
Mesophilic, Anaerobic Digestion in a Full-Scale, Commercial Biogas Reactor Kills Seeds More Efficiently than Lab-Scale Systems


Abstract
:1. Introduction
2. Materials and Methods
2.1. Species and Seed Collection
2.2. Treatments
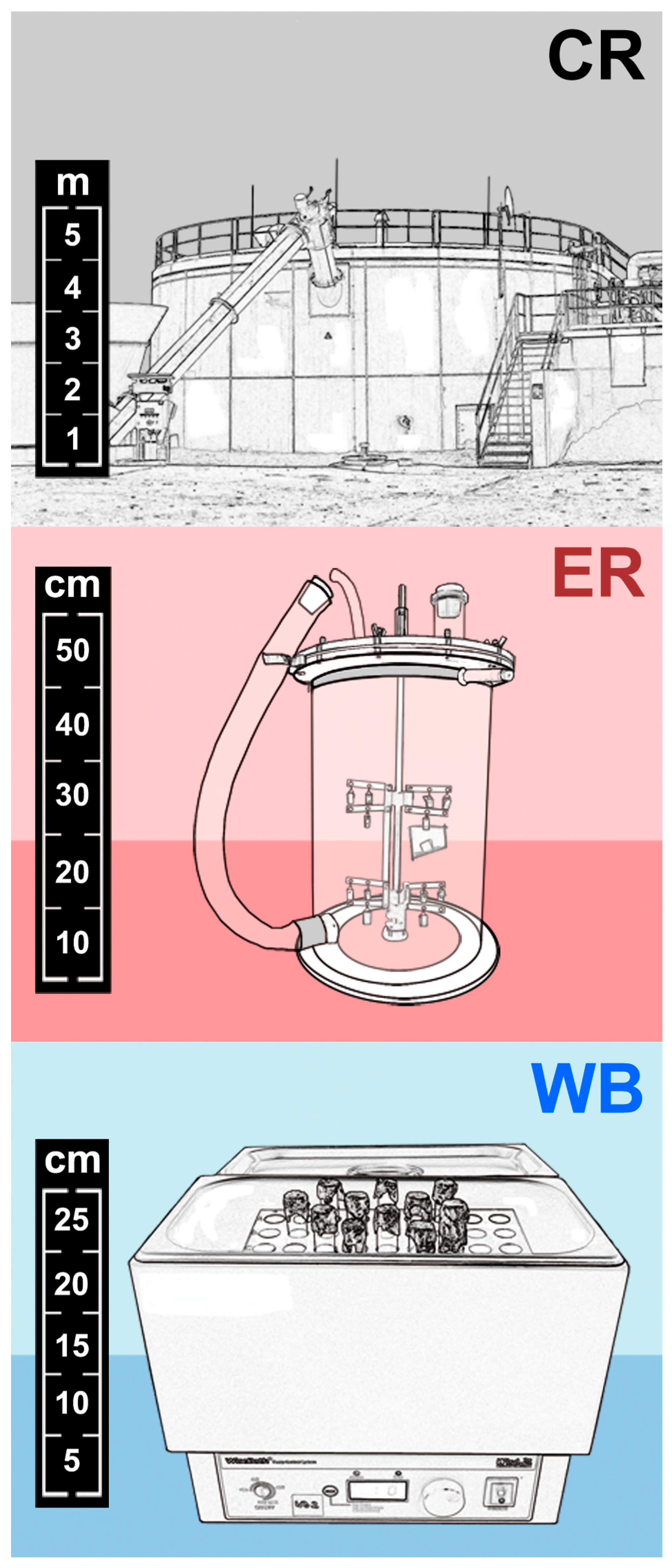
2.2.1. Anaerobic Digestion in a Commercial Biogas Reactor
2.2.2. Anaerobic Digestion in Experimental Biogas Reactors
2.2.3. Buffer Solution in a Water Bath
2.3. Seed Viability
2.4. Data Analyses
- V(t): proportion of viable seeds as a function of the time of exposure in AD (t);
- Vmax: maximum proportion of viable seeds (upper asymptote);
- SLP: parameter proportional to the slope of V(t) in the inflection point;
- MIT (median inactivation time): the time after which V(t) reaches 50% of Vmax.
