrossRef]
- Marchetti, O.; Majcherczyk, P.A.; Glauser, M.P.; Bille, J.; Moreillon, P.; Sanglard, D. Sensitive Bioassay for Determination of Fluconazole Concentrations in Plasma Using a Candida albicans Mutant Hypersusceptible to Azoles. Antimicrob. Agents Chemother. 2001, 45, 696–700. [Google Scholar] [CrossRef] [Green Version]
- A Fonzi, W.; Irwin, M.Y. Isogenic strain construction and gene mapping in Candida albicans. Genetics 1993, 134, 717–728. [Google Scholar] [CrossRef]
- Davis, R.H.; Hunt, T.P. Modeling and Measurement of Yeast Flocculation. Biotechnol. Prog. 1986, 2, 91–97. [Google Scholar] [CrossRef]
- Fontana, A.; Bore, C.; Ghommidh, C.; Guiraud, J.P. Structure and Sucrose Hydrolysis Activity of Saccharorn yces cerevisiae Aggregates. Biotechnol. Bioeng. 1992, 40, 475–482. [Google Scholar] [CrossRef]
- van Hamersveld, E.H.; van der Lans, R.G.J.M.; Luyben, K.C.A.M. Quantification of Brewers’ Yeast Flocculation in a Stirred Tank: Effect of Physical Parameters on Flocculation; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1997. [Google Scholar]
- Haddad, S.A.; Lindegren, C.C. A Method for Determining the Weight of an Individual Yeast Cell. Appl. Environ. Microbiol. 1953, 1, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Guizar-Sicairos, M.; Thurman, S.T.; Fienup, J.R. Efficient subpixel image registration algorithms. Opt. Lett. 2008, 33, 156–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwolek-Mirek, M.; Zadrag-Tecza, R. Comparison of methods used for assessing the viability and vitality of yeast cells. FEMS Yeast Res. 2014, 14, 1068–1079. [Google Scholar] [CrossRef] [Green Version]
- Suwunnakorn, S.; Wakabayashi, H.; Kordalewska, M.; Perlin, D.S.; Rustchenko, E. FKS2 and FKS3 Genes of Opportunistic Human Pathogen Candida albicans Influence Echinocandin Susceptibility. Antimicrob. Agents Chemother. 2018, 62, e02299-17. [Google Scholar] [CrossRef] [Green Version]
- Sanglard, D.; Ischer, F.; Marchetti, O.; Entenza, J.; Bille, J. Calcineurin A of Candida albicans: Involvement in antifungal tolerance, cell morphogenesis and virulence. Mol. Microbiol. 2003, 48, 959–976. [Google Scholar] [CrossRef]
- Cordeiro, R.A.; Teixeira, C.E.C.; Brilhante, R.S.N.; Castelo-Branco, D.S.C.M.; Paiva, M.A.N.; Leite, J.J.G.; Lima, D.T.; Monteiro, A.J.; Sidrim, J.J.C.; Rocha, M.F.G. Minimum inhibitory concentrations of amphotericin B, azoles and caspofungin against Candida species are reduced by farnesol. Med. Mycol. 2013, 51, 53–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetti, O.; Moreillon, P.; Entenza, J.M.; Vouillamoz, J.; Glauser, M.P.; Bille, J.; Sanglard, D. Fungicidal Synergism of Fluconazole and Cyclosporine in Candida albicans Is Not Dependent on Multidrug Efflux Transporters Encoded by the CDR1, CDR2, CaMDR1, and FLU1 Genes. Antimicrob. Agents Chemother. 2003, 47, 1565–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rettig, J.R.; Folch, A. Large-Scale Single-Cell Trapping and Imaging Using Microwell Arrays. Anal. Chem. 2005, 77, 5628–5634. [Google Scholar] [CrossRef] [PubMed]
