
The Production of Bioaroma by Auriporia aurulenta Using Agroindustrial Waste as a Substrate in Submerged Cultures

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical
2.2. Agroindustrial Residue
2.3. Strains and Growing Conditions of Crops
2.4. Submerged Fermentation
Kinetics and Optimization of the Fermentation Process for the Production of Aroma Compounds
2.5. Analysis of Volatile Compounds
2.5.1. Separation of Volatile Compounds
2.5.2. Identification of Volatile Compounds
2.6. Statistical Analysis
3. Results
3.1. Volatile Profile of Fermented Products
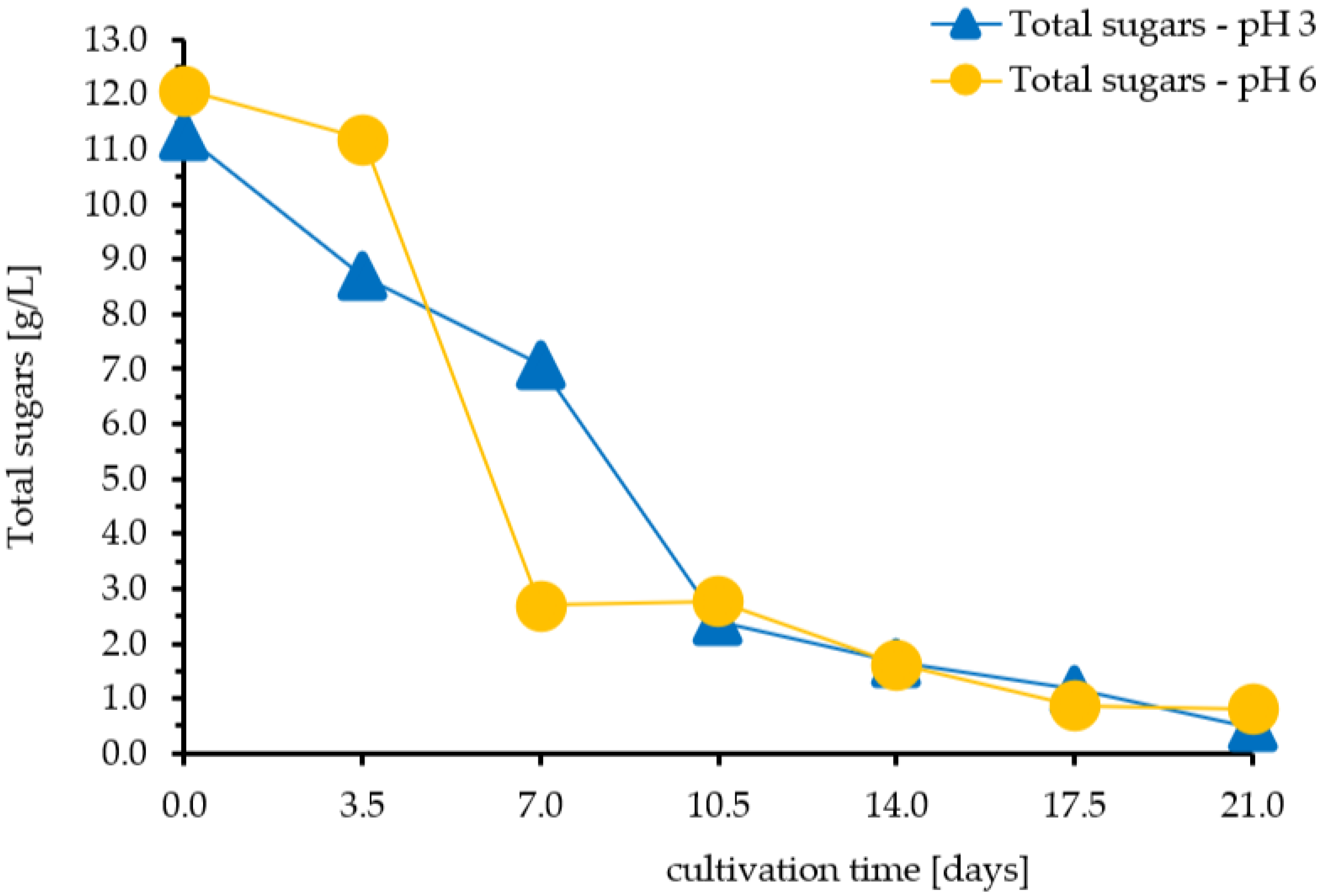
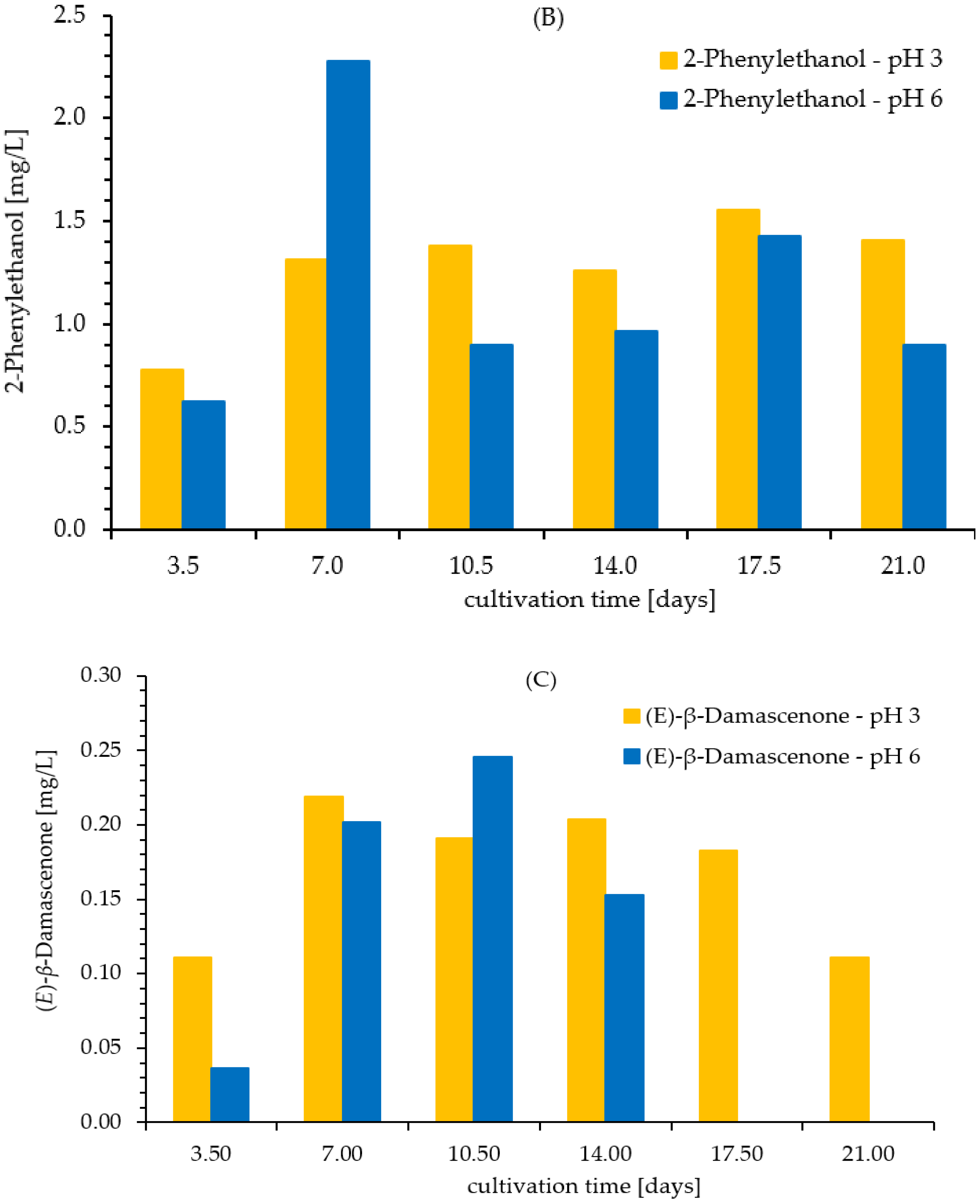
3.2. Kinetics of Formation of Aroma Compounds
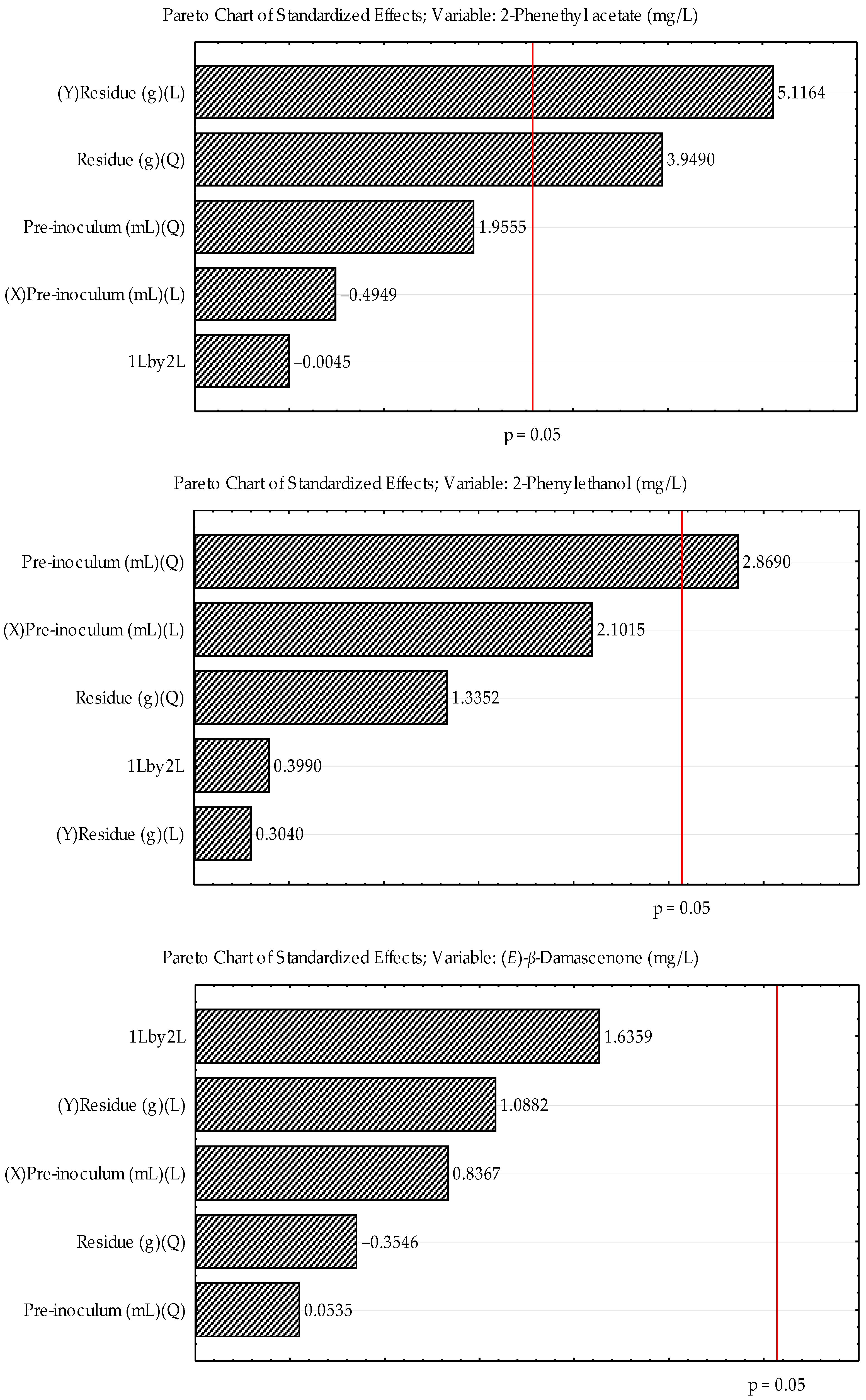
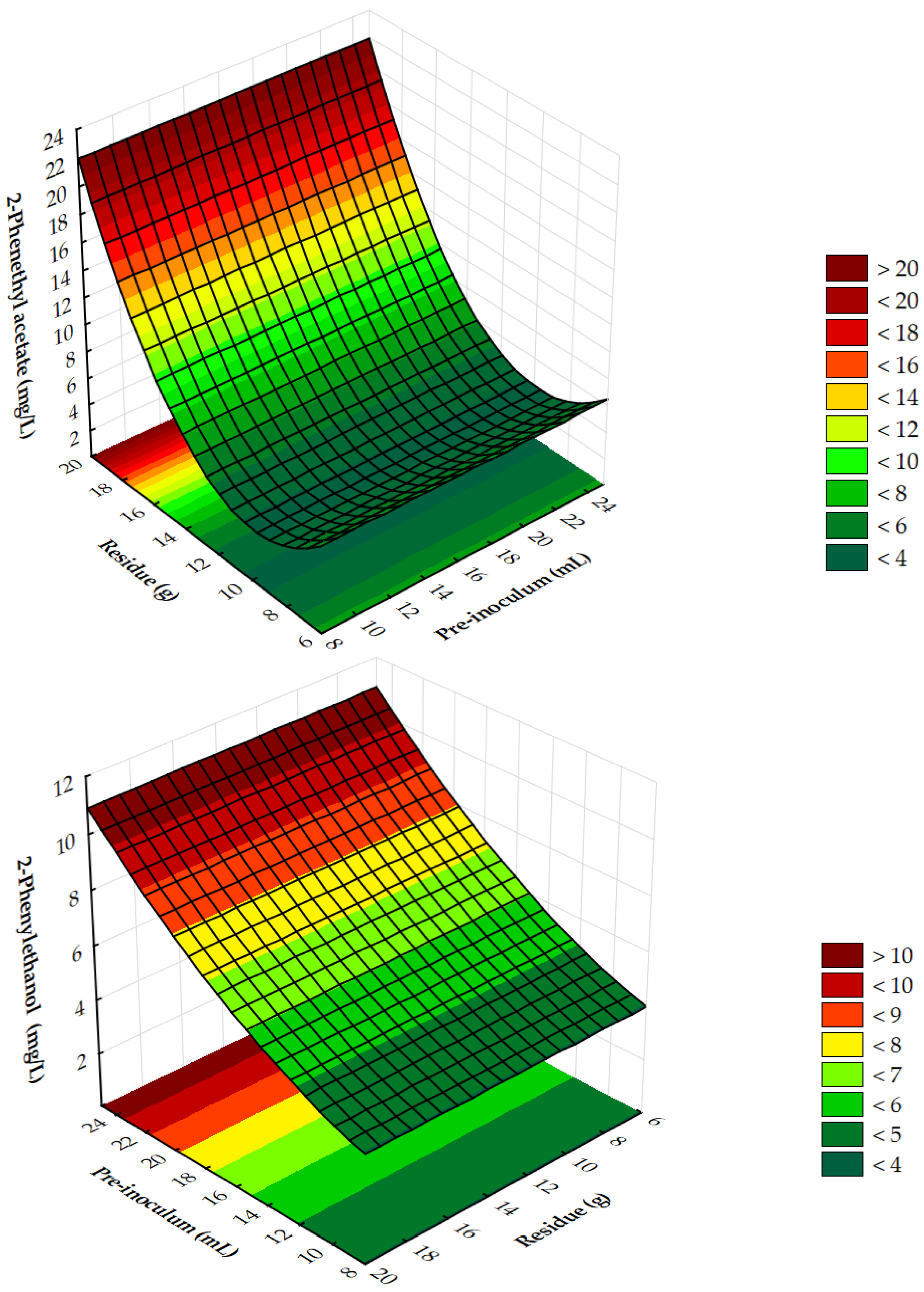
3.3. Fermentation—Process Optimization
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bouws, H.; Wattenberg, A.; Zorn, H. Fungal secretomes—Nature’s toolbox for white biotechnology. Appl. Microbiol. Biotechnol. 2008, 80, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Alchihab, M.; Aldric, J.-M.; Aguedo, M.; Destain, J.; Wathelet, J.-P.; Thonart, P. The use of Macronet resins to recover γ-decalactone produced by Rhodotorula aurantiaca from the culture broth. J. Ind. Microbiol. Biotechnol. 2010, 37, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Fraatz, M.A.; Riemer, S.J.L.; Stöber, R.; Kaspera, R.; Nimtz, M.; Berger, R.G.; Zorn, H. A novel oxygenase from Pleurotus sapidus transforms valencene to nootkatone. J. Mol. Catal. B Enzym. 2009, 61, 202–207. [Google Scholar] [CrossRef]
- Krings, U.; Hinz, M.; Berger, R.G. Degradation of [2H]phenylalanine by the basidiomycete Ischnoderma benzoinum. J. Biotechnol. 1996, 51, 123–129. [Google Scholar] [CrossRef]
- Lapadatescu, C.; Giniès, C.; Le Quéré, J.-L.; Bonnarme, P. Novel scheme for biosynthesis of aryl metabolites from L-phenylalanine in the fungus Bjerkandera adusta. Appl. Environ. Microbiol. 2000, 66, 1517–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krügener, S.; Krings, U.; Zorn, H.; Berger, R.G. A dioxygenase of Pleurotus sapidus transforms (+)-valencene regio-specifically to (+)-nootkatone via a stereo-specific allylic hydroperoxidation. Bioresour. Technol. 2010, 101, 457–462. [Google Scholar] [CrossRef]
- Stentelaire, C.; Lesage-Meessen, L.; Oddou, J.; Bernard, O.; Bastin, G.; Ceccaldi, B.C.; Asther, M. Design of a fungal bioprocess for vanillin production from vanillic acid at scalable level by Pycnoporus cinnabarinus. J. Biosci. Bioeng. 2000, 89, 223–230. [Google Scholar] [CrossRef]