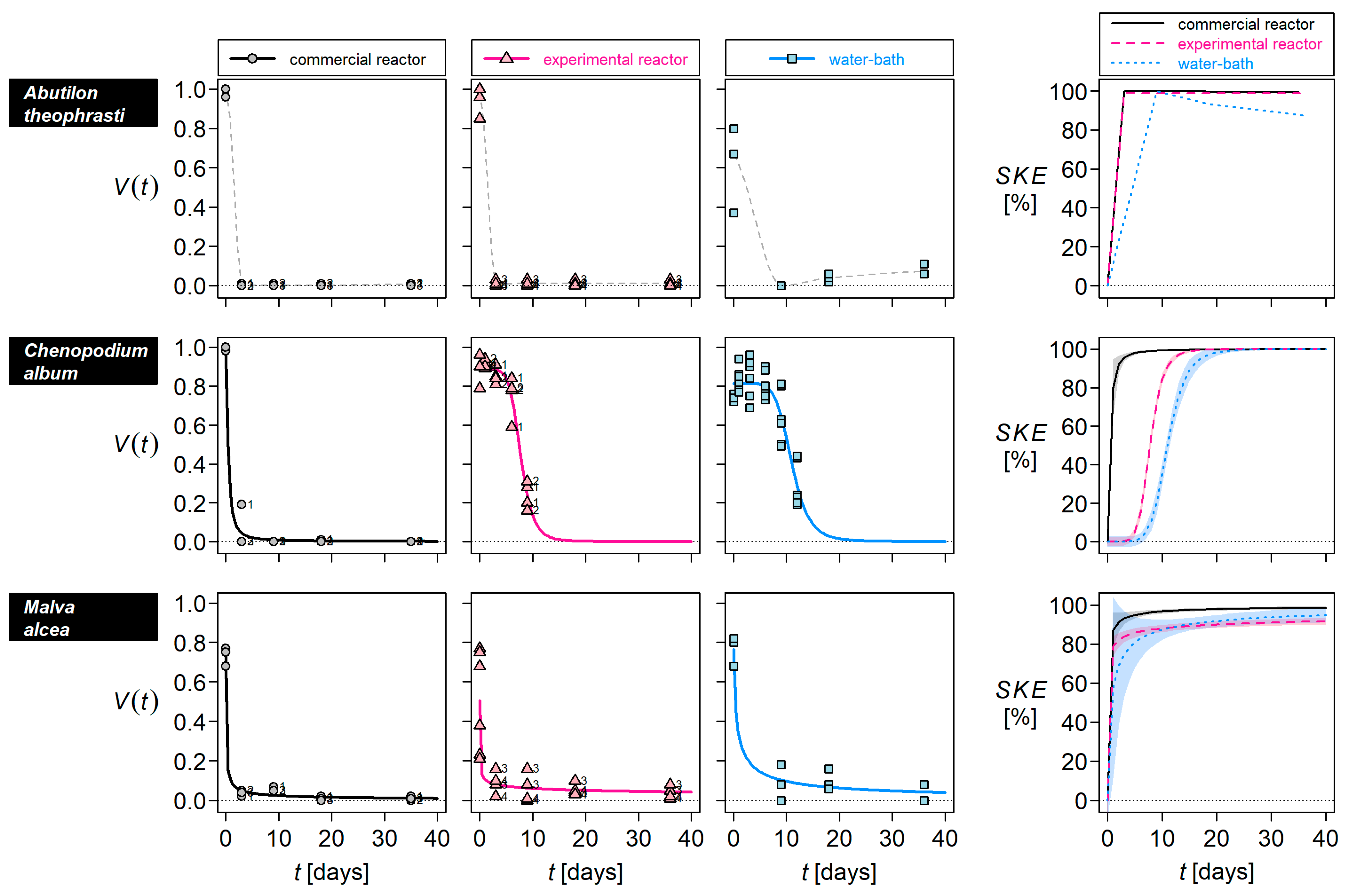
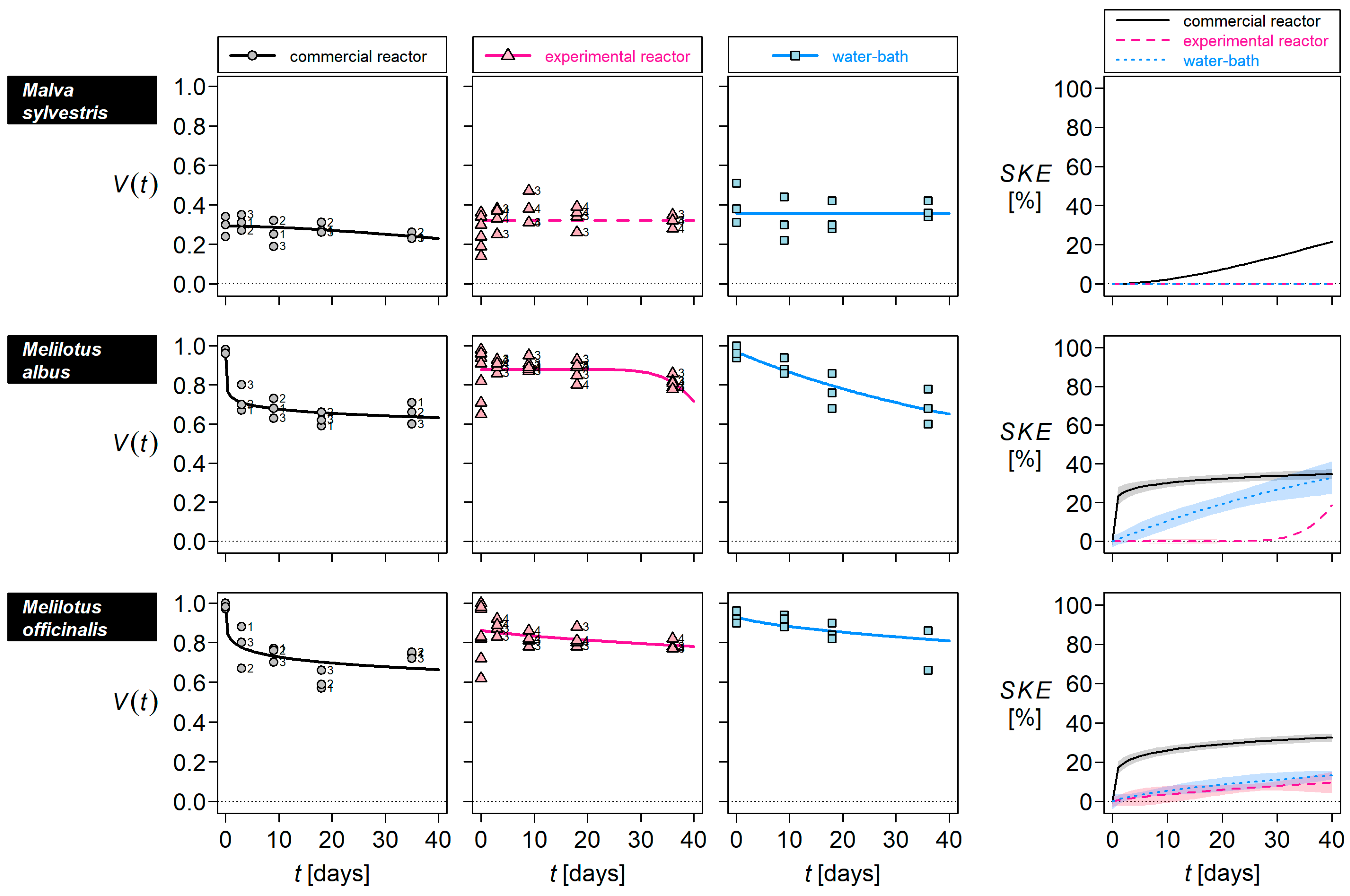
3. Results
4. Discussion
4.1. Seed-Killing Efficacy of the Commercial Reactor
4.2. Factors Inactivating Seeds
4.3. Estimating Seed Survival in Commercial Reactors
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Treatment | Exposure Time [Days] | A. theophrasti | C. album | M. alcea | M. sylvestris | M. albus | M. officinalis |
| CR | 0 | 3 # | 3 # | 3 # | 3 # | 3 # | 3 # |
| 3 | 3 # | 3 # | 3 # | 3 # | 3 # | 3 # | |
| 9 | 3 # | 3 # | 3 # | 3 # | 3 # | 3 # | |
| 18 | 3 ## | 3 ## | 3 ## | 3 ## | 3 ## | 3 ## | |
| 35 | 3 ### | 3 ### | 3 ### | 3 ### | 3 ### | 3 ### | |
| ER | 0 | 4 # | 3 # | 6 # | 6 # | 9 # | 9 # |
| 1 | - | 4 # | - | - | - | - | |
| 3 | 4 # | 4 # | 4 # | 4 # | 6 # | 4 # | |
| 6 | - | 4 ## | - | - | - | - | |
| 9 | 4 # | 4 ### | 4 # | 4 # | 6 # | 4 # | |
| 18 | 4 ## | - | 4 ## | 4 ## | 6 ## | 4 ## | |
| 36 | 4 ### | - | 4 ### | 4 ### | 6 ### | 4 ### | |
| WB | 0 | 3 + | 5 + | 3 + | 3 + | 3 + | 3 + |
| 1 | - | 6 + | - | - | - | - | |
| 3 | - | 6 + | - | - | - | - | |
| 6 | - | 6 + | - | - | - | - | |
| 9 | 3 + | 6 + | 3 + | 3 + | 3 + | 3 + | |
| 12 | - | 6 + | - | - | - | - | |
| 18 | 3 + | - | 3 + | 3 + | 3 + | 3 + | |
| 36 | 3 + | - | 3 + | 3 + | 3 + | 3 + |
References
- Monlau, F.; Sambusiti, C.; Ficara, E.; Aboulkas, A.; Barakat, A.; Carrère, H. New opportunities for agricultural digestate valorization: Current situation and perspectives. Energy Environ. Sci. 2015, 8, 2600–2621. [Google Scholar] [CrossRef]
- Mancini, E.; Raggi, A. A review of circularity and sustainability in anaerobic digestion processes. J. Environ. Manag. 2021, 291, 112695. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lee, D.-J. Valorization of anaerobic digestion digestate: A prospect review. Bioresour. Technol. 2021, 323, 124626. [Google Scholar] [CrossRef]
- Walsh, J.J.; Jones, D.L.; Edwards-Jones, G.; Williams, A.P. Replacing inorganic fertilizer with anaerobic digestate may maintain agricultural productivity at less environmental cost. J. Plant Nutr. Soil Sci. 2012, 175, 840–845. [Google Scholar] [CrossRef]
- Magrí, A. Research Trends on Nutrient Management from Digestates Assessed Using a Bibliometric Approach. Front. Sustain. Food Syst. 2018, 2, 40. [Google Scholar] [CrossRef]