- Hoang, A. Caspofungin acetate: An antifungal agent. Am. J. Health Pharm. 2001, 58, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Shrivastava, M.; Whiteway, M.; Jiang, Y. Candida albicans targets that potentially synergize with fluconazole. Crit. Rev. Microbiol. 2021, 47, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Pristov, K.E.; Ghannoum, M.A. Resistance of Candida to azoles and echinocandins worldwide. Clin. Microbiol. Infect. 2019, 25, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Berman, J.; Krysan, D.J. Drug resistance and tolerance in fungi. Nat. Rev. Genet. 2020, 18, 319–331. [Google Scholar] [CrossRef]
- Willaert, R.G. Micro- and Nanoscale Approaches in Antifungal Drug Discovery. Fermentation 2018, 4, 43. [Google Scholar] [CrossRef] [Green Version]
- Morschhäuser, J. The genetic basis of fluconazole resistance development in Candida albicans. Biochim. Biophys. Acta Mol. Basis Dis. 2002, 1587, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Pourakbari, B.; Teymuri, M.; Mahmoudi, S.; Valian, S.K.; Movahedi, Z.; Eshaghi, H.; Mamishi, S. Expression of Major Efflux Pumps in Fluconazole-Resistant Candida albicans. Infect. Disord. Drug Targets 2017, 17, 178–184. [Google Scholar] [CrossRef]
- Calabrese, D.; Bille, J.; Sanglard, D. A novel multidrug efflux transporter gene of the major facilitator superfamily from Candida albicans (FLU1) conferring resistance to fluconazole. Microbiology 2000, 146, 2743–2754. [Google Scholar] [CrossRef] [Green Version]
- Villalba, M.I.; Rossetti, E.; Bonvallat, A.; Yvanoff, C.; Radonicic, V. Simple optical nanomotion method for single-bacterium viability and antibiotic response testing. Proc. Natl. Acad. Sci. USA 2023. [Google Scholar]
- Roslon, I.; Japaridze, A.; Rodenhuis, S.; Hamoen, L.; Ghatkesar, M.; Steeneken, P.; Alijani, F. Microwell-enhanced optical detection of single bacteria. bioRxiv 2023. [Google Scholar] [CrossRef]
- Rosłoń, I.E.; Japaridze, A.; Steeneken, P.G.; Dekker, C.; Alijani, F. Probing nanomotion of single bacteria with graphene drums. Nat. Nanotechnol. 2022, 17, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Starodubtseva, M.N.; Chelnokova, I.A.; Shkliarava, N.M.; Villalba, M.I.; Tapalski, D.V.; Kasas, S.; Willaert, R.G. Modulation of the nanoscale motion rate of Candida albicans by X-rays. Front. Microbiol. 2023, 14, 833. [Google Scholar] [CrossRef]




















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radonicic, V.; Yvanoff, C.; Villalba, M.I.; Devreese, B.; Kasas, S.; Willaert, R.G. Single-Cell Optical Nanomotion of Candida albicans in Microwells for Rapid Antifungal Susceptibility Testing. Fermentation 2023, 9, 365. https://doi.org/10.3390/fermentation9040365
Radonicic V, Yvanoff C, Villalba MI, Devreese B, Kasas S, Willaert RG. Single-Cell Optical Nanomotion of Candida albicans in Microwells for Rapid Antifungal Susceptibility Testing. Fermentation. 2023; 9(4):365. https://doi.org/10.3390/fermentation9040365
Chicago/Turabian StyleRadonicic, Vjera, Charlotte Yvanoff, Maria Ines Villalba, Bart Devreese, Sandor Kasas, and Ronnie G. Willaert. 2023. "Single-Cell Optical Nanomotion of Candida albicans in Microwells for Rapid Antifungal Susceptibility Testing" Fermentation 9, no. 4: 365. https://doi.org/10.3390/fermentation9040365
APA StyleRadonicic, V., Yvanoff, C., Villalba, M. I., Devreese, B., Kasas, S., & Willaert, R. G. (2023). Single-Cell Optical Nanomotion of Candida albicans in Microwells for Rapid Antifungal Susceptibility Testing. Fermentation, 9(4), 365. https://doi.org/10.3390/fermentation9040365