
Concentrated Manipueira as an Alternative Low-Cost Substrate to Rhodotorula glutinis for Biotechnological Production of High Contents of Carotenoids

 , and
, and 
Abstract
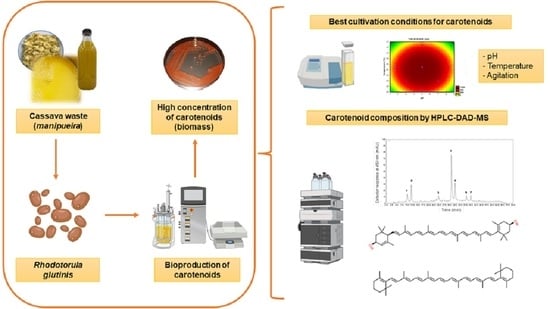
:
1. Introduction
2. Materials and Methods
2.1. Manipueira
2.2. Physico–Chemical Characterization of Concentrated Manipueira
2.3. Starter Culture Preparation
2.4. Carotenoids and Biomass Production
2.5. Carotenoid Composition by HPLC-DAD-APCI-MS/MS
2.6. Statistical Analysis
3. Results and Discussion
3.1. Physico–Chemical Characterization of the Concentrated Manipueira
3.2. Best Cultivation Conditions for Carotenoids and Biomass Production
3.3. Carotenoid Composition Produced by Rhodotorula glutinis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zoz, L.; Carvalho, J.C.; Soccol, V.T.; Casagrande, T.C.; Cardoso, L. Torularhodin and torulene: Bioproduction, properties and prospective applications in food and cosmetics—A review. Braz. Arch. Biol. Technol. 2015, 58, 278–288. [Google Scholar] [CrossRef] [Green Version]
- Squina, F.M.; Mercadante, A.Z. Análise, por CLAE, de carotenóides de cinco linhagens de Rhodotorula. Rev. Bras. Cienc. Farm. 2003, 39, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Valduga, E.; Tatsch, P.O.; Tiggemann, L.; Treichel, H.; Toniazzo, G.; Zeni, J.; Di Luccio, M.; Fúrigo Júnior, A. Carotenoids production: Microorganisms as source of natural dyes. Quim. Nova 2009, 32, 2429–2436. [Google Scholar] [CrossRef]
- Igreja, W.S.; Maia, F.A.; Lopes, A.S.; Chisté, R.C. B’iotechnological Production of Carotenoids Using Low Cost-Substrates Is Influenced by Cultivation Parameters: A Review. Int. J. Mol. Sci. 2021, 22, 8819. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Almanza, A.; Montanez, J.C.; Aguilar-González, M.A.; Martínez-Ávila, C.; Rodríguez-Herrera, R.; Aguilar, C.N. Rhodotorula glutinis as source of pigments and metabolites for food industry. Food Biosci. 2014, 5, 64–72. [Google Scholar] [CrossRef]
- Mata-Gómez, L.C.; Montañez, J.C.; Méndez-Zavala, A.; Aguilar, C.N. Biotechnological production of carotenoids by yeasts: An overview. Microb. Cell Factories 2014, 13, 12. [Google Scholar] [CrossRef] [Green Version]
- Britton, G.; Liaanen-Jensen, S.; Pfander, H. Carotenoids; Birkhäuser: Basel, Switzerland, 2004. [Google Scholar] [CrossRef]
- Mercadante, A.Z. Carotenoids in Foods: Sources and Stability during Processing and Storage; Socaciu, C., Ed.; CRC Press: New York, NY, USA, 2007; pp. 213–235. [Google Scholar]
- Khoo, K.S.; Lee, S.Y.; Ooi, C.W.; Fu, X.; Miao, X.; Ling, T.C.; Show, P.L. Recent advances in biorefinery of astaxanthin from Haematococcus pluvialis. Bioresour. Technol. 2019, 288, 121606. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, X.; Too, H.-P. Microbial astaxanthin biosynthesis: Recent achievements, challenges, and commercialization outlook. Appl. Microbiol. Biotechnol. 2020, 104, 5725–5737. [Google Scholar] [CrossRef]
- Arthur, E.K.; Azeko, S.T. Surface hardening of ferrous materials with cassava (Manihot spp.) waste: A review. Sci. Afr. 2020, 9, e00483. [Google Scholar] [CrossRef]
