
Comparative Analysis of TPR Gene Family in Cucurbitaceae and Expression Profiling under Abiotic Stress in Cucumis melo L.

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatment
2.2. Identification of TPRs and Their Biochemical Characterization
2.3. Phylogenetic Relationship, Exon/Intron Structure, and Protein Motif
2.4. Chromosomal Location and Collinearity Analyses
2.5. Promoter Cis-Regulatory Element and Gene Ontology Analysis
2.6. Expression Analysis of CmTPR Genes
3. Results
3.1. Identification of TPR Genes in Cucurbitaceae
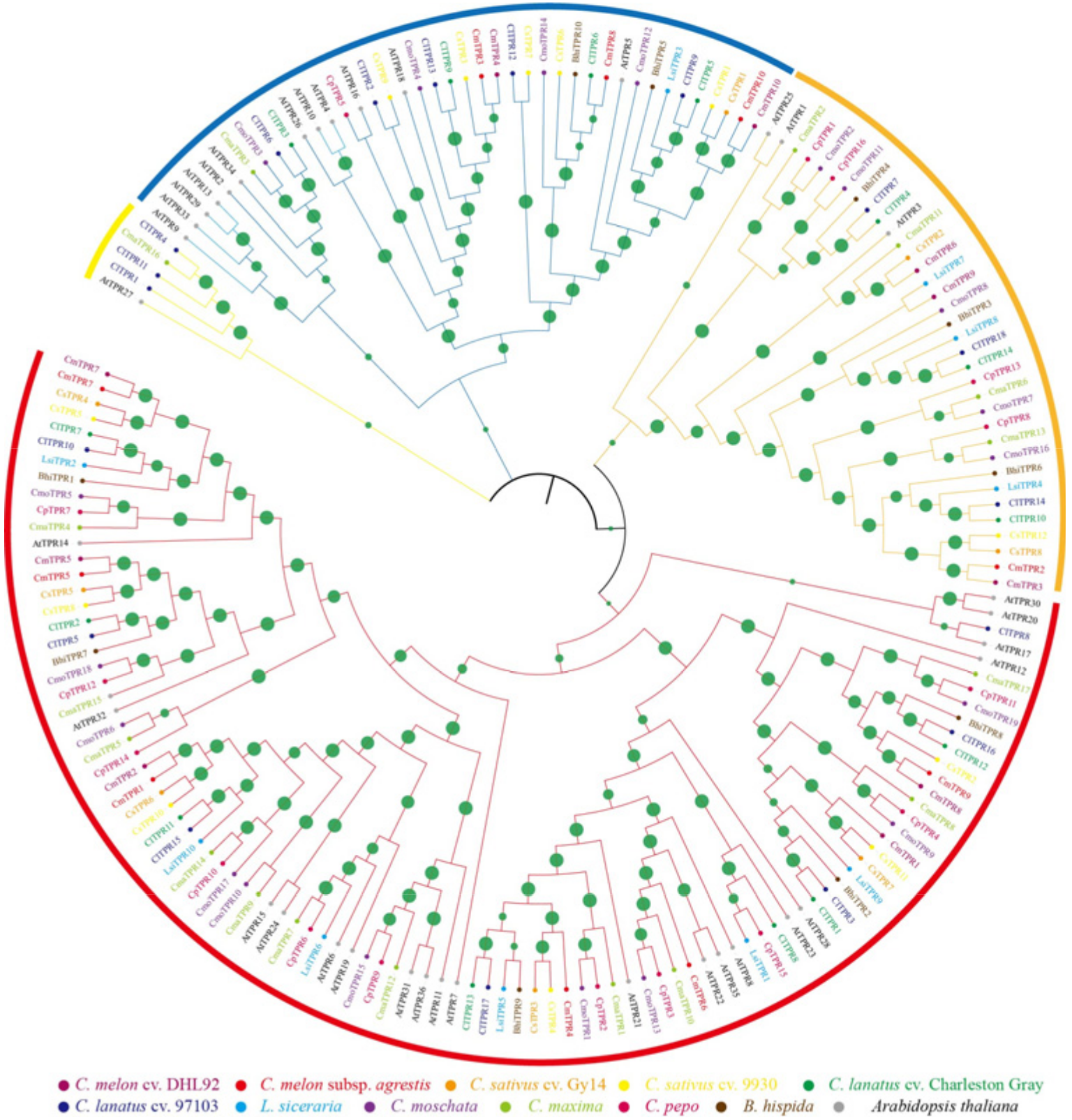
3.2. Phylogenetic Divergence of TPR Genes in Cucurbitaceae
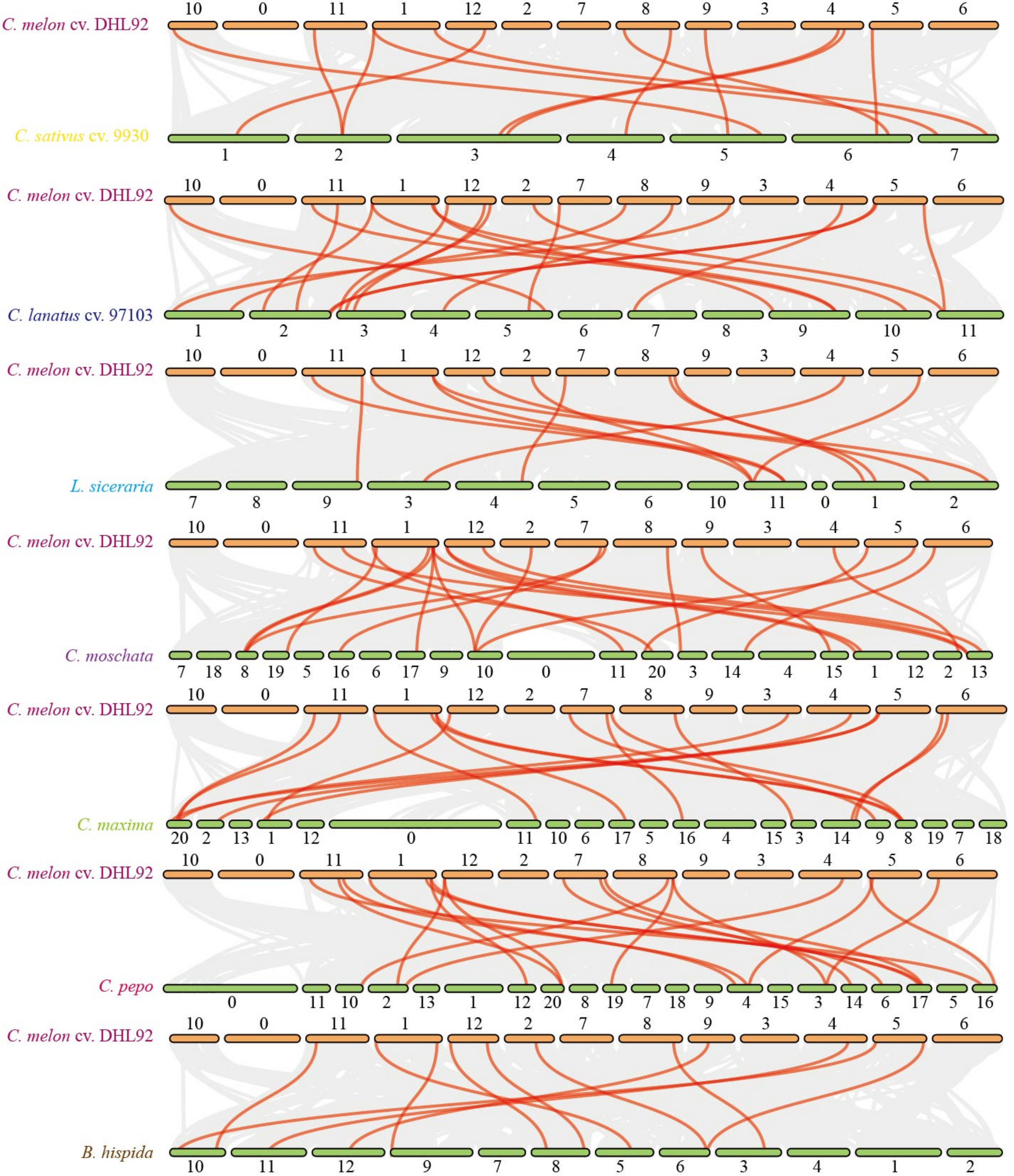
3.3. Collinearity Analysis of TPR Genes among Different Species
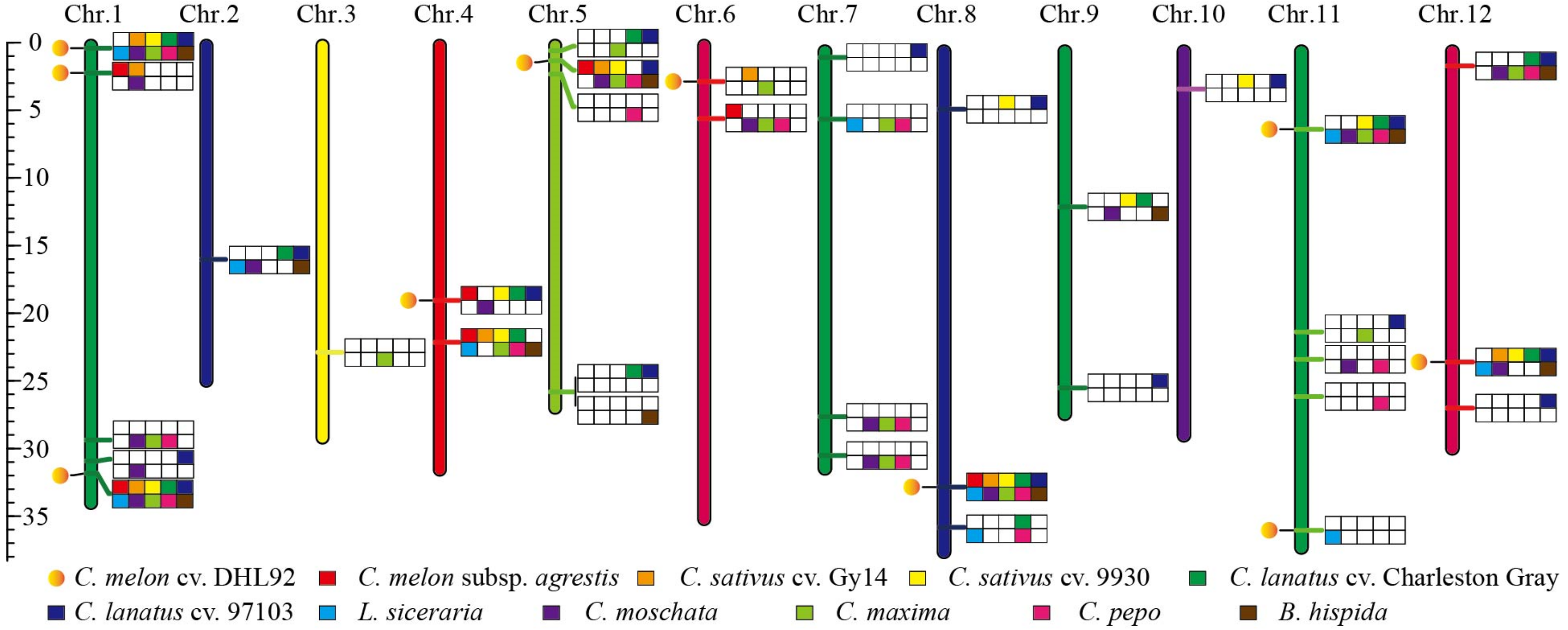
3.4. Construction of Integration Gene Map for TPR Genes
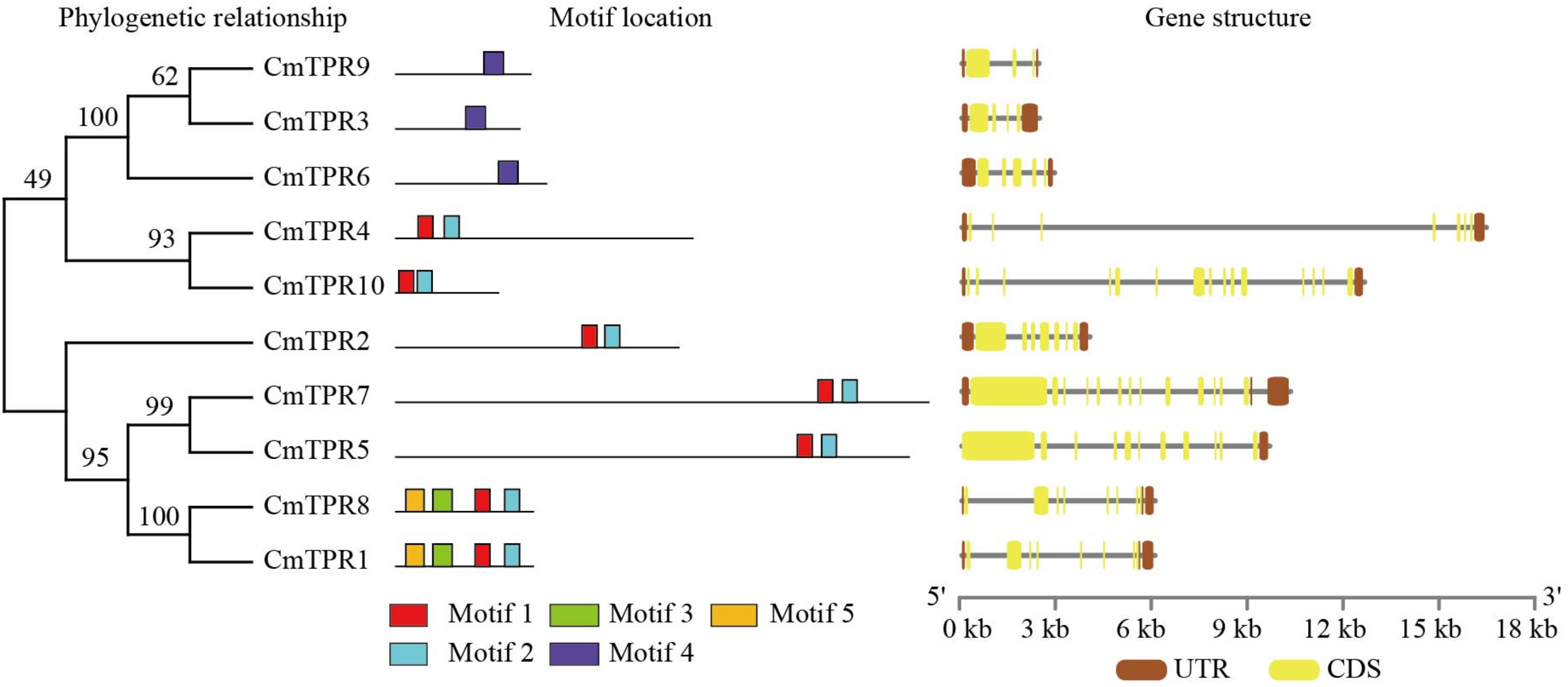
3.5. Phylogenetic Relationship, Gene Structure, and Motif Analysis in Melon