- Meng, X.; Wang, Q.; Lv, Z.; Cai, Y.; Zhu, M.; Li, J.; Ma, X.; Cui, Z.; Ren, L. Novel seedling substrate made by different types of biogas residues: Feasibility, carbon emission reduction and economic benefit potential. Ind. Crops Prod. 2022, 184, 115028. [Google Scholar] [CrossRef]
- Wobiwo, A.F.; Alleluya, V.K.; Emaga, T.H.; Boda, M.; Fokou, E.; Gillet, S.; Deleu, M.; Gerin, P.A. Recovery of fibers and biomethane from banana peduncles biomass through anaerobic digestion. Energy Sustain. Dev. 2017, 37, 60–65. [Google Scholar] [CrossRef]
- Gebhardt, M.; Milwich, M.; Lemmer, A.; Gresser, G.T. Composites based on biogas digestate. Compos. Part C Open Access 2022, 9, 100311. [Google Scholar] [CrossRef]
- Liu, Z.; Liao, W.; Liu, Y. A sustainable biorefinery to convert agricultural residues into value-added chemicals. Biotechnol. Biofuels 2016, 9, 197. [Google Scholar] [CrossRef]
- Weckerle, T.; Ewald, H.; Guth, P.; Knorr, K.-H.; Philipp, B.; Holert, J. Biogas digestate as a sustainable phytosterol source for biotechnological cascade valorization. Microb. Biotechnol. 2023, 16, 337–349. [Google Scholar] [CrossRef]
- Theuerl, S.; Herrmann, C.; Heiermann, M.; Grundmann, P.; Landwehr, N.; Kreidenweis, U.; Prochnow, A. The Future Agricultural Biogas Plant in Germany: A Vision. Energies 2019, 12, 396. [Google Scholar] [CrossRef]
- Fröschle, B.; Heiermann, M.; Lebuhn, M.; Messelhäusser, U.; Plöchl, M. Hygiene and Sanitation in Biogas Plants. In Biogas Science and Technology, 1st ed.; Gübitz, G., Bauer, A., Bochmann, G., Gronauer, A., Weiss, S., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 63–99. ISBN 978-3-319-21992-9. [Google Scholar]
- Lebuhn, M.; Ostertag, J.; Hartel, M.; Knabel, M. Empfehlungen für eine gute fachliche Praxis in landwirtschaftlichen Biogasanlagen aus hygienischer Sicht III; Arbeitsgemeinschaft Landtechnik und landwirtschaftliches Bauwesen in Bayern e.V.; ALB Bayern e.V.: Freising, Germany, 2023. [Google Scholar]
- Engeli, H.; Edelmann, W.; Fuchs, J.; Rottermann, K. Survival of Plant Pathogens and Weed Seeds during Anaerobic Digestion. Wat. Sci. Tech. 1993, 27, 69–76. [Google Scholar] [CrossRef]
- Lorenz, H.; Hellwald, K.-H.; Buchenauer, H. Untersuchungen zur Inaktivierung von Indikatororganismen (Phytohygiene) in anaeroben Kofermentationsanlagen: Teil 1. In Untersuchungen zur Seuchen- und Phytohygiene in Anaerobanlagen (Halb- bzw. großtechnische Anlagen); Knie, A., Haumacher, R., Philipp, W., Martens, W., Böhm, R., Eds.; LUBW: Stuttgart, Germany, 2001; pp. 1–76. [Google Scholar]
- Johansen, A.; Nielsen, H.B.; Hansen, C.M.; Andreasen, C.; Carlsgart, J.; Hauggaard-Nielsen, H.; Roepstorff, A. Survival of weed seeds and animal parasites as affected by anaerobic digestion at meso- and thermophilic conditions. Waste Manag. 2013, 33, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Šarapatka, B.; Holub, M.; Lhotská, M. The effect of farmyard manure anaerobic treatment on weed seed viability. Biol. Agric. Hortic. 1993, 10, 1–8. [Google Scholar] [CrossRef]
- Strauß, G.; Kaplan, T.; Jacobi, T. Keimfähigkeit von Samen verschiedener (gentechnisch veränderter) Nutzpflanzen in Abhängigkeit von Prozessparametern und Verweildauer in einer Biogasanlage. J. Verbr. Lebensm. 2012, 7, 19–25. [Google Scholar] [CrossRef]
- Westerman, P.R.; Hildebrandt, F.; Gerowitt, B. Weed seed survival following ensiling and mesophilic anaerobic digestion in batch reactors. Weed Res. 2012, 52, 286–295. [Google Scholar] [CrossRef]
- Hahn, J.; Westerman, P.R.; de Mol, F.; Heiermann, M.; Gerowitt, B. Viability of Wildflower Seeds after Mesophilic Anaerobic Digestion in Lab-Scale Biogas Reactors. Front. Plant Sci. 2022, 13, 942346. [Google Scholar] [CrossRef]
- Raghu, S.; Anderson, R.C.; Daehler, C.C.; Davis, A.S.; Wiedenmann, R.N.; Simberloff, D.; Mack, R.N. Ecology. Adding biofuels to the invasive species fire? Science 2006, 313, 1742. [Google Scholar] [CrossRef]
- Simberloff, D. Invasion Biologists and the Biofuels Boom: Cassandras or Colleagues. Weed Sci. 2008, 56, 867–872. [Google Scholar] [CrossRef]
- Westerman, P.R.; Gerowitt, B. Weed Seed Survival during Anaerobic Digestion in Biogas Plants. Bot. Rev. 2013, 79, 281–316. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C.; Li, X. Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Spec. Biol. 2000, 15, 139–152. [Google Scholar] [CrossRef]
- Leonhardt, C.; Weinhappel, M.; Gansberger, M.; Brandstetter, A.; Schally, H.; Pfundtner, E. Untersuchungen zur Verbreitungsgefahr von Samenübertragbaren Krankheiten, Unkräutern und Austriebsfähigen Pflanzenteilen mit Fermentationsendprodukten aus Biogasanlagen: Endbericht zum Forschungsprojekt 100296/2. 2010. Available online: http://www.ages.at/uploads/media/100296_Endbericht_biogas_dafne_letztfassung.pdf (accessed on 11 December 2012).
- Westerman, P.R.; Heiermann, M.; Pottberg, U.; Rodemann, B.; Gerowitt, B. Weed seed survival during mesophilic anaerobic digestion in biogas plants. Weed Res. 2012, 52, 307–316. [Google Scholar] [CrossRef]
- Hassani, M.; Vallius, E.; Rasi, S.; Sormunen, K. Risk of Invasive Lupinus polyphyllus Seed Survival in Biomass Treatment Processes. Diversity 2021, 13, 264. [Google Scholar] [CrossRef]
- O’Connor, S.; Ehimen, E.; Pillai, S.C.; Black, A.; Tormey, D.; Bartlett, J. Biogas production from small-scale anaerobic digestion plants on European farms. Renew. Sustain. Energy Rev. 2021, 139, 110580. [Google Scholar] [CrossRef]
- Westerik, M.; Kleizen, R. Onderzoek Sanitatie Tijdens Anaërobe Vergisting Ter Bestrijding Van Onkruidzaden En Ziektekiemen; HoSt: Hengelo, The Netherlands, 2006. [Google Scholar]
- Eckford, R.E.; Newman, J.C.; Li, X.; Watson, P.R. Thermophilic anaerobic digestion of cattle manure reduces seed viability for four weed species. Int. J. Agric. Biol. Eng. 2012, 5, 71–75. [Google Scholar]
- Schrade, S.; Oechsner, H.; Pekrun, C.; Claupein, W. Einfluss des Biogasprozesses auf die Keimfähigkeit von Samen. Landtechnik 2003, 58, 90–91. [Google Scholar] [CrossRef]
- Zhou, L.; Hülsemann, B.; Merkle, W.; Guo, J.; Dong, R.; Piepho, H.-P.; Gerhards, R.; Müller, J.; Oechsner, H. Influence of Anaerobic Digestion Processes on the Germination of Weed Seeds. Gesunde Pflanzen 2020, 72, 181–194. [Google Scholar] [CrossRef]
- Katovich, E.J.; Becker, R.L.; Doll, J. Weed Seed Survival in Anaerobic Digesters; Environmental Impacts and Economic Comparison of Alternative Dairy Systems. 2004. Available online: www.mnproject.org (accessed on 3 February 2020).
- Marcinisyn, E.; Peitzmeier, M.; Heckmann, J. Überprüfung der phyto- und seuchenhygienischen Unbedenklichkeit von Vergärungsrückständen aus der anaeroben Behandlung von Bioabfällen: TV 3—Praxisuntersuchungen. Abschlussbericht, FuE-Vorhaben FKZ 200 33 331; University of Hohenheim: Hohenheim, Germany, 2004. [Google Scholar]
- Baute, K.A.; Robinson, D.E.; van Eerd, L.L.; Edson, M.; Sikkema, P.H.; Gilroyed, B.H. Survival of seeds from perennial biomass species during commercial-scale anaerobic digestion. Weed Res. 2016, 56, 258–266. [Google Scholar] [CrossRef]
- Tanke, A.; Müller, J.; de Mol, F. Seed Viability of Heracleum mantegazzianum (Apiaceae) Is Quickly Reduced at Temperatures Prevailing in Biogas Plants. Agronomy 2019, 9, 332. [Google Scholar] [CrossRef]
- Heiermann, M.; Plogsties, V. Schlussbericht “Wildpflanzen-Samen in der Biogas-Prozesskette—Eintrags- und Überlebensrisiko unter dem Einfluss von Prozessparametern”: Teilprojekt 2 (FKZ 22401513). 2018. Available online: https://www.fnr.de/index.php?id=11150&fkz=22401513 (accessed on 16 February 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Ritz, C.; Streibig, J.C. R Package “drc”. Available online: https://cran.r-project.org/web/packages/drc/drc.pdf (accessed on 15 April 2023).
- Ward, A.J.; Hobbs, P.J.; Holliman, P.J.; Jones, D.L. Optimisation of the anaerobic digestion of agricultural resources. Bioresour. Technol. 2008, 99, 7928–7940. [Google Scholar] [CrossRef]
- vTI. Biogasmessprogramm II: 61 Biogasanlagen im Vergleich. Available online: https://edocs.tib.eu/files/e01fb10/62358767X.pdf (accessed on 1 May 2022).
- Westerman, P.R.; Gerowitt, B. The probability of maize biomass contamination with weed seeds. J. Plant Dis. Protect. 2012, 119, 68–73. [Google Scholar] [CrossRef]
- Thompson, K. Seeds and seed banks. New Phytol. 1987, 106, 23–34. [Google Scholar] [CrossRef]
- Harper, J.L. Population Biology of Plants; Academic Press: London, UK, 1977; ISBN 0-12-325850-2. [Google Scholar]
- Federal Ministry of Environment, Nature Conservation, Nuclear Safety and Consumer Protection. Verordnung über die Verwertung von Bioabfällen auf Landwirtschaftlich, Forstwirtschaftlich und Gärtnerisch Genutzten Böden; Bioabfallverordnung—BioAbfV; Federal Ministry of Environment, Nature Conservation, Nuclear Safety and Consumer Protection: Berlin, Germany, 1998.
- Jiang, Y.; Xie, S.H.; Dennehy, C.; Lawlor, P.G.; Hu, Z.H.; Wu, G.X.; Zhan, X.M.; Gardiner, G.E. Inactivation of pathogens in anaerobic digestion systems for converting biowastes to bioenergy: A review. Renew. Sustain. Energy Rev. 2020, 120, 109654. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, Y. Is anaerobic digestion a reliable barrier for deactivation of pathogens in biosludge? Sci. Tot. Environ. 2019, 668, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, R.; Massé, D.I.; Singh, G. A critical review on inhibition of anaerobic digestion process by excess ammonia. Bioresour. Technol. 2013, 143, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.B. The seed microbiome: Origins, interactions, and impacts. Plant Soil 2018, 422, 7–34. [Google Scholar] [CrossRef]
- Wan, X.; Wu, W.; Li, C.; Liu, Y.; Wen, X.; Liao, Y. Soil ammonia volatilization following urea application suppresses root hair formation and reduces seed germination in six wheat varieties. Environ. Exp. Bot. 2016, 132, 130–139. [Google Scholar] [CrossRef]
- Kosegarten, H.; Grolig, F.; Wieneke, J.; Wilson, G.; Hoffmann, B. Differential Ammonia-Elicited Changes of Cytosolic pH in Root Hair Cells of Rice and Maize as Monitored by 2’,7’-bis- (2 -Carboxyethyl)-5 (and -6)-CarboxyfIuorescein- Fluorescence Ratio. Plant Physiol. 1997, 113, 451–461. [Google Scholar] [CrossRef]
- Bremner, J.M. Recent research on problems in the use of urea as a nitrogen fertilizer. In Nitrogen Economy in Tropical Soils; Ahmad, N., Ed.; Springer: Dordrecht, The Netherlands, 1995; pp. 321–329. [Google Scholar]
- Li, Z.-G.; Lu, X.-Q.; Chen, J. Gasotransmitter ammonia accelerates seed germination, seedling growth, and thermotolerance acquirement in maize. Plant Signal. Behav. 2023, 18, 2163338. [Google Scholar] [CrossRef]
- Milotić, T.; Hoffmann, M. How does gut passage impact endozoochorous seed dispersal success? Evidence from a gut environment simulation experiment. Basic Appl. Ecol. 2016, 17, 165–176. [Google Scholar] [CrossRef]
- Chen, M.-H.; Nelson, E.B. Seed-colonizing microbes from municipal biosolids compost suppress Pythium ultimum damping-off on different plant species. Phytopathology 2008, 98, 1012–1018. [Google Scholar] [CrossRef] [PubMed]
- Theuerl, S.; Klang, J.; Heiermann, M.; de Vrieze, J. Marker microbiome clusters are determined by operational parameters and specific key taxa combinations in anaerobic digestion. Bioresour. Technol. 2018, 263, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Hahn, J.; de Mol, F.; Müller, J. Ensiling Reduces Seed Viability: Implications for Weed Management. Front. Agron. 2021, 3, 1–13. [Google Scholar] [CrossRef]



| Parameter | Commercial Reactor | Experimental Reactor | Water Bath | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Run | Min | Mean | Max | Run | Min | Mean | Max | Min | Mean | Max | |
| T | ∅ | 41.3 | 44.6 | 46.1 | ∅ | 40.3 | 42.1 | 43.3 | 41.9 | 42.0 | 42.1 |
| [°C] | 1 | 41.3 | 43.0 | 44.6 | 1 | 41.9 | 42.5 | 42.9 | |||
| 2 | 44.7 | 45.1 | 46.0 | 2 | 42.1 | 42.5 | 42.7 | ||||
| 3 | 45.4 | 45.7 | 46.1 | 3 | 41.1 | 41.5 | 42.0 | ||||
| 4 | 40.3 | 42.0 | 43.3 | ||||||||
| pH | ∅ | 7.7 | 7.7 | 7.8 | ∅ | 7.4 | 7.7 | 8.0 | - | 7.0 | - |
| 1 | - | 7.7 | - | 1 | 7.6 | 7.6 | 7.7 | ||||
| 2 | - | 7.7 | - | 2 | 7.6 | 7.6 | 7.6 | ||||
| 3 | - | 7.8 | - | 3 | 7.4 | 7.7 | 8.0 | ||||
| 4 | 7.7 | 7.8 | 7.9 | ||||||||
| TS | ∅ | 9.0 | 9.2 | 9.7 | ∅ | 5.6 | 6.0 | 6.8 | na | ||
| [%FM] | 1 | - | 9.7 | - | 1 | 6.5 | 6.5 | 6.5 | |||
| 2 | - | 9.0 | - | 2 | 6.5 | 6.6 | 6.8 | ||||
| 3 | - | 9.0 | - | 3 | 5.6 | 5.7 | 5.8 | ||||
| 4 | 5.8 | 6.0 | 6.4 | ||||||||
| VS | ∅ | 84.4 | 84.4 | 84.5 | ∅ | 70.0 | 74.5 | 77.4 | na | ||
| [%TS] | 1 | - | 84.4 | - | 1 | 75.9 | 76.1 | 76.2 | |||
| 2 | - | 84.5 | - | 2 | 76.1 | 76.2 | 76.4 | ||||
| 3 | - | 84.4 | - | 3 | 70.0 | 74.5 | 77.4 | ||||
| 4 | 71.7 | 73.8 | 75.8 | ||||||||
| NH4-N | ∅ | 3.2 | 3.3 | 3.5 | ∅ | 1.5 | 1.8 | 1.9 | na | ||
| [g.L−1] | 1 | - | 3.2 | - | 1 | 1.8 | 1.8 | 1.9 | |||
| 2 | - | 3.3 | - | 2 | 1.8 | 1.8 | 1.9 | ||||
| 3 | - | 3.5 | - | 3 | 1.5 | 1.7 | 1.8 | ||||
| 4 | 1.8 | 1.8 | 1.9 | ||||||||
| AA | ∅ | 0 | 0.1 | 0.2 | ∅ | 0.1 | 0.3 | 0.8 | na | ||
| [g.L−1] | 1 | - | 0.2 | - | 1 | 0.3 | 0.3 | 0.3 | |||
| 2 | - | 0.1 | - | 2 | 0.3 | 0.3 | 0.3 | ||||
| 3 | nd | 3 | 0.1 | 0.3 | 0.8 | ||||||
| 4 | 0.2 | 0.3 | 0.4 | ||||||||
| VFA | ∅ | 0.1 | 0.1 | 0.2 | ∅ | 0.1 | 0.4 | 0.9 | na | ||
| [g.L−1] | 1 | - | 0.2 | - | 1 | 0.3 | 0.3 | 0.3 | |||
| 2 | - | 0.1 | - | 2 | 0.3 | 0.3 | 0.3 | ||||
| 3 | nd | 3 | 0.1 | 0.4 | 0.9 | ||||||
| 4 | 0.3 | 0.4 | 0.7 | ||||||||
| Biogas | na | ∅ | 5.8 | 13.9 | 17.2 | na | |||||
| [lN.d−1] | 1 | 8.0 | 13.3 | 17.0 | |||||||
| 2 | 5.8 | 13.2 | 15.9 | ||||||||
| 3 | 9.9 | 12.1 | 14.0 | ||||||||
| 4 | 8.1 | 14.6 | 17.2 | ||||||||
| CH4 | ∅ | 50.0 | 51.7 | 53.0 | ∅ | 51.7 | 54.7 | 58.2 | na | ||
| [Vol.%] | 1 | 51.0 | 51.5 | 52.0 | 1 | 55.0 | 57.2 | 58.2 | |||
| 2 | 51.0 | 51.9 | 52.0 | 2 | 55.2 | 57.1 | 57.9 | ||||
| 3 | 50.0 | 51.9 | 53.0 | 3 | 54.4 | 55.7 | 56.7 | ||||
| 4 | 51.7 | 53.6 | 56.2 | ||||||||
| EPG | ∅ | 1.2 | 5.8 | 6.0 | na | na | |||||
| [MWhel] | 1 | 5.5 | 5.9 | 6.0 | |||||||
| 2 | 4.7 | 5.9 | 6.0 | ||||||||
| 3 | 1.2 | 5.7 | 6.0 | ||||||||
| Species | Control for | n | V | G/V | ||
|---|---|---|---|---|---|---|
| Abutilon theophrasti | CR | 3 | 0.99 | (0.01) | 0.46 | (0.04) |
| ER | 4 | 0.95 | (0.04) | 0.46 | na | |
| WB | 3 | 0.61 | (0.13) | na | ||
| Chenopodium album | CR | 3 | 0.99 | (0.01) | 1.00 | 0 |
| ER | 3 | 0.88 | (0.05) | 1.00 | 0 | |
| WB | 5 | 0.75 | (0.01) | na | ||
| Malva alcea | CR | 3 | 0.73 | (0.03) | 0.70 | (0.06) |
| ER | 6 | 0.50 | (0.11) | 0.58 | (0.06) | |
| WB | 3 | 0.77 | (0.04) | na | ||
| Malva sylvestris | CR | 3 | 0.29 | (0.03) | 0.11 | (0.01) |
| ER | 6 | 0.26 | (0.04) | 0.11 | (0.02) | |
| WB | 3 | 0.40 | (0.06) | na | ||
| Melilotus albus | CR | 3 | 0.97 | (0.01) | 0.10 | (0.03) |
| ER | 9 | 0.85 | (0.04) | 0.18 | (0.03) | |
| WB | 3 | 0.97 | (0.02) | na | ||
| Melilotus officinalis | CR | 3 | 0.98 | (0.01) | 0.06 | (0.02) |
| ER | 9 | 0.86 | (0.04) | 0.08 | (0.01) | |
| WB | 3 | 0.93 | (0.02) | na | ||
| Species | Treatment | Vmax | SLP | MIT [Days] | DRT [Days] | ||||
|---|---|---|---|---|---|---|---|---|---|
| Abutilon theophrasti x | all | - | - | - | - | ||||
| Chenopodium album | CR | 0.99 (0.01) | a | 1.52 (0.29) | a | 0.40 (0.19) | a | 1.7 (0.4) | a |
| ER | 0.88 (0.01) | b | 6.71 (0.41) | b | 7.75 (0.09) | b | 10.8 (0.2) | b | |
| WB | 0.81 (0.01) | c | 6.66 (0.84) | b | 10.97 (0.20) | c | 15.3 (0.7) | c | |
| Malva alcea | CR | 0.73 (0.03) | a | 0.64 (0.16) | ab | 0.05 (0.07) | a | 1.6 (0.8) | a |
| ER | 0.51 (0.02) | b | 0.29 (0.06) | b | 0.01 (0.01) | a | 20.1 (7.2) | ab | |
| WB | 0.76 (0.04) | a | 0.70 (0.35) | a | 0.64 (1.02) | b | 14.6 (4.1) | b | |
| Malva sylvestris | CR | 0.29 (0.02) | a | 1.77 (2.19) | a | 83.14 (78.65) | a | 287.6 (707.9) | a |
| ER | 0.32 (0.01) | ab | 2.99 (120.08) | a | >365 - | a | >365 - | a | |
| WB | 0.36 (0.02) | b | 2.09 (15.14) | a | >365 - | a | >365 - | a | |
| Melilotus albus | CR | 0.97 (0.01) | a | 0.15 (0.04) | a | >365 - | a | >365 - | a |
| ER | 0.88 (0.01) | b | 9.86 - | ab | 46.45 - | a | 58.0 - | b | |
| WB | 0.97 (0.01) | a | 1.03 (0.26) | b | 80.44 (27.02) | a | >365 - | ab | |
| Melilotus officinalis | CR | 0.98 (0.01) | a | 0.23 (0.03) | b | >365 - | a | >365 - | a |
| ER | 0.86 (0.01) | b | 0.76 (0.74) | b | >365 - | a | >365 - | a | |
| WB | 0.93 (0.02) | c | 0.70 - | b | >365 - | a | >365 - | a | |
| Species | Seed-Killing Efficacy [%] of 36 Days in | ||
|---|---|---|---|
| Commercial Reactor | Experimental Reactor | Water-Bath | |
| Abutilon theophrasti | 99 | 98 | 88 |
| Chenopodium album | 100 | 100 | 100 |
| Malva alcea | 99 | 91 | 94 |
| Malva sylvestris | 19 | 0 | 0 |
| Melilotus albus | 34 | 7 | 30 |
| Melilotus officinalis | 32 | 9 | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hahn, J.; Westerman, P.R.; Gerowitt, B.; Heiermann, M. Mesophilic, Anaerobic Digestion in a Full-Scale, Commercial Biogas Reactor Kills Seeds More Efficiently than Lab-Scale Systems. Fermentation 2023, 9, 481. https://doi.org/10.3390/fermentation9050481
Hahn J, Westerman PR, Gerowitt B, Heiermann M. Mesophilic, Anaerobic Digestion in a Full-Scale, Commercial Biogas Reactor Kills Seeds More Efficiently than Lab-Scale Systems. Fermentation. 2023; 9(5):481. https://doi.org/10.3390/fermentation9050481
Chicago/Turabian StyleHahn, Juliane, Paula Renate Westerman, Bärbel Gerowitt, and Monika Heiermann. 2023. "Mesophilic, Anaerobic Digestion in a Full-Scale, Commercial Biogas Reactor Kills Seeds More Efficiently than Lab-Scale Systems" Fermentation 9, no. 5: 481. https://doi.org/10.3390/fermentation9050481
APA StyleHahn, J., Westerman, P. R., Gerowitt, B., & Heiermann, M. (2023). Mesophilic, Anaerobic Digestion in a Full-Scale, Commercial Biogas Reactor Kills Seeds More Efficiently than Lab-Scale Systems. Fermentation, 9(5), 481. https://doi.org/10.3390/fermentation9050481