
Initial Study of Fungal Bioconversion of guishe (Agave lechuguilla Residue) Juice for Bioherbicide Activity on Model Seeds
 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Extract Conditioning
2.2. Fungal Strain Growth and Fermentation Conditions
2.3. Analytical Methods
2.3.1. Spectrophotometric Analysis
2.3.2. Phenolic Content by HPLC-ESI-MS
2.4. Germination and Vegetative Growth Effects
2.5. Data Analyses
3. Results
3.1. Guishe Juice Characterization
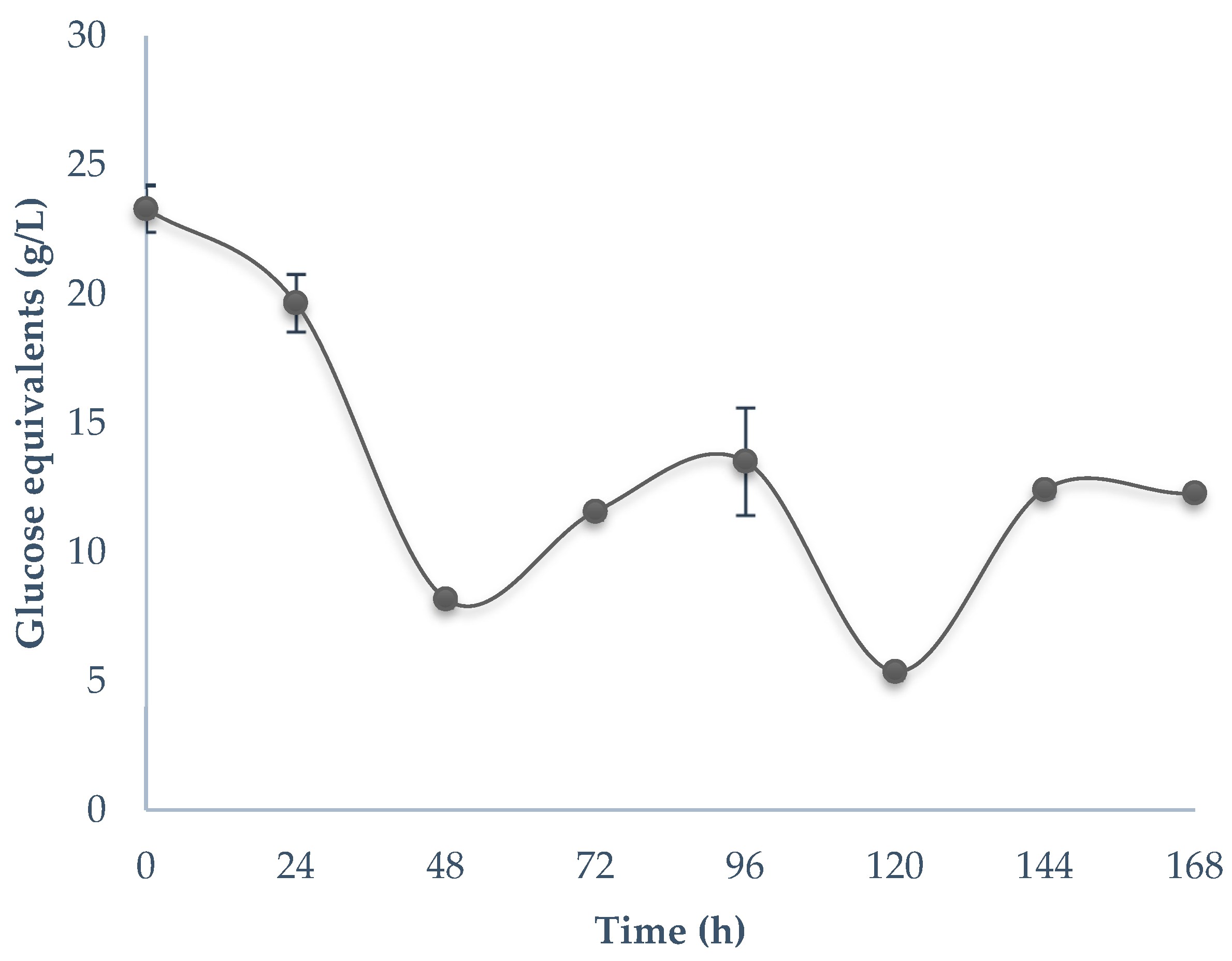
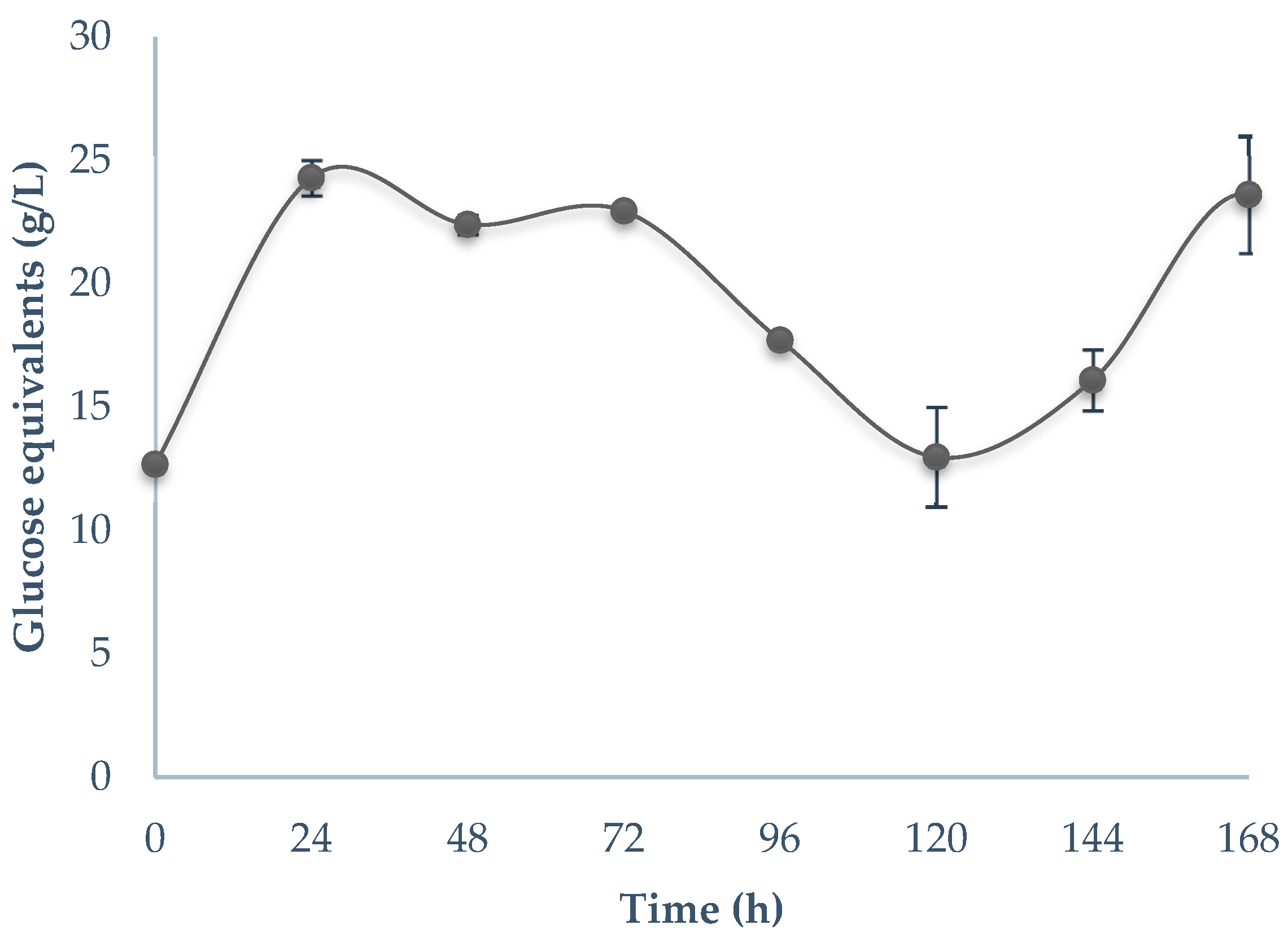
3.2. Bioprocessing of Guishe Juice
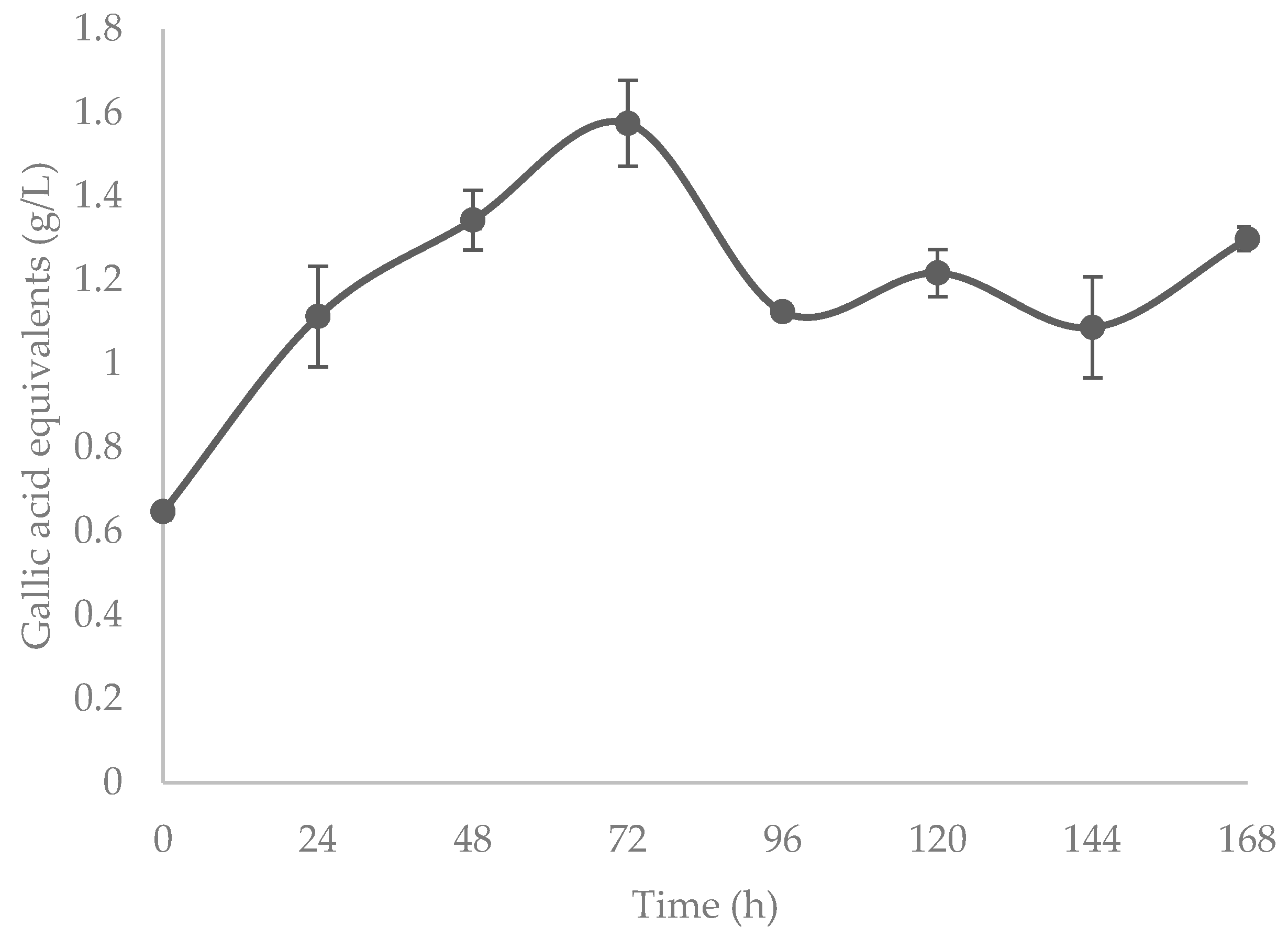
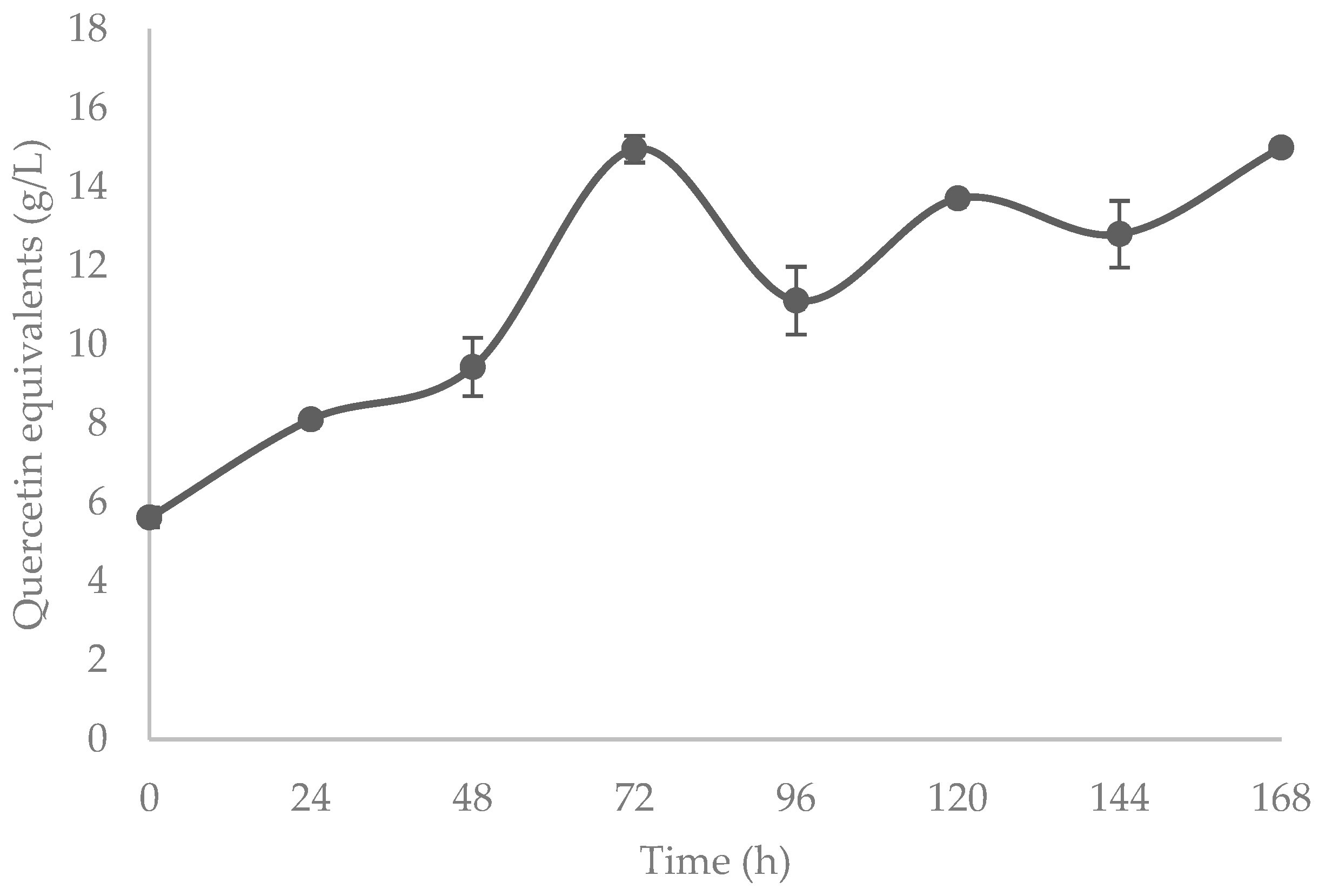
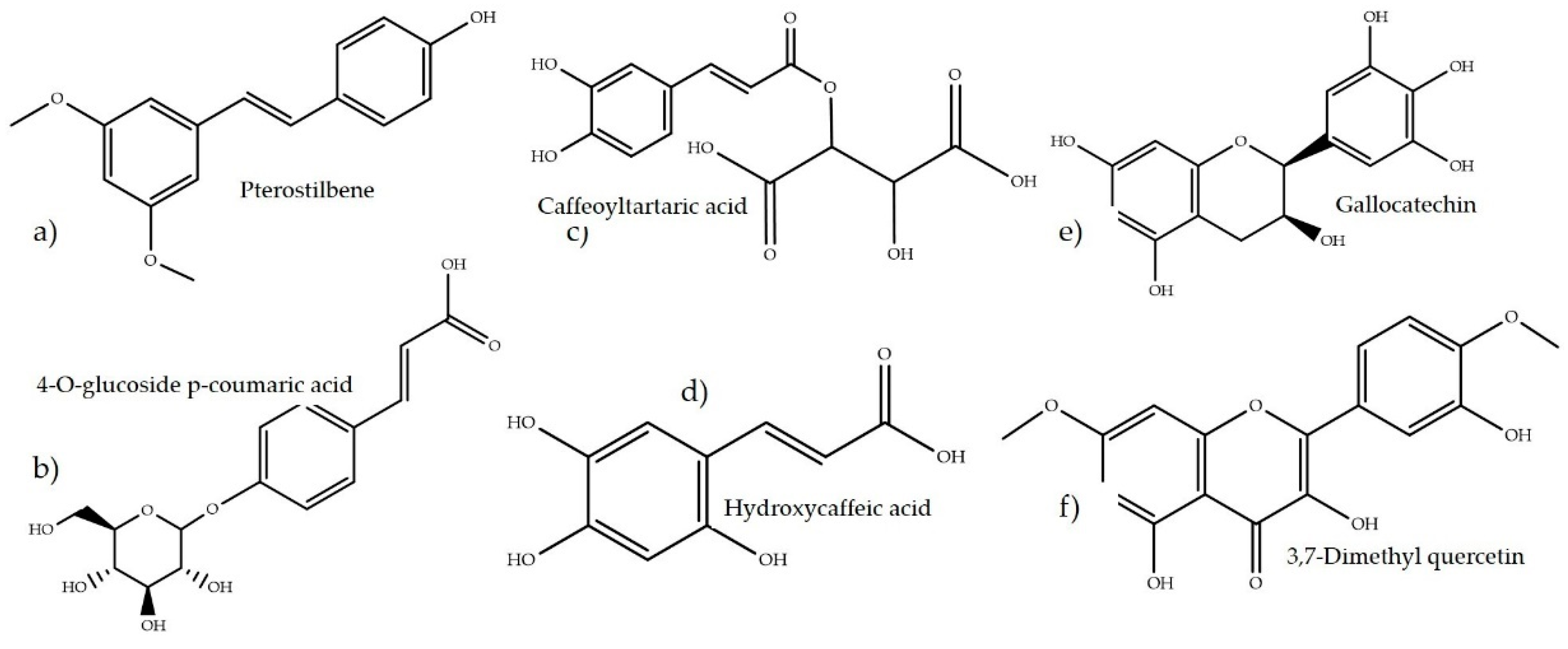
3.3. Flavonoids and Hydrolyzable Phenolics
3.4. Allelopathic Activity
4. Discussion
4.1. Guishe Juice Characterization
4.2. Bioprocessing of Guishe
4.3. Flavonoids and Hydrolyzable Phenolics
4.4. Allelopathic Activity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kong, C.H.; Xuan, T.D.; Khanh, T.D.; Tran, H.D.; Trung, N.T. Allelochemicals and Signaling Chemicals in Plants. Molecules 2019, 24, 2737. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, W.; Ain, Q.; Siddiqui, M.B.; Alharby, H.; Hakeem, K.R. Allelochemicals change macromolecular content of some selected weeds. S. Afr. J. Bot. 2020, 130, 177–184. [Google Scholar] [CrossRef]
- Ullah, H.; Khan, N.; Khan, I.A. Complementing cultural weed control with plant allelopathy: Implications for improved weed management in wheat crop. Acta Ecol. Sin. 2023, 43, 27–33. [Google Scholar] [CrossRef]
- Motmainna, M.; Juraimi, A.S.B.; Uddin, M.K.; Asib, N.B.; Mominul Islam, A.K.M.; Hasan, M. Assessment of allelopathic compounds to develop new natural herbicides: A review. Allelopath. J. 2021, 52, 19–37. [Google Scholar] [CrossRef]