- Chaiareekitwat, S.; Latif, S.; Mahayothee, B.; Khuwijitjaru, P.; Nagle, M.; Amawan, S.; Müller, J. Protein composition, chlorophyll, carotenoids, and cyanide content of cassava leaves (Manihot esculenta Crantz) as influenced by cultivar, plant age, and leaf position. Food Chem. 2022, 372, 131173. [Google Scholar] [CrossRef] [PubMed]
- Cereda, M.P.; Vasconcelos, S.P. Varieties and Landraces Cultural Practices and Traditional Uses; Volume 2 in Underground Starchy Crops of South American Origin; Chapter 9—Cassava Cyanogenic Glycosides: Importance, Toxicity, and Dosage Methods; Academic Press: Cambridge, MA, USA, 2023; pp. 179–209. [Google Scholar] [CrossRef]
- Brito, B.N.C.; Chisté, R.C.; Lopes, A.S.; Glória, M.B.A.; Pena, R.S. Influence of spontaneous fermentation of manipueira on bioactive amine and carotenoid profiles during tucupi production. Food Res. Int. 2019, 120, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Maróstica, M.R., Jr.; Pastore, G.M. Biotransformation of citronellol in rose-oxide using cassava wastewater as a medium. Food Sci. Technol. 2006, 26, 690–696. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1997; Volume 2, 850 p. [Google Scholar]
- Mandelli, F.; Miranda, V.S.; Rodrigues, E.; Mercadante, A.Z. Identification of carotenoids with high antioxidant capacity produced by extremophile microorganisms. World J. Microbiol. Biotechnol. 2012, 28, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Meléndez-Martínez, A.J.; Britton, G.; Vicario, I.M.; Heredia, F.J. Relationship between the colour and the chemical structure of carotenoid pigments. Food Chem. 2007, 101, 1145–1150. [Google Scholar] [CrossRef]
- Chisté, R.C.; Mercadante, A.Z. Identification and Quantification, by HPLC-DAD-MS/MS, of Carotenoids and Phenolic Compounds from the Amazonian Fruit Caryocar villosum. J. Agric. Food Chem. 2012, 60, 5884–5892. [Google Scholar] [CrossRef] [PubMed]
- De Rosso, V.V.; Mercadante, A.Z. Identification and quantification of carotenoids by HPLC-PDA-MS/MS, from Amazonian fruits. J. Agric. Food Chem. 2007, 55, 5062–5072. [Google Scholar] [CrossRef]
- Santana Neto, J.A.; Muniz, E.N.; Santos, G.R.A.; Macedo, F.A.F.; Rangel, J.H.D.A.; Gomes, L.C. Effect of cassava wastewater on physicochemical characteristics and fatty acids composition of meat from feedlot-finished lambs. Acta Sci.—Anim. Sci. 2017, 39, 377. [Google Scholar] [CrossRef] [Green Version]
- Damasceno, S.; Cereda, M.P.; Pastore, G.M.; Oliveira, J.G. Production of volatile compounds by Geotrichum fragrans using cassava wastewater as substrate. Process Biochem. 2003, 39, 411–414. [Google Scholar] [CrossRef]
- Ribeiro, J.E.S.; Sant’Ana, A.M.S.; Martini, M.; Sorce, C.; Andreucci, A.; Melo, D.J.N.; Silva, F.L.H. Rhodotorula glutinis cultivation on cassava wastewater for carotenoids and fatty acids generation. Biocatal. Agric. Biotechnol. 2019, 22, 101419. [Google Scholar] [CrossRef]
- Buzzini, P.; Martini, A. Production of carotenoids by strains of Rhodotorula glutinis cultured in raw materials of agro-industrial origin. Bioresour. Technol. 2000, 71, 41–44. [Google Scholar] [CrossRef]
