


Purification and Identification of EPS Produced by Five Lactic Acid Bacteria and Evaluation of Their Effects on the Texture of Fermented Goat Milk

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of EPS
2.2. Isolation and Purification of EPS from LAB
2.3. Determination of Molecular Weight Distribution using Gel Permeation Chromatography (GPC)
2.4. Determination of Monosaccharide Composition in Polysaccharides
2.5. Structural Characteristics Determination of EPS using Infrared Spectroscopy
2.6. Preparation of Fermented Goat Milk and Sensory Evaluation
2.7. Determination of PH Value of Fermented Goat Milk
2.8. Determination of Acidity of Fermented Goat Milk
2.9. Determination of Texture (Hardness, Consistency, and Viscosity) of Fermented Goat Milk
2.10. Statistical Analysis
3. Results and Discussion
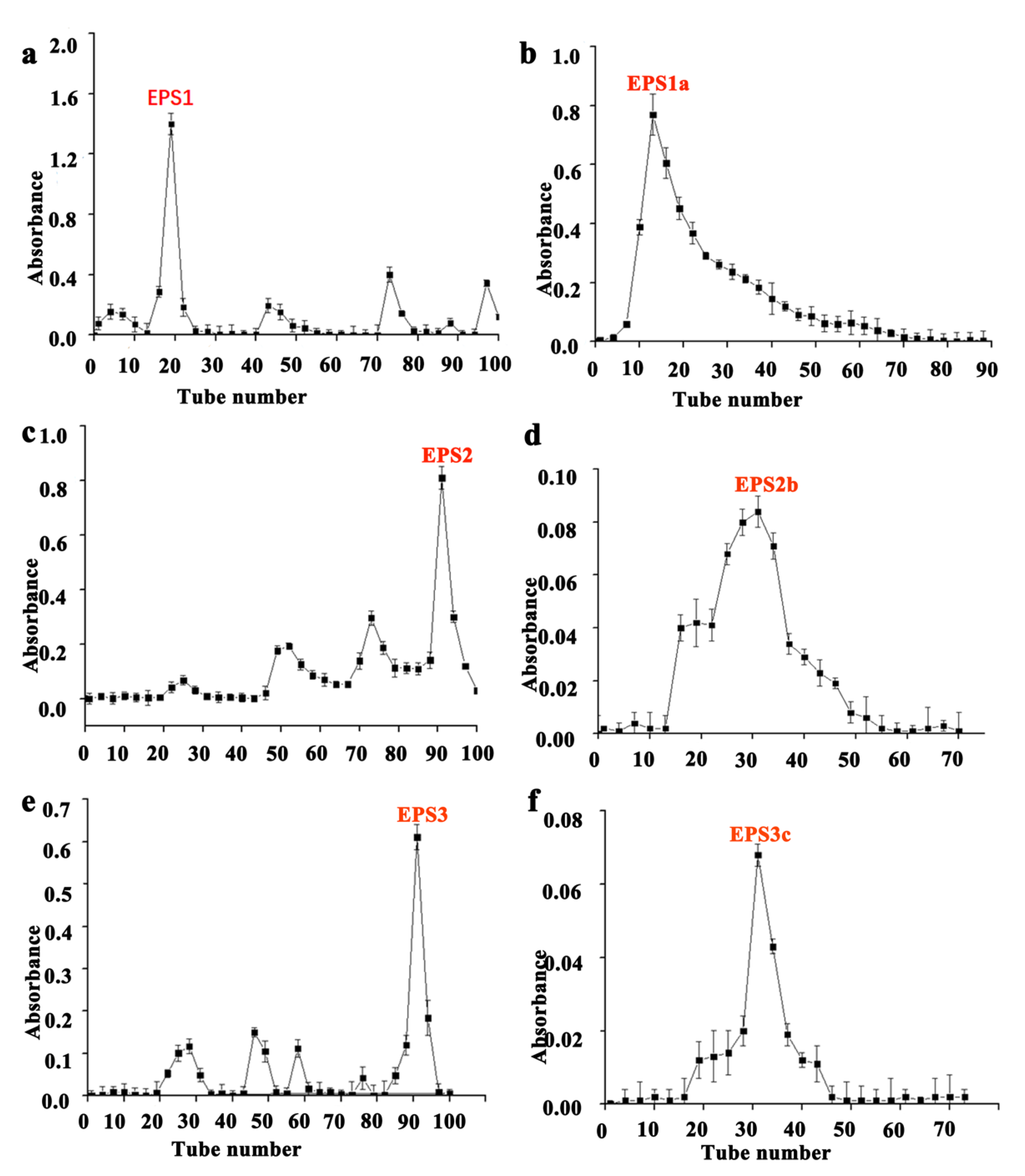
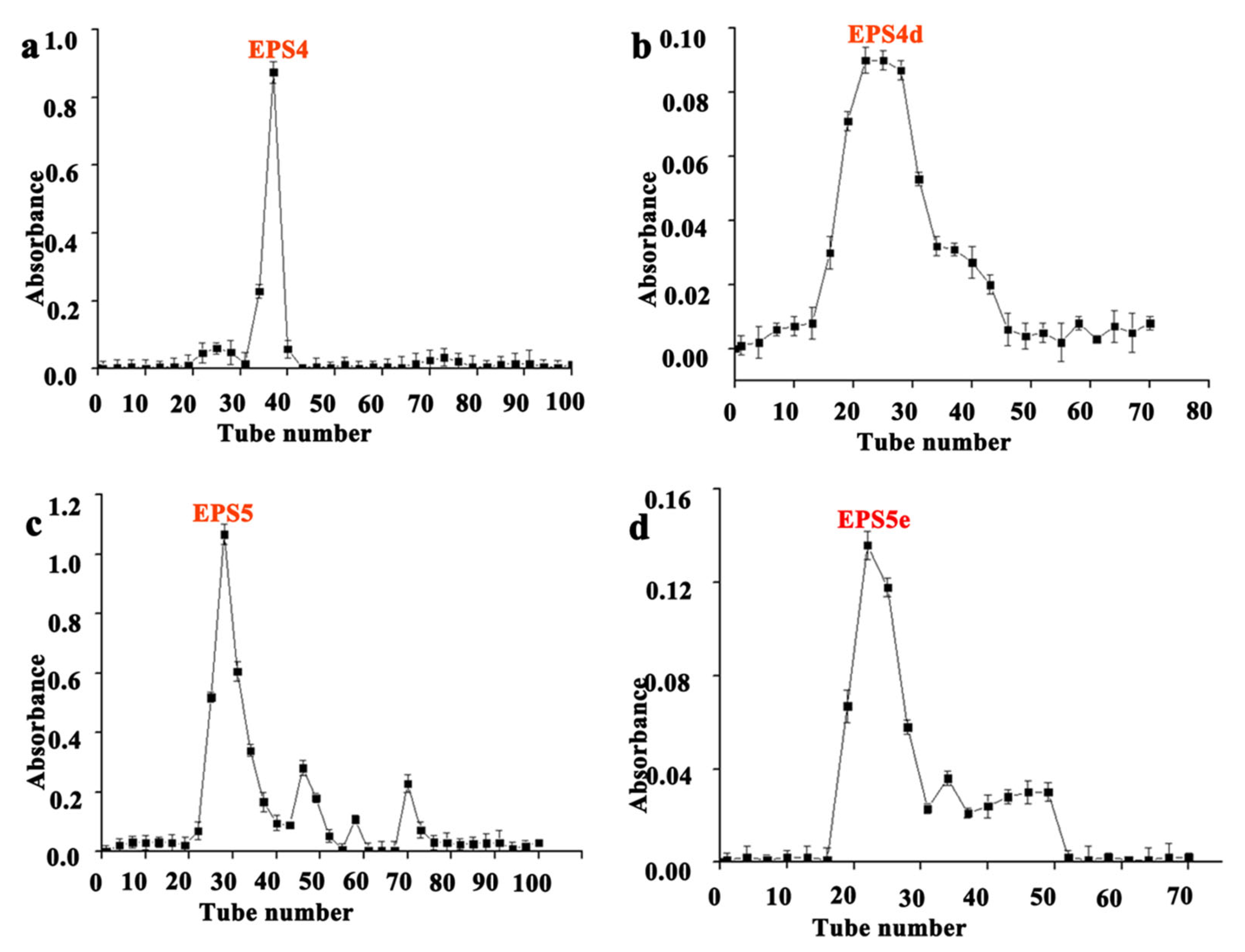
3.1. EPS Isolation and Purification from Five Strains of LAB
3.2. Molecular Weight Determination of EPS of Five Strains of LAB
3.3. Analysis of Monosaccharide Composition of EPS Produced by Five Strains of LAB
3.4. Infrared Spectrum Analysis of EPS Produced by Five Strains of LAB
3.5. Evaluation of the Effect of Five Strains of EPS-Producing LAB on the Texture of Fermented Goat Milk
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Zhao, X.; Yang, Y.; Zhao, A.; Yang, Z. Characterization and bioactivities of an exopolysaccharide produced by Lactobacillus plantarum YW32. Int. J. Biol. Macromol. 2015, 74, 119–126. [Google Scholar] [CrossRef]
- Guo, C.F.; Zhang, L.W.; Li, J.Y.; Zhang, Y.C.; Xue, C.H.; Yi, H.X.; Du, M.; Han, X. Screening of bile salt hydrolase-active lactic acid bacteria for potential cholesterol-lowering probiotic use. In Proceedings of the 2nd International Conference on Advanced Materials Research, Chengdu, China, 7–8 January 2012; pp. 139–146. [Google Scholar]
- Rafter, J. The effects of probiotics on colon cancer development. Nutr. Res. Rev. 2004, 17, 277–284. [Google Scholar] [CrossRef]
- Sanna, S.; van Zuydam, N.R.; Mahajan, A.; Kurilshikov, A.; Vich Vila, A.; Võsa, U.; Mujagic, Z.; Masclee, A.A.; Jonkers, D.M.; Oosting, M. Causal relationships among the gut microbiome, short-chain fatty acids and metabolic diseases. Nat. Genet. 2019, 51, 600–605. [Google Scholar] [CrossRef]