- Meftaul, I.M.; Venkateswarlu, K.; Dharmarajan, R.; Annamalai, P.; Asaduzzaman, M.; Parven, A.; Megharaj, M. Controversies over human health and ecological impacts of glyphosate: Is it to be banned in modern agriculture? Environ. Pollut. 2020, 263, 114372. [Google Scholar] [CrossRef]
- Wang, X.; Lu, Q.; Guo, J.; Ares, I.; Martínez, M.; Martínez-Larrañaga, M.R.; Wang, X.; Anadón, A.; Martínez, M.A. Oxidative Stress and Metabolism: A Mechanistic Insight for Glyphosate Toxicology. Annu. Rev. Pharmacol. Toxicol. 2022, 62, 617–639. [Google Scholar] [CrossRef] [PubMed]
- Anh, L.; Van Quan, N.; Tuan Nghia, L.; Xuan, T.D. Phenolic allelochemicals: Achievements, limitations, and prospective approaches in weed management. Weed Biol. Manag. 2021, 21, 37–67. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy and Allelochemicals of Leucaena leucocephala as an Invasive Plant Species. Plants 2022, 11, 672. [Google Scholar] [CrossRef]
- De la Rosa, L.A.; Moreno-Escamilla, J.O.; Rodrigo-García, J.; Álvarez-Parrilla, E. Phenolic compounds. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Woodhead Publishing: Kidlington, UK, 2019; pp. 253–271. [Google Scholar] [CrossRef]
- Ríos-González, L.J.; Medina-Morales, M.A.; Rodríguez-De la Garza, J.A.; Romero-Galarza, A.; Dávila-Medina, D.; Morales-Martínez, T.K. Comparison of dilute acid pretreatment of agave assisted by microwave versus ultrasound to enhance enzymatic hydrolysis. Bioresour. Technol. 2021, 319, 124099. [Google Scholar] [CrossRef]
- Carmona, J.; Morales Martínez, T.; Mussatto, S.; Castillo Quiroz, D.; Ríos González, L. Chemical, structural and functional properties of lechuguilla (Agave lechuguilla Torr). Rev. Mex. De Cienc. For. 2017, 8, 100–122. [Google Scholar]
- Díaz-Jiménez, L.; Carlos-Hernández, S.; Jasso de Rodríguez, D.; Rodríguez-García, R. Conceptualization of a biorefinery for guishe revalorization. Ind. Crop. Prod. 2019, 138, 111441. [Google Scholar] [CrossRef]