- Bhosale, P.; Gadre, R. β-Carotene production in sugarcane molasses by a Rhodotorula glutinis mutant. J. Ind. Microbiol. Biotechnol. 2001, 26, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Sakaki, H.; Nakanishi, T.; Tada, A.; Miki, W.; Komemushi, S. Activation of torularhodin production by Rhodotorula glutinis using weak white light irradiation. J. Biosci. Bioeng. 2001, 92, 294–297. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.L.; Goodwin, T.W. A comparison of synthetic β-zeacarotene and samples obtained from Rhodotorula glutinis and yellow corn. Phytochemistry 1965, 4, 193–195. [Google Scholar] [CrossRef]
- Maldonade, I.R.; Rodriguez-Amaya, D.B.; Scamparini, A.R.P. Carotenoids of yeasts isolated from the Brazilian ecosystem. Food Chem. 2008, 107, 145–150. [Google Scholar] [CrossRef]
- Squina, F.M.; Yamashita, F.; Pereira, J.L.; Mercadante, A.Z. Production of carotenoids by Rhodotorula rubra and R. glutinis in culture medium supplemented with sugar cane juice. Food Biotechnol. 2002, 16, 227–235. [Google Scholar] [CrossRef]
- El-Banna, A.A.; El-Razek, A.M.A.; El-Mahdy, A.R. Some Factors Affecting the Production of Carotenoids by Rhodotorula glutinis var. glutinis. Food Nutr. Sci. 2012, 3, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Simpson, K.L.; Nakayama, T.O.M.; Chichester, C.O. Biosynthesis of yeast carotenoids. J. Bacteriol. 1964, 88, 1688–1694. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.A.; An, G.H. Astaxanthin from Microbial Sources. Crit. Rev. Biotechnol. 1991, 11, 297–326. [Google Scholar] [CrossRef]
- Berera, R.; van Stokkum, I.H.M.; Kennis, J.T.M.; van Grondelle, R.; Dekker, J.P. The light-harvesting function of carotenoids in the cyanobacterial stress-inducible IsiA complex. Chem. Phys. 2010, 373, 65–70. [Google Scholar] [CrossRef]
- Kot, A.M.; Błażejak, S.; Gientka, I.; Kieliszek, M.; Bryś, J. Torulene and torularhodin: “new” fungal carotenoids for industry? Microb. Cell Fact. 2018, 17, 49. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Experiments | Variables | Total Carotenoids (μg/g Dry Biomass) | Dry Biomass (g) | ||
|---|---|---|---|---|---|
| pH | Temperature (°C) | Agitation (rpm) | |||
| 1 | 3.0 (−1) | 30 (−1) | 100 (−1) | 1028.7 | 10.3 |
| 2 | 7.0 (+1) | 30 (−1) | 100 (−1) | 674.6 | 6.5 |
| 3 | 3.0 (−1) | 40 (+1) | 100 (−1) | 882.2 | 8.9 |
| 4 | 7.0 (+1) | 40 (+1) | 100 (−1) | 673.4 | 7.5 |
| 5 | 3.0 (−1) | 30 (−1) | 200 (+1) | 788.9 | 9.8 |
| 6 | 7.0 (+1) | 30 (−1) | 200 (+1) | 759.2 | 4.7 |
| 7 | 3.0 (−1) | 40 (+1) | 200 (+1) | 641.1 | 6.7 |
| 8 | 7.0 (+1) | 40 (+1) | 200 (+1) | 666.4 | 7.5 |
| 9 | 1.64 (−α) | 35 (0) | 150 (0) | 134.8 | 1.9 |
| 10 | 8.36 (+α) | 35 (0) | 150 (0) | 516.3 | 7.4 |
| 11 | 5.0 (0) | 27 (−α) | 150 (0) | 538.6 | 4.0 |
| 12 | 5.0 (0) | 43 (+α) | 150(0) | 106.0 | 5.3 |
| 13 | 5.0 (0) | 35 (0) | 66 (−α) | 523.9 | 4.4 |
| 14 | 5.0 (0) | 35 (0) | 234 (+α) | 529.0 | 5.7 |
| 15 | 5.0 (0) | 35 (0) | 150 (0) | 1432.2 | 11.1 |