- Donot, F.; Fontana, A.; Baccou, J.; Schorr-Galindo, S. Microbial exopolysaccharides: Main examples of synthesis, excretion, genetics and extraction. Carbohydr. Polym. 2012, 87, 951–962. [Google Scholar] [CrossRef]
- Wang, X.; Shao, C.; Liu, L.; Guo, X.; Xu, Y.; Lü, X. Optimization, partial characterization and antioxidant activity of an exopolysaccharide from Lactobacillus plantarum KX041. Int. J. Biol. Macromol. 2017, 103, 1173–1184. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Luo, J.; Zhang, L.; Wang, W.; Gou, D. MiR-103 controls milk fat accumulation in goat (Capra hircus) mammary gland during lactation. PLoS ONE 2013, 8, e79258. [Google Scholar] [CrossRef]
- Chen, Z.; Luo, J.; Sun, S.; Cao, D.; Shi, H.; Loor, J.J. miR-148a and miR-17–5p synergistically regulate milk TAG synthesis via PPARGC1A and PPARA in goat mammary epithelial cells. RNA Biol. 2017, 14, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Getaneh, G.; Mebrat, A.; Wubie, A.; Kendie, H. Review on goat milk composition and its nutritive value. J. Nutr. Health Sci. 2016, 3, 1–10. [Google Scholar]
- Malau-Aduli, B.; Eduvie, L.; Lakpini, C.; Malau-Aduli, A. Effect of supplementation on the milk yield and composition of Red Sokoto does. In Proceedings of the 26th Annual Conference of the Nigerian Society for Animal Production, Ilorin, Nigeria, 21–25 March 1999; Nigerian Society for Animal Production: Bauchi, Nigeria; Volume 26, pp. 353–356. [Google Scholar]
- Silva, F.A.; de Oliveira, M.E.G.; de Figueirêdo, R.M.F.; Sampaio, K.B.; de Souza, E.L.; de Oliveira, C.E.V.; Pintado, M.M.E.; do Egypto, R.d.C.R. The effect of Isabel grape addition on the physicochemical, microbiological and sensory characteristics of probiotic goat milk yogurt. Food Funct. 2017, 8, 2121–2132. [Google Scholar] [CrossRef]
- Clark, S.; García, M.B.M. A 100-year review: Advances in goat milk research. J. Dairy Sci. 2017, 100, 10026–10044. [Google Scholar] [CrossRef]
- Delgado, K.; Vieira, C.; Dammak, I.; Frasão, B.; Brígida, A.; Costa, M.; Conte-Junior, C. Different ultrasound exposure times influence the physicochemical and microbial quality properties in probiotic goat milk yogurt. Molecules 2020, 25, 4638. [Google Scholar] [CrossRef] [PubMed]
- Zeidan, A.A.; Poulsen, V.K.; Janzen, T.; Buldo, P.; Derkx, P.M.; Øregaard, G.; Neves, A.R. Polysaccharide production by lactic acid bacteria: From genes to industrial applications. FEMS Microbiol. Rev. 2017, 41, S168–S200. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Lai, T.; Yao, M.; Zhang, M.; Yang, Z. Interaction of the Exopolysaccharide from Lactobacillus plantarum YW11 with Casein and Bioactivities of the Polymer Complex. Foods 2021, 10, 1153. [Google Scholar] [CrossRef] [PubMed]
- Ua-Arak, T.; Jakob, F.; Vogel, R.F. Fermentation pH modulates the size distributions and functional properties of Gluconobacter albidus TMW 2.1191 levan. Front. Microbiol. 2017, 8, 807. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.S.; Passos, C.P.; Madureira, P.; Vilanova, M.; Coimbra, M.A. Structure–function relationships of immunostimulatory polysaccharides: A review. Carbohydr. Polym. 2015, 132, 378–396. [Google Scholar] [CrossRef]
- Shangguan, W.; Chen, H.; Li, Y.; Wang, Z.; Guo, H.; Meng, J. Screening and identification of new types of exopolysaccharides-producing lactic acid in the inner mongolia dairy products. Acta Univ. Cibiniensis Ser. E Food Technol. 2019, 23, 75–84. [Google Scholar] [CrossRef]
- Savadogo, A.; Ouattara, C.A.; Savadogo, P.W.; Barro, N.; Ouattara, A.S.; Traoré, A.S. Identification of exopolysaccharides-producing lactic acid bacteria from Burkina Faso fermented milk samples. Afr. J. Biotechnol. 2004, 3, 189–194. [Google Scholar]
- Liu, Q.; Ge, X.; Chen, L.; Cheng, D.; Yun, Z.; Xu, W.; Shao, R. Purification and analysis of the composition and antioxidant activity of polysaccharides from Helicteres angustifolia L. Int. J. Biol. Macromol. 2018, 107, 2262–2268. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.t.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Wang, M.; Geng, L.; Xue, B.; Wang, Z.; Xu, W.; Shu, C.; Zhang, J. Structure characteristics and function of a novel extracellular polysaccharide from Bacillus thuringiensis strain 4D19. Int. J. Biol. Macromol. 2021, 189, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Zhao, M. Purification, characterization and anti-proliferation activities of polysaccharides extracted from Viscum coloratum (Kom.) Nakai. Carbohydr. Polym. 2016, 149, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Sun, F.; Cheng, X.; Li, X.; Mu, H.; Wang, S.; Geng, H.; Duan, J. Preparation and biological activities of an extracellular polysaccharide from Rhodopseudomonas palustris. Int. J. Biol. Macromol. 2019, 131, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Bao, C.; Wang, C. Response surface methodology for optimizing fermentation conditions of goat yogurt with Bifidobacterium bifidum and Lactobacillus casei. Emir. J. Food Agric. 2016, 28, 547–553. [Google Scholar] [CrossRef]
- Ghendov-Moşanu, A.; Sturza, R.; Opriş, O.; Lung, I.; Popescu, L.; Popovici, V.; Soran, M.-L.; Patraş, A. Effect of lipophilic sea buckthorn extract on cream cheese properties. J. Food Sci. Technol. 2020, 57, 628–637. [Google Scholar] [CrossRef] [PubMed]
- Tenea, G.N.; Suárez, J. Probiotic Potential and Technological Properties of Bacteriocinogenic Lactococcus lactis subsp. lactis UTNGt28 from a native Amazonian fruit as a yogurt starter culture. Microorganisms 2020, 8, 733. [Google Scholar] [CrossRef]
- Costa, M.P.; Rosario, A.I.L.; Silva, V.L.; Vieira, C.P.; Conte-Junior, C.A. Rheological, physical and sensory evaluation of low-fat cupuassu goat milk yogurts supplemented with fat replacer. Food Sci. Anim. Resour. 2022, 42, 210. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Li, S.; Li, Q.; Fan, W.; Kiatoukosin, L.; Chen, J. Extracellular polysaccharides of endophytic fungus Alternaria tenuissima F1 from Angelica sinensis: Production conditions, purification, and antioxidant properties. Int. J. Biol. Macromol. 2019, 133, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.Y.M.; Reehana, N.; Jayaraj, K.A.; Ahamed, A.A.P.; Dhanasekaran, D.; Thajuddin, N.; Alharbi, N.S.; Muralitharan, G. Statistical optimization of exopolysaccharide production by Lactobacillus plantarum NTMI05 and NTMI20. Int. J. Biol. Macromol. 2016, 93, 731–745. [Google Scholar] [CrossRef]
- Alshawwa, S.Z.; Alshallash, K.S.; Ghareeb, A.; Elazzazy, A.M.; Sharaf, M.; Alharthi, A.; Abdelgawad, F.E.; El-Hossary, D.; Jaremko, M.; Emwas, A.-H. Assessment of Pharmacological Potential of Novel Exopolysaccharide Isolated from Marine Kocuria sp. Strain AG5: Broad-Spectrum Biological Investigations. Life 2022, 12, 1387. [Google Scholar] [CrossRef]
- Jiang, G.; Li, R.; He, J.; Yang, L.; Chen, J.; Xu, Z.; Zheng, B.; Yang, Y.; Xia, Z.; Tian, Y. Extraction, Structural Analysis, and Biofunctional Properties of Exopolysaccharide from Lactiplantibacillus pentosus B8 Isolated from Sichuan Pickle. Foods 2022, 11, 2327. [Google Scholar] [CrossRef]
- Fan, Y.; Li, X.; Tian, R.; Tang, R.; Zhang, J. Characterization and biological activity of a novel exopolysaccharide produced by Pediococcus pentosaceus SSC–12 from silage. Microorganisms 2021, 10, 18. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Z.; Qiu, L.; Zhang, F.; Xu, X.; Wei, H.; Tao, X. Characterization and bioactivities of the exopolysaccharide from a probiotic strain of Lactobacillus plantarum WLPL04. J. Dairy Sci. 2017, 100, 6895–6905. [Google Scholar] [CrossRef] [PubMed]
- Ismail, B.; Nampoothiri, K.M. Production, purification and structural characterization of an exopolysaccharide produced by a probiotic Lactobacillus plantarum MTCC 9510. Arch. Microbiol. 2010, 192, 1049–1057. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, X.; Tian, Z.; Yang, Y.; Yang, Z. Characterization of an exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibet Kefir. Carbohydr. Polym. 2015, 125, 16–25. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, Z.; Tao, X.; Wei, H. Characterization and sulfated modification of an exopolysaccharide from Lactobacillus plantarum ZDY2013 and its biological activities. Carbohydr. Polym. 2016, 153, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Pachekrepapol, U.; Lucey, J.; Gong, Y.; Naran, R.; Azadi, P. Characterization of the chemical structures and physical properties of exopolysaccharides produced by various Streptococcus thermophilus strains. J. Dairy Sci. 2017, 100, 3424–3435. [Google Scholar] [CrossRef]
- Fuso, A.; Bancalari, E.; Castellone, V.; Caligiani, A.; Gatti, M.; Bottari, B. Feeding Lactic Acid Bacteria with Different Sugars: Effect on Exopolysaccharides (EPS) Production and Their Molecular Characteristics. Foods 2023, 12, 215. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Cui, Y.; Yue, F.; Liu, L.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Exopolysaccharides produced by lactic acid bacteria and Bifidobacteria: Structures, physiochemical functions and applications in the food industry. Food Hydrocoll. 2019, 94, 475–499. [Google Scholar] [CrossRef]
- Wu, L.; Sun, H.; Hao, Y.; Zheng, X.; Song, Q.; Dai, S.; Zhu, Z. Chemical structure and inhibition on α-glucosidase of the polysaccharides from Cordyceps militaris with different developmental stages. Int. J. Biol. Macromol. 2020, 148, 722–736. [Google Scholar] [CrossRef]
- Wang, T.; Hu, Q.; Zhou, M.; Xia, Y.; Nieh, M.-P.; Luo, Y. Development of “all natural” layer-by-layer redispersible solid lipid nanoparticles by nano spray drying technology. Eur. J. Pharm. Biopharm. 2016, 107, 273–285. [Google Scholar] [CrossRef]
- Chen, Y.; Xie, M.-Y.; Nie, S.-P.; Li, C.; Wang, Y.-X. Purification, composition analysis and antioxidant activity of a polysaccharide from the fruiting bodies of Ganoderma atrum. Food Chem. 2008, 107, 231–241. [Google Scholar] [CrossRef]
- Ji, X.; Hou, C.; Yan, Y.; Shi, M.; Liu, Y. Comparison of structural characterization and antioxidant activity of polysaccharides from jujube (Ziziphus jujuba Mill.) fruit. Int. J. Biol. Macromol. 2020, 149, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Wang, B. Chemical characterization and ameliorating effect of polysaccharide from Chinese jujube on intestine oxidative injury by ischemia and reperfusion. Int. J. Biol. Macromol. 2011, 48, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Sardari, R.R.; Kulcinskaja, E.; Ron, E.Y.; Björnsdóttir, S.; Friðjónsson, Ó.H.; Hreggviðsson, G.Ó.; Karlsson, E.N. Evaluation of the production of exopolysaccharides by two strains of the thermophilic bacterium Rhodothermus marinus. Carbohydr. Polym. 2017, 156, 1–8. [Google Scholar] [CrossRef]
- Ramamoorthy, S.; Gnanakan, A.; Lakshmana, S.S.; Meivelu, M.; Jeganathan, A. Structural characterization and anticancer activity of extracellular polysaccharides from ascidian symbiotic bacterium Bacillus thuringiensis. Carbohydr. Polym. 2018, 190, 113–120. [Google Scholar] [CrossRef]
- Abarquero, D.; Renes, E.; Fresno, J.M.; Tornadijo, M.E. Study of exopolysaccharides from lactic acid bacteria and their industrial applications: A review. Int. J. Food Sci. Technol. 2022, 57, 16–26. [Google Scholar] [CrossRef]
- Korcz, E.; Varga, L. Exopolysaccharides from lactic acid bacteria: Techno-functional application in the food industry. Trends Food Sci. Technol. 2021, 110, 375–384. [Google Scholar] [CrossRef]
- Han, X.; Yang, Z.; Jing, X.; Yu, P.; Zhang, Y.; Yi, H.; Zhang, L. Improvement of the texture of yogurt by use of exopolysaccharide producing lactic acid bacteria. BioMed Res. Int. 2016, 2016, 7945675. [Google Scholar] [CrossRef]
- Li, C.; Li, W.; Chen, X.; Feng, M.; Rui, X.; Jiang, M.; Dong, M. Microbiological, physicochemical and rheological properties of fermented soymilk produced with exopolysaccharide (EPS) producing lactic acid bacteria strains. LWT-Food Sci. Technol. 2014, 57, 477–485. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EPS | Mw (Da) | Mn (Da) | Mw/Mn | Molar Mass Distributions |
|---|---|---|---|---|
| EPS1a | 2.41 × 104 | 8.16 × 103 | 2.96 | 3000.0~5000.0 g/mol accounting for 25.9%, 5000.0~40,000.0 g/mol accounting for 55.9%, 40,000.0~88,000.0 g/mol accounting for 13.5%, and 88,000.0~209,195.0 g/mol accounting for 4.7%. |
| EPS2b | 1.62 × 104 | 9.36 × 103 | 1.735 | 3000.0~5000.0 g/mol accounting for 16.9%, 5000.0~11,000.0 g/mol accounting for 29.4%, 11,000.0~48,000.0 g/mol accounting for 49.9%, and 48,000.0~78,852.0 g/mol accounting for 3.7%. |
| EPS3c | 6.42 × 103 | 3.52 × 103 | 1.822 | 1600.0~4300.0 g/mol accounting for 49.8%, 4300.0~7200.0 g/mol accounting for 17.0%, 7200.0~20,000.0 g/mol accounting for 26.7%, and 20,000.0~37,352.0 g/mol accounting for 6.5%. |
| EPS4d | 6.45 × 103 | 4.22 × 103 | 1.531 | 2200.0~3200.0 g/mol accounting for 29.1%, 3200.0~6200.0 g/mol accounting for 34.6%, 6200.0~8200.0 g/mol accounting for 14.3%, 8200.0~20,000.0 g/mol accounting for 18.5%, and 20,000.0~45,402.0 g/mol accounting for 3.5%. |
| EPS5e | 1.26 × 104 | 4.87 × 103 | 2.582 | 2000.0~3500.0 g/mol accounting for 39.2%, 3500.0~16,000.0 g/mol accounting for 32.9%, 16,000.0~28,000.0 g/mol accounting for 21.8%, 28,000.0~67,000.0 g/mol accounting for 3.1%, and 67,000.0~164,802.0 g/mol accounting for 3.0%. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Shu, G.; Wang, J.; Wang, Z.; Liu, Y.; Li, Y.; Chen, L. Purification and Identification of EPS Produced by Five Lactic Acid Bacteria and Evaluation of Their Effects on the Texture of Fermented Goat Milk. Fermentation 2023, 9, 527. https://doi.org/10.3390/fermentation9060527
Liu G, Shu G, Wang J, Wang Z, Liu Y, Li Y, Chen L. Purification and Identification of EPS Produced by Five Lactic Acid Bacteria and Evaluation of Their Effects on the Texture of Fermented Goat Milk. Fermentation. 2023; 9(6):527. https://doi.org/10.3390/fermentation9060527
Chicago/Turabian StyleLiu, Gengdian, Guowei Shu, Jiansheng Wang, Zhanmin Wang, Yu Liu, Yilin Li, and Li Chen. 2023. "Purification and Identification of EPS Produced by Five Lactic Acid Bacteria and Evaluation of Their Effects on the Texture of Fermented Goat Milk" Fermentation 9, no. 6: 527. https://doi.org/10.3390/fermentation9060527
APA StyleLiu, G., Shu, G., Wang, J., Wang, Z., Liu, Y., Li, Y., & Chen, L. (2023). Purification and Identification of EPS Produced by Five Lactic Acid Bacteria and Evaluation of Their Effects on the Texture of Fermented Goat Milk. Fermentation, 9(6), 527. https://doi.org/10.3390/fermentation9060527