- Peña-Rodríguez, A.; Morreeuw, Z.; García-Luján, J.; Rodríguez-Jaramillo, M.d.C.; Guzmán-Villanueva, L.; Escobedo-Fregoso, C.; Tovar-Ramírez, D.; Reyes, A.G. Evaluation of Agave lechuguilla by-product crude extract as a feed additive for juvenile shrimp Litopenaeus vannamei. Aquac. Res. 2020, 51, 1336–1345. [Google Scholar] [CrossRef]
- Morreeuw, Z.; Escobedo-Fregoso, C.; Ríos-González, L.J.; Castillo-Quiroz, D.; Reyes, A.G. Transcriptome-based metabolic profiling of flavonoids in Agave lechuguilla waste biomass. Plant Sci. 2021, 305, 110748. [Google Scholar] [CrossRef]
- Ladhari, A.; Zarrelli, A.; Di Meo, M.C.; Ghannem, M.; Ben Mimoun, M. Physiological mechanisms and adaptation strategies of Lactuca sativa L. in response to Olea europaea L. and Ficus carica L. allelochemicals. S. Afr. J. Bot. 2022, 147, 106–118. [Google Scholar] [CrossRef]
- Rossi, Y.E.; Vanden Braber, N.L.; Díaz Vergara, L.I.; Montenegro, M.A. Bioactive ingredients obtained from agro-industrial by-products: Recent advances and innovation in micro-and nanoencapsulation. J. Agric. Food Chem. 2021, 69, 15066–15075. [Google Scholar] [CrossRef]
- Shin, H.Y.; Kim, S.M.; Lee, J.H.; Lim, S.T. Solid-state fermentation of black rice bran with Aspergillus awamori and Aspergillus oryzae: Effects on phenolic acid composition and antioxidant activity of bran extracts. Food Chem. 2019, 272, 235–241. [Google Scholar] [CrossRef]
- Sopalun, K.; Laosripaiboon, W.; Wachirachaikarn, A.; Iamtham, S. Potential and chemical composition of bioactive compounds from endophytic fungi associated with thai mangrove plants. S. Afr. J. Bot. 2021, 141, 66–76. [Google Scholar] [CrossRef]
- Hassane, A.M.A.; Taha, T.M.; Awad, M.F.; Mohamed, H.; Melebari, M. Radical scavenging potency, HPLC profiling and phylogenetic analysis of endophytic fungi isolated from selected medicinal plants of Saudi Arabia. Electron. J. Biotechnol. 2022, 58, 37–45. [Google Scholar] [CrossRef]