- Patil, P.D.; Yadav, G.D. Comparative studies of white-rot fungal strains (Trametes hirsuta mtcc-1171 and Phanerochaete chrysosporium ncim-1106) for effective degradation and bioconversion of ferulic acid. ACS Omega 2018, 3, 14858–14868. [Google Scholar] [CrossRef] [Green Version]
- Galadima, A.I.; Salleh, M.M.; Hussin, H.; Shiong, C.C.; Yahaya, A.; Mohamad, S.E.; Aziz, S.A.; Yusof, N.N.M.; Al-Junid, A.F.M. Improvement of biovanillin production with two-stage pH control strategy from lemongrass leaves hydrolysates using Phanerochaete chrysosporium ATCC 24725 in batch culture. Biomass Conv. Bioref. 2022, 12, 2727–2736. [Google Scholar] [CrossRef]
- Lesage-Meessen, L.; Lomascolo, A.; Bonnin, E.; Thibault, J.-F.; Buleon, A.; Roller, M.; Asther, M.; Record, E.; Ceccaldi, B.C.; Asther, M. A biotechnological process involving filamentous fungi to produce natural crystalline vanillin from maize bran. Appl. Biochem. Biotechnol. 2002, 102, 141–153. [Google Scholar] [CrossRef]
- Wickramasinghe, P.C.K.; Munafo, J.P. Fermentation dynamics and benzylic derivative production in Ischnoderma resinosum isolates. ACS Omega 2020, 5, 22268–22277. [Google Scholar] [CrossRef] [PubMed]
- Feron, G.; Mauvais, G.; Lherminier, J.; Michel, J.; Wang, X.-D.; Viel, C.; Cachon, R. Metabolism of fatty acid in yeast: Addition of reducing agents to the reaction medium influences β-oxidation activities, γ-decalactone production, and cell ultrastructure in Sporidiobolus ruinenii cultivated on ricinoleic acid methyl ester. Can. J. Microbiol. 2007, 53, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Baqueiro-Peña, I.; Guerrero-Beltrán, J.Á. Vanilla (Vanilla planifolia Andr.), its residues and other industrial by-products for recovering high value flavor molecules: A review. J. Appl. Res. Med. Aromat. Plants 2017, 6, 1–9. [Google Scholar] [CrossRef]
- Priefert, H.; Rabenhorst, J.; Steinbüchel, A. Biotechnological production of vanillin. Appl. Microbiol. Biotechnol. 2001, 56, 296–314. [Google Scholar] [CrossRef] [PubMed]
- Bosse, A.K.; Fraatz, M.A.; Zorn, H. Formation of complex natural flavours by biotransformation of apple pomace with basidiomycetes. Food Chem. 2013, 141, 2952–2959. [Google Scholar] [CrossRef]
- Alvarez, M.V.; Cabred, S.; Ramirez, C.L.; Fanovich, M.A. Valorization of an agroindustrial soybean residue by supercritical fluid extraction of phytochemical compounds. J. Supercrit. Fluids 2019, 143, 90–96. [Google Scholar] [CrossRef]