| 16 | 5.0 (0) | 35 (0) | 150 (0) | 1381.3 | 10.4 |
| 17 | 5.0 (0) | 35 (0) | 150 (0) | 1415.1 | 10.9 |
| Parameter | Content * |
|---|---|
| Moisture (g/100 mL) | 89.57 ± 0.17 |
| Total ashes (g/100 mL) | 0.61 ± 0.01 |
| Total lipids (g/100 mL) | 1.28 ± 0.11 |
| Total proteins (g/100 mL) | 0.70 ± 0.03 |
| Total carbohydrates (g/100 mL) | 7.85 ± 0.01 |
| pH | 6.24 ± 0.02 |
| Total acidity (meq NaOH/100 mL) | 2.68 ± 0.11 |
| Soluble solids (Brix) | 11.93 ± 0.12 |
| Reducing sugars (g/100 mL) | 4.19 ± 0.06 |
| Total sugars (g/100 mL) | 7.21 ± 0.05 |
| Peak a | Carotenoid | Contents (μg/g Dry Biomass) | tR (min) b | λmax (nm) c | %III/II | %AB/AII | [M+H]+ (m/z) | MS/MS Fragment Ions (m/z) |
|---|---|---|---|---|---|---|---|---|
| 1 | 5,8-Epoxy-lutein | 220.57 ± 4.90 | 10.2 | 398, 420, 446 | 80 | 0 | n.d. | 567.5 [M+H-18]+ → 549.4 [M+H-18-18]+; 531.5 [M+H-18-18-18]+; 511.4 [M+H-56-18]+; 475.3 [M+H-92-18]+; 221.0 |
| 2 | (all-E)-Lutein | 242.37 ± 4.11 | 12.3 | 420, 444, 472 | 60 | 0 | 569.5 | 551.4 [M+H-18]+; 533.4 [M+H-18-18]+; 495.4 [M+H-56-18]+; 459.2 [M+H-92-18]+ |
| 3 | (13Z)-β-Carotene | 108.89 ± 20.87 | 25.1 | 338, 422, 446, 468 | 25 | 39 | 537.4 | 444.3 [M-92]+ |
| 4 | (all-E)-β-Carotene | 239.32 ± 16.85 | 31.6 | 426, 450, 478 | 12 | 0 | 537.4 | 444.3 [M-92]+ |
| 5 | (9Z)-β-Carotene | 150.74 ± 7.36 | 33.7 | 338, 420, 446, 472 | 25 | 8 | 537.4 | 444.3 [M-92]+ |
| 6 | β-Zeacarotene 1 | 64.89 ± 5.38 | 39.2 | 402, 426, 452 | 78 | 0 | 539.5 | 446.2 [M-92]+; 401.2 [M-137]+; 309.1 [M-137-92]+ |
| 7 | β-Zeacarotene 2 | 69.24 ± 3.60 | 41.3 | 404, 426, 450 | 78 | n.c | 539.5 | 446.2 [M-92]+; 401.2 [M-137]+; 309.1 [M-137-92]+ |
| Total sum | 1096.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maia, F.d.A.; Igreja, W.S.; Xavier, A.A.O.; Mercadante, A.Z.; Lopes, A.S.; Chisté, R.C. Concentrated Manipueira as an Alternative Low-Cost Substrate to Rhodotorula glutinis for Biotechnological Production of High Contents of Carotenoids. Fermentation 2023, 9, 617. https://doi.org/10.3390/fermentation9070617
Maia FdA, Igreja WS, Xavier AAO, Mercadante AZ, Lopes AS, Chisté RC. Concentrated Manipueira as an Alternative Low-Cost Substrate to Rhodotorula glutinis for Biotechnological Production of High Contents of Carotenoids. Fermentation. 2023; 9(7):617. https://doi.org/10.3390/fermentation9070617
Chicago/Turabian StyleMaia, Felipe de Andrade, Willen Silva Igreja, Ana Augusta Odorissi Xavier, Adriana Zerlotti Mercadante, Alessandra Santos Lopes, and Renan Campos Chisté. 2023. "Concentrated Manipueira as an Alternative Low-Cost Substrate to Rhodotorula glutinis for Biotechnological Production of High Contents of Carotenoids" Fermentation 9, no. 7: 617. https://doi.org/10.3390/fermentation9070617
APA StyleMaia, F. d. A., Igreja, W. S., Xavier, A. A. O., Mercadante, A. Z., Lopes, A. S., & Chisté, R. C. (2023). Concentrated Manipueira as an Alternative Low-Cost Substrate to Rhodotorula glutinis for Biotechnological Production of High Contents of Carotenoids. Fermentation, 9(7), 617. https://doi.org/10.3390/fermentation9070617