- Martins, S.; Mussatto, S.I.; Martínez-Avila, G.; Montañez-Saenz, J.; Aguilar, C.N.; Teixeira, J.A. Bioactive phenolic compounds: Production and extraction by solid-state fermentation. A review. Biotechnol. Adv. 2011, 29, 365–373. [Google Scholar] [CrossRef]
- Silva-Beltrán, N.P.; Ruiz-Cruz, S.; Cira-Chávez, L.A.; Estrada-Alvarado, M.I.; Ornelas-Paz, J.D.; López-Mata, M.A.; Del-Toro-Sánchez, C.L.; Ayala-Zavala, J.F.; Márquez-Ríos, E. Tomatidine Contents and Antioxidant and Antimicrobial Activities of Extracts of Tomato Plant. Int. J. Anal. Chem. 2015, 2015, 284071. [Google Scholar] [CrossRef]
- Wong-Paz, J.E.; Contreras-Esquivel, J.C.; Rodríguez-Herrera, R.; Carrillo-Inungaray, M.L.; López, L.I.; Nevárez-Moorillón, V.; Aguilar, C.N. Total phenolic content, in vitro antioxidant activity and chemical composition of plant extracts from semiarid Mexican region. Asian Pac. J. Trop. Med. 2015, 8, 104–111. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, D.A.; Smith, F. Colorimetric Method for determination of sugar and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Estrada-Gil, L.; Contreras-Esquivel, J.C.; Flores-Gallegos, C.; Zugasti-Cruz, A.; Govea-Salas, M.; Mata-Gómez, M.A.; Rodríguez-Herrera, R.; Ascacio-Valdés, J.A. Recovery of Bioactive Ellagitannins by Ultrasound/Microwave-Assisted Extraction from Mexican Rambutan Peel (Nephelium lappaceum L.). Molecules 2022, 27, 1592. [Google Scholar] [CrossRef]
- Chong, J.; Poutaraud, A.; Hugueney, P. Metabolism and roles of stilbenes in plants. Plant Sci. 2009, 177, 143–155. [Google Scholar] [CrossRef]
- Chen, L.; Li, J.; Zhu, Y.; Guo, L.; Ji, R.; Miao, Y.; Guo, L.; Du, H.; Liu, D. Caffeic Acid, an Allelochemical in Artemisia argyi, Inhibits Weed Growth via Suppression of Mitogen-Activated Protein Kinase Signaling Pathway and the Biosynthesis of Gibberellin and Phytoalexin. Front. Plant Sci. 2022, 12, 802198. [Google Scholar] [CrossRef]
- Jiao, X.L.; Bi, X.B.; Gao, W.W. Allelopathic effect of p-coumaric acid on american ginseng and its physiological mechanism. Acta Ecol. Sin. 2015, 35, 3006–3013. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Allelopathy of knotweeds as invasive plants. Plants 2022, 11, 3. [Google Scholar] [CrossRef]
- Pérez-Zavala, M.L.; Hernández-Arzaba, J.C.; Bideshi, D.K.; Barboza-Corona, J.E. Agave: A natural renewable resource with multiple applications. J. Sci. Food Agric. 2020, 100, 5324–5333. [Google Scholar] [CrossRef]
- Das, D.; Ramananda, B.; Selvaraj, R. Optimization of inulinase production by a newly isolated Penicillium amphipolaria strain using solid-state fermentation of hardy sugarcane stems. Biocatal. Agric. Biotechnol. 2020, 30, 101875. [Google Scholar] [CrossRef]
- Brown, P.; Saa, S. Biostimulants in agricultura. Front. Plant Sci. 2015, 6, 671. [Google Scholar] [CrossRef]
- Xiao, J.; Muzashvili, T.S.; Georgiev, M.I. Advances in the biotechnological glycosylation of valuable flavonoids. Biotechnol. Adv. 2014, 32, 1145–1156. [Google Scholar] [CrossRef]
- Lopez-Trujillo, J.; Medina-Morales, M.A.; Sanchez-Flores, A.; Arévalo, C.; Ascacio-Valdes, J.A.; Mellado, M.; Aguilar, C.N.; Aguilera-Carbó, A.F. Solid bioprocess of tarbush (Flourensia cernua) leaves for β-glucosidase production by Aspergillus niger: Initial approach to fiber–glycoside interaction for enzyme induction. 3 Biotech 2017, 7, 271. [Google Scholar] [CrossRef]
- Medina-Morales, M.A.; López-Trujillo, J.; Gómez-Narváez, L.; Mellado, M.; García-Martínez, E.; Ascacio-Valdés, J.A.; Aguilar, C.N.; Aguilera-Carbó, A. Effect of growth conditions on β-glucosidase production using Flourensia cernua leaves in a solid-state fungal bioprocess. 3 Biotech 2017, 7, 355. [Google Scholar] [CrossRef]
- Xue, P.; Liao, W.; Chen, Y.; Xie, J.; Chang, X.; Peng, G.; Huang, Q.; Wang, Y.; Sun, N.; Yu, Q. Release characteristic and mechanism of bound polyphenols from insoluble dietary fiber of navel orange peel via mixed solid-state fermentation with Trichoderma reesei and Aspergillus niger. LWT 2022, 161, 113387. [Google Scholar] [CrossRef]
- Ding, X.; Yu, Y.; Ding, Z. Interaction between active compounds from Rosa roxburghii Tratt and β -glucosidase: Characterization of complexes and binding mechanism. LWT 2022, 165, 113707. [Google Scholar] [CrossRef]
- Contreras-Domínguez, M.; Guyot, S.; Marnet, N.; Le Petit, J.; Perraud-Gaime, I.; Roussos, S.; Augur, C. Degradation of procyanidins by Aspergillus fumigatus: Identification of a novel aromatic ring cleavage product. Biochimie 2006, 88, 1899–1908. [Google Scholar] [CrossRef]
- Cheng, F.; Cheng, Z. Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy. Front. Plant Sci. 2015, 6, 1020. [Google Scholar] [CrossRef]
- Nadeem, M.; Tanveer, A.; Khaliq, A.; Murtaza, G. Supressão do crescimento de plântulas de milho por resíduos de ervade-jacaré (Alternanthera philoxeroides). Planta Daninha 2017, 35, 1–8. [Google Scholar] [CrossRef]
- Sunaina Singh, N.B. Alleviation of allelopathic stress of benzoic acid by indole acetic acid in Solanum lycopersicum. Sci. Hortic. 2015, 192, 211–217. [Google Scholar] [CrossRef]
- Li, Z.H.; Wang, Q.; Ruan, X.; Pan, C.; Jiang, D.A. Phenolics and plant allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef]
- Cruz-Requena, M.; Aguilera-Carbó, A.F.; Mellado, M.; García-Martínez, E.; De la Cruz, R.; Medina-Morales, M.A. Production of β-glucosidase in solid state cultures and its application in the food industry. In Handbook of Research on Food Science and Technology; Volume 2: Food Biotechnology and Microbiology; Apple Academics; Taylor and Francis Group: Waretown NJ, USA, 2019; pp. 59–84. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds of Interest | g/L |
|---|---|
| Total sugars | 23.3 ± 0.90 |
| Reducing sugars | 12.67 ± 0.170 |
| Hydrolyzable polyphenols | 0.64 ± 0.014 |
| Flavonoids | 5.62 ± 0.024 |
| Mass (m/z) | Compound | Family | 1 RT (min) |
|---|---|---|---|
| 254.9 | Pterostilbene | Stilbenes | 14.96 |
| 194.9 | Hydroxycaffeic acid | Hydroxycinnamic acids | 21.183 |
| 325 | 4-O-glucoside p-coumaric acid | Hydroxycinnamic acids | 54.594 |
| 311.1 | Caffeoyltartaric acid | Hydroxycinnamic acids | 55.685 |
| Mass (m/z) | Compound | Family | 1 RT (min) |
|---|---|---|---|
| 24 h | |||
| 304.7 | (+)-Gallocatechin | Catechins | 3.466 |
| 254.9 | Pterostilbene | Stilbenes | 14.77 |
| 195 | Hydroxycaffeic acid | Hydroxycinnamic acids | 20.542 |
| 917 | 3,7-Dimethyl quercetin | Methoxyflavones | 55.706 |
| 325 | 4-O-glucoside p-coumaric acid | Hydroxycinnamic acids | |
| 48 h | |||
| 304.7 | (+)-Gallocatechin | Catechins | 3.336 |
| 254.9 | Pterostilbene | Stilbenes | 14.859 |
| 194.9 | Hydroxycaffeic acid | Hydroxycinnamic acids | 21.513 |
| 325 | 4-O-glucoside p-coumaric acid | Hydroxycinnamic acids | 55.948 |
| 72 h | |||
| 304.7 | (+)-Gallocatechin | Catechins | 3.39 |
| 254.9 | Pterostilbene | Stilbenes | 14.249 |
| 194.9 | Hydroxycaffeic acid | Hydroxycinnamic acids | 20.825 |
| 325 | 4-O-glucoside p-coumaric acid | Hydroxycinnamic acids | 55.73 |
| 96 h | |||
| 304.7 | (+)-Gallocatechin | Catechins | 3.504 |
| 254.9 | Pterostilbene | Stilbenes | 14.515 |
| 195 | Hydroxycaffeic acid | Hydroxycinnamic acids | 21.071 |
| Treatment | 24 h | 48 h | 72 h | |||
|---|---|---|---|---|---|---|
| Control treatment | ||||||
| Water | 70 | abc | 0 | a | 0 | b |
| Glyphosate | 70 ± 5.77 | ab | 0 | a | 0 | b |
| Bioprocesses guishe juice | ||||||
| 0.50% | 60 ± 17.32 | bc | 3.33 ± 5.77 | a | 0.00 | b |
| 1.00% | 36.67 ± 20.82 | c | 3.33 ± 5.77 | a | 0.00 | b |
| 2.00% | 36.67 ± 15.28 | c | 3.33 ± 5.77 | a | 0.00 | b |
| 100% | 100 | a | 96.67 ± 5.77 | a | 96.67 ± 5.77 | a |
| Treatment | 24 h | 48 h | 72 h | |||
|---|---|---|---|---|---|---|
| Control treatment | ||||||
| Water | 30 | a | 46.67 ± 5.77 | abc | 46.67 ± 5.77 | ab |
| Glyphosate | 63.33 | a | 63.33 ± 5.77 | ab | 66.67 ± 5.77 | ab |
| Bioprocess guishe juice | ||||||
| 0.50% | 100 | a | 33.33 ± 15.28 | bc | 26.67 ± 20.82 | b |
| 1.00% | 100 | a | 32.00 ± 11.55 | bc | 26.67 ± 15.28 | b |
| 2.00% | 100 | a | 30.00 ± 20.00 | c | 16.67 ± 15.28 | b |
| 100% | 100 | a | 86.67 ± 11.55 | a | 76.67 ± 11.55 | a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez Robles, J.H.; Luna Enríquez, C.F.; Reyes, A.G.; Cruz Requena, M.; Ríos González, L.J.; Morales Martínez, T.K.; Ascacio Valdés, J.A.; Medina Morales, M.A. Initial Study of Fungal Bioconversion of guishe (Agave lechuguilla Residue) Juice for Bioherbicide Activity on Model Seeds. Fermentation 2023, 9, 421. https://doi.org/10.3390/fermentation9050421
Sánchez Robles JH, Luna Enríquez CF, Reyes AG, Cruz Requena M, Ríos González LJ, Morales Martínez TK, Ascacio Valdés JA, Medina Morales MA. Initial Study of Fungal Bioconversion of guishe (Agave lechuguilla Residue) Juice for Bioherbicide Activity on Model Seeds. Fermentation. 2023; 9(5):421. https://doi.org/10.3390/fermentation9050421
Chicago/Turabian StyleSánchez Robles, José Humberto, Cristina Fernanda Luna Enríquez, Ana G. Reyes, Marisol Cruz Requena, Leopoldo J. Ríos González, Thelma K. Morales Martínez, Juan A. Ascacio Valdés, and Miguel A. Medina Morales. 2023. "Initial Study of Fungal Bioconversion of guishe (Agave lechuguilla Residue) Juice for Bioherbicide Activity on Model Seeds" Fermentation 9, no. 5: 421. https://doi.org/10.3390/fermentation9050421
APA StyleSánchez Robles, J. H., Luna Enríquez, C. F., Reyes, A. G., Cruz Requena, M., Ríos González, L. J., Morales Martínez, T. K., Ascacio Valdés, J. A., & Medina Morales, M. A. (2023). Initial Study of Fungal Bioconversion of guishe (Agave lechuguilla Residue) Juice for Bioherbicide Activity on Model Seeds. Fermentation, 9(5), 421. https://doi.org/10.3390/fermentation9050421