- Diaz, A.B.; Blandino, A.; Caro, I. Value added products from fermentation of sugars derived from agro-food residues. Trends Food Sci. Technol. 2018, 71, 52–64. [Google Scholar] [CrossRef]
- Freitas, L.C.; Barbosa, J.R.; da Costa, A.L.C.; Bezerra, F.W.F.; Pinto, R.H.H.; Carvalho Junior, R.N.d. From waste to sustainable industry: How can agro-industrial wastes help in the development of new products? Resour. Conserv. Recycl. 2021, 169, 105466. [Google Scholar] [CrossRef]
- Monteiro, S.A.; Barbosa, M.M.; Maia da Silva, F.F.; Bezerra, R.F.; da Silva Maia, K. Preparation, phytochemical and bromatological evaluation of flour obtained from the acerola (Malpighia punicifolia) agroindustrial residue with potential use as fiber source. LWT-Food Sci. Technol. 2020, 134, 110142. [Google Scholar] [CrossRef]
- Karode, B.; Patil, U.; Jobanputra, A. Biotransformation of low cost lignocellulosic substrates into vanillin by white rot fungus, Phanerochaete chrysosporium NCIM 1197. Indian J. Biotechnol. 2013, 12, 281–283. [Google Scholar]
- Thibault, J.; Micard, V.; Renard, C.; Asther, M.; Delattre, M.; Lesage-Meessen, L.; Faulds, C.; Kroon, P.; Williamson, G.; Duarte, J.; et al. Fungal bioconversion of agricultural by-products to vanillin. LWT Food Sci. Technol. 1998, 31, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Rejani, C.T.; Radhakrishnan, S. Microbial conversion of vanillin from ferulic acid extracted from raw coir pith. Nat. Prod. Res. 2020, 36, 901–908. [Google Scholar] [CrossRef]
- Lun, O.K.; Wai, T.B.; Ling, L.S. Pineapple cannery waste as a potential substrate for microbial biotranformation to produce vanillic acid and vanillin. Int. Food Res. J. 2014, 21, 953–958. [Google Scholar]
- Barbosa, E.D.S.; Perrone, D.; Amaral Vendramini, A.L.D.; Ferreira Leite, S.G. Vanillin production by Phanerochaete chrysosporium grown on green coconut agro-industrial husk in solid state fermentation. BioResources 2008, 3, 1042–1050. [Google Scholar]
- Chattopadhyay, P.; Banerjee, G.; Sen, S.K. Cleaner production of vanillin through biotransformation of ferulic acid esters from agroresidue by Streptomyces sannanensis. J. Clean. Prod. 2018, 182, 272–279. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, P.; Singh, J.; Singh, S.; Nain, L. Prospecting the potential of agroresidues as substrate for microbial flavor production. Front. Sustain. Food Syst. 2020, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.L. Use of dinitrosalicylic reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Grosse, M.; Strauss, E.; Krings, U.; Berger, R.G. Response of the sesquiterpene synthesis in submerged cultures of the Basidiomycete Tyromyces floriformis to the medium composition. Mycologia 2019, 111, 885–894. [Google Scholar] [CrossRef]
- Medeiros, A.B.P.; Pandey, A.; Freitas, R.J.S.; Christen, P.; Soccol, C.R. Optimization of the production of aroma compounds by Kluyveromyces marxianus in solid-state fermentation using factorial design and response surface methodology. Biochem. Eng. J. 2000, 6, 33–39. [Google Scholar] [CrossRef]
- Cho, I.H.; Namgung, H.J.; Choi, H.K.; Kim, Y.S. Volatiles and key odorants in the pileus and stipe of pine-mushroom (Tricholoma matsutake Sing.). Food Chem. 2008, 106, 71–76. [Google Scholar] [CrossRef]
- Van den Ban, E.C.D.; Willemen, H.M.; Wassink, H.; Laane, C.; Haaker, H. Bioreduction of carboxylic acids by Pyrococcus furiosus in batch cultures. Enzyme Microb. Technol. 1999, 25, 251–257. [Google Scholar] [CrossRef]
- Kawabe, T.; Morita, H. Production of benzaldehyde and benzyl alcohol by the mushroom Polyporus tuberaster K2606. J. Agric. Food Chem. 1994, 42, 2556–2560. [Google Scholar] [CrossRef]
- Madrera, R.R.; Bedriñana, R.P.; Valles, B.S. Production and characterization of aroma compounds from apple pomace by solid-state fermentation with selected yeasts. LWT Food Sci. Technol. 2015, 64, 1342–1353. [Google Scholar] [CrossRef]
- Rigling, M.; Yadav, M.; Yagishita, M.; Nedele, A.-K.; Sun, J.; Zhang, Y. Biosynthesis of pleasant aroma by enokitake (Flammulina velutipes) with a potential use in a novel tea drink. LWT-Food Sci. Technol. 2020, 140, 110646. [Google Scholar] [CrossRef]
- Pineau, B.; Barbe, J.-C.; Van Leeuwen, C.; Dubourdieu, D. Which impact for β-damascenone on red wines aroma? J. Agric. Food Chem. 2007, 55, 4103–4108. [Google Scholar] [CrossRef]
- Vilanova, M.; Ugliano, M.; Varela, C.; Siebert, T.; Pretorius, I.; Henschke, P. Assimilable nitrogen utilisation and production of volatile and non-volatile compounds in chemically defined medium by Saccharomyces cerevisiae wine yeasts. Appl. Microbiol. Biotechnol. 2007, 77, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Kim, M.J.; Dhandapani, S.; Tjhang, J.G.; Yin, J.-L.; Wong, L.; Sarojam, R.; Chua, N.-H.; Jang, I.-C. The floral transcriptome of ylang ylang (Cananga odorata var. fruticosa) uncovers biosynthetic pathways for volatile organic compounds and a multifunctional and novel sesquiterpene synthase. J. Exp. Bot. 2015, 66, 3959–3975. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhang, Y.; Zu, Y.; Yang, L. Chemical composition and antioxidant activity of the essential oil of Schisandra chinensis fruits. Nat. Prod. Res. 2012, 26, 842–849. [Google Scholar] [CrossRef]
- Jurado, J.M.; Ballesteros, O.; Alcázar, A.; Pablos, F.; Martín, M.J.; Vílchez, J.L.; Navalón, A. Characterization of aniseed-flavoured spirit drinks by headspace solid-phase microextraction gas chromatography–mass spectrometry and chemometrics. Talanta 2007, 72, 506–511. [Google Scholar] [CrossRef]
- Ferreira, A.C.S.; Monteiro, J.; Oliveira, C.; Guedes de Pinho, P. Study of major aromatic compounds in port wines from carotenoid degradation. Food Chem. 2008, 110, 83–87. [Google Scholar] [CrossRef]
- Kolwek, J.; Behrens, C.; Linke, D.; Krings, U.; Berger, R.G. Cell-free one-pot conversion of (+)-valencene to (+)-nootkatone by a unique dye-decolorizing peroxidase combined with a laccase from Funalia trogii. J. Ind. Microbiol. Biotechnol. 2018, 45, 89–101. [Google Scholar] [CrossRef]
- Püth, N.; Ersoy, F.; Berger, R.G.; Krings, U. Impact of agro-industrial side-streams on sesquiterpene production by submerged cultured Cerrena unicolor. Foods 2023, 12, 668. [Google Scholar] [CrossRef] [PubMed]
- Tilay, A.; Bule, M.; Annapure, U. Production of biovanillin by one-step biotransformation using fungus Pycnoporous cinnabarinus. J. Agric. Food Chem. 2010, 58, 4401–4405. [Google Scholar] [CrossRef]
- Tang, P.L.; Hassan, O. Bioconversion of ferulic acid attained from pineapple peels and pineapple crown leaves into vanillic acid and vanillin by Aspergillus niger I-1472. BMC Chem. 2020, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Galadima, A.I.; Salleh, M.M.; Hussin, H.; Mohd Safri, N.; Mohd Noor, R.; Chong, C.S.; Yahya, A.; Mohamad, S.E.; Abd-Aziz, S.; Yusof, N.N.M.; et al. One-step conversion of lemongrass leaves hydrolysate to biovanillin by Phanerochaete chrysosporium atcc 24725 in batch culture. Waste Biomass Valorization 2020, 11, 4067–4080. [Google Scholar] [CrossRef]
- Mantzouridou, F.T.; Paraskevopoulou, A.; Lalou, S. Yeast flavour production by solid state fermentation of orange peel waste. Biochem. Eng. J. 2015, 101, 1–8. [Google Scholar] [CrossRef]
- Rossi, S.C.; Vandenberghe, L.P.S.; Pereira, B.M.P.; Gago, F.D.; Rizzolo, J.A.; Pandey, A.; Soccol, C.R.; Medeiros, A.B.P. Improving fruity aroma production by fungi in SSF using citric pulp. Food Res. Int. 2009, 42, 484–486. [Google Scholar] [CrossRef]
- Lu, Z.-M.; Tao, W.-Y.; Xu, H.-Y.; Lim, J.; Zhang, X.-M.; Wang, L.-P.; Chen, J.-H.; Xu, Z.-H. Analysis of volatile compounds of Antrodia camphorata in submerged culture using headspace solid-phase microextraction. Food Chem. 2011, 127, 662–668. [Google Scholar] [CrossRef]
- Kawabe, T.; Morita, H. Volatile components in culture fluid of Polyporus tuberaster. J. Agric. Food Chem. 1993, 41, 637–640. [Google Scholar] [CrossRef]
- Gallois, A.; Gross, B.; Langlois, D.; Spinnler, H.-E.; Brunerie, P. Influence of culture conditions on production of flavour compounds by 29 ligninolytic Basidiomycetes. Mycol. Res. 1990, 94, 494–504. [Google Scholar] [CrossRef]
- Lomascolo, A.; Stentelaire, C.; Asther, M.; Lesage-Meessen, L. Basidiomycetes as new biotechnological tools to generate natural aromatic flavours for the food industry. Trends Biotechnol. 1999, 17, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Abraham, B.G.; Berger, R.G. Higher fungi for generating aroma components through novel biotechnologies. J. Agric. Food Chem. 1994, 42, 2344–2348. [Google Scholar] [CrossRef]
- Gross, B.; Gallois, A.; Spinnler, H.-E.; Langlois, D. Volatile compounds produced by the ligninolytic fungus Phlebia radiata Fr. (Basidiomycetes) and influence of the strain specificity on the odorous profile. J. Biotechnol. 1989, 10, 303–308. [Google Scholar] [CrossRef]
- Chen, D.; Yap, Z.Y.; Liu, S.-Q. Evaluation of the performance of Torulaspora delbrueckii, Williopsis saturnus, and Kluyveromyces lactis in lychee wine fermentation. Int. J. Food Microbiol. 2015, 206, 45–50. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variables | −1.41 | −1 | 0 | 1 | 1.41 |
|---|---|---|---|---|---|
| X (mL) | 8.00 | 13.12 | 16.50 | 21.56 | 25.00 |
| Y (g) | 6.25 | 9.04 | 13.12 | 17.22 | 20.00 |
| Compounds | RI+ | RI* | Umbu Residue | Cajá Residue | Persimmon Residue | Plum Residue | Control | Odor Description |
|---|---|---|---|---|---|---|---|---|
| 2-Methyl-1-propanol | 618 | 618 | 0.87 a | 0.68 b | 0.50 c | 0.41 d | 0.91 a | Wine |
| 2-Methyl-1-butanol | 737 | 736 | 0.69 b | 0.28 c | 0.20 d | 0.07 e | 1.07 a | Alcoholic, fruity, wine |
| 3-Methyl-1-butanol | 765 | 768 | 2.66 a | 0.88 d | 1.83 b | 0.80 d | 1.63 c | Fermented, banana |
| 2-Methylpropyl acetate | 772 | 772 | 0.06 a | 0.03 b | Sweet, fruity, banana | |||
| Isoamyl acetate | 876 | 877 | 0.80 a | 0.57 b | 0.23 c | 0.20 c | 0.82 a | Sweet, fruity, banana |
| 2-Methylbutanol acetate | 879 | 879 | 0,13 a | 0.03 b | Fruity, sweet, banana | |||
| Prenyl acetate | 910 | 909 | 0,01 c | 0.02 b | 0.04 a | Banana, fruity, jasmine | ||
| Benzaldehyde | 962 | 962 | 0.03 b | 1.27 a | Fruity, almond | |||
| 1-Octen-3-ol | 980 | 980 | 0.07 b | 0.01 d | 0.01 d | 0.12 a | 0.03 c | Green, fungus, mushroom |
| 3-Octanone | 987 | 984 | 0.19 a | 0.19 a | 0.14 b | 0.14 b | 0.09 c | Moldy, mushroom |
| Butyl butanoate | 995 | 995 | 0.03 | |||||
| 3-Octanol | 996 | 996 | 0.13 b.c | 0.15 a | 0.12 c | 0.14 b | 0.06 d | Woody, mushroom |
| (Z)-3-Hexenyl acetate | 1008 | 1005 | 0.46 a | 0.09 b | Sweet, fruity, banana | |||
| Hexyl acetate | 1015 | 1015 | 0.22 a | 0.03 b | 0.03 b | Fruity, banana | ||
| (E)-2-Hexenyl acetate | 1018 | 1007 | 0.10 | Apple, banana | ||||
| Benzyl alcohol | 1037 | 1038 | 0.61 d | 2.07 a | 0.64 c,d | 1.68 b | 0.76 c | Fruity, floral, candy |
| (Z)-3-Octenol | 1050 | 1047 | 0.02 | Fresh, herbal, melon | ||||
| (E)-2-Octenol | 1065 | 1069 | 0.02 c | 0.16 a | 0.15 b | 0.01 d | Vegetable, citrus, green | |
| 1-Octanol | 1071 | 1070 | 0.06 c | 0.07 c | 0.25 a | 0.19 b | 0.07 c | Citric, mushroom |
| (Z)-5-Octenol | 1071 | 1051 | 0.10 | Green, mushroom | ||||
| Linalool | 1100 | 1099 | 0.14 b | 0.01 c | 0.31 a | Pink, floral, orange | ||
| Heptyl acetate | 1112 | 1114 | 0.04 | Woody, apricot, pear | ||||
| 1-Octenyl-3-acetate | 1113 | 1113 | 0.21 | Fruity, green, herbal | ||||
| 2-Phenylethanol | 1118 | 1112 | 2.25 a | 0.24 c | 0.01 d | 0.77 b | 0.27 c | Floral, roses |
| Benzyl acetate | 1169 | 1162 | 0.68 c | 1.32 b | 0.15 d | 0.16 d | 1.41 a | Floral, fruity, jasmine |
| Methyl salicylate | 1199 | 1200 | 0.06 | Slightly, phenolic, mint | ||||
| Octyl acetate | 1235 | 1220 | 0.01 d | 0.24 a | 0.03 c | 0.09 b | Floral | |
| 3-Phenylpropanol | 1254 | 1253 | 0.37 b | 0.60 a | 0.04 d | 0.13 c | Spicy, sweet | |
| Heptyl butanoate | 1273 | 1275 | 0.07 | Fruity, floral, green, tea | ||||
| 2-Phenethyl acetate | 1277 | 1271 | 5.07 a | 0.84 b.c | 0.77 b | 1.24 b | 0.54 c | Fruity, honey, rose, floral |
| γ-Octalactone | 1278 | 1277 | 0.37 a | 0.09 b | 0.01 c | 0.01 c | 0.10 b | Sweet, coconut |
| (E)-Cinnamaldehyde | 1280 | 1283 | 0.06 | Sweet, cinnamon | ||||
| p-Cymen-7-ol | 1295 | 1295 | 0.18 | Cumin, spicy, herbs | ||||
| Myrtenyl acetate | 1299 | 1305 | 0.05 a | 0.01 b | Fruity, sweet, herbal | |||
| Cinnamyl alcohol | 1307 | 1312 | 0.39 | Sweet, green, hyacinth | ||||
| 1,4-p-Menthadien-7-ol | 1320 | 1315 | 0.16 | |||||
| Benzyl butanoate | 1343 | 1345 | 0.06 | Apricot, fruity, jasmine | ||||
| 2-Phenylethyl propanoate | 1350 | 1350 | 0.02 | Honey, fruity, floral | ||||
| 3-Phenylpropyl acetate | 1371 | 1373 | 0.68 b | 0.21 c | 0.04 e | 0.11 d | 0.77 a | Spicy, cinnamon |
| Methyl cinnamate | 1383 | 1380 | 0.02 a | 0.03 a | Sweet, balsamic | |||
| (E)-2-Hexenyl hexanoate | 1384 | 1375 | 0.03 | Herbal, green | ||||
| (E)-β-Damascenone | 1386 | 1382 | 0.18 | Fruity, honey, rose, apple | ||||
| Cuminyl acetate | 1422 | 1432 | 0.40 | Fresh, fruity, sweet, herbs | ||||
| Cinnamyl acetate | 1451 | 1453 | 0.33 b | 0.41 a | Cinnamon, floral, sweet | |||
| Ethyl (E)-cinnamate | 1456 | 1463 | 0.04 | Floral, honey |
| Experimental Values | Predicted Values | ||||||
|---|---|---|---|---|---|---|---|
| Assay | Pre-Inoculum (mL) | Residue (g) | 2-Phenethyl Acetate (mg/L) | 2-Phenylethanol (mg/L) | (E)-β-Damascenone (mg/L) | 2-Phenethyl Acetate (mg/L) | 2-Phenylethanol (mg/L) |
| 1 | 11.44 | 9.04 | 7.32 | 5.41 | 0.19 | 4.00 | 4.80 |
| 2 | 21.56 | 9.04 | 3.57 | 6.71 | 0.18 | 4.00 | 8.93 |
| 3 | 11.44 | 17.22 | 13.05 | 2.86 | 0.23 | 13.43 | 4.80 |
| 4 | 21.56 | 17.22 | 9.28 | 6.25 | 0.47 | 13.43 | 8.93 |
| 5 | 8.00 | 13.12 | 8.25 | 8.22 | 0.24 | 5.78 | 3.97 |
| 6 | 25.00 | 13.12 | 10.10 | 15.56 | 0.23 | 5.78 | 10.91 |
| 7 | 16.5 | 6.24 | 4.23 | 6.65 | 0.23 | 6.14 | 6.55 |
| 8 | 16.5 | 20.00 | 24.47 | 9.90 | 0.19 | 21.96 | 6.55 |
| 9 | 16.5 | 13.12 | 4.10 | 4.15 | 0.22 | 5.78 | 6.55 |
| 10 | 16.5 | 13.12 | 4.11 | 4.67 | 0.24 | 5.78 | 6.55 |
| 11 | 16.5 | 13.12 | 3.41 | 4.68 | 0.26 | 5.78 | 6.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandes, R.D.D.; De Jesus, M.S.; Araujo, H.C.S.; Dos Santos, R.A.R.; Nogueira, J.P.; Leite Neta, M.T.S.; Narain, N. The Production of Bioaroma by Auriporia aurulenta Using Agroindustrial Waste as a Substrate in Submerged Cultures. Fermentation 2023, 9, 593. https://doi.org/10.3390/fermentation9070593
Sandes RDD, De Jesus MS, Araujo HCS, Dos Santos RAR, Nogueira JP, Leite Neta MTS, Narain N. The Production of Bioaroma by Auriporia aurulenta Using Agroindustrial Waste as a Substrate in Submerged Cultures. Fermentation. 2023; 9(7):593. https://doi.org/10.3390/fermentation9070593
Chicago/Turabian StyleSandes, Rafael Donizete Dutra, Mônica Silva De Jesus, Hannah Caroline Santos Araujo, Raquel Anne Ribeiro Dos Santos, Juliete Pedreira Nogueira, Maria Terezinha Santos Leite Neta, and Narendra Narain. 2023. "The Production of Bioaroma by Auriporia aurulenta Using Agroindustrial Waste as a Substrate in Submerged Cultures" Fermentation 9, no. 7: 593. https://doi.org/10.3390/fermentation9070593
APA StyleSandes, R. D. D., De Jesus, M. S., Araujo, H. C. S., Dos Santos, R. A. R., Nogueira, J. P., Leite Neta, M. T. S., & Narain, N. (2023). The Production of Bioaroma by Auriporia aurulenta Using Agroindustrial Waste as a Substrate in Submerged Cultures. Fermentation, 9(7), 593. https://doi.org/10.3390/fermentation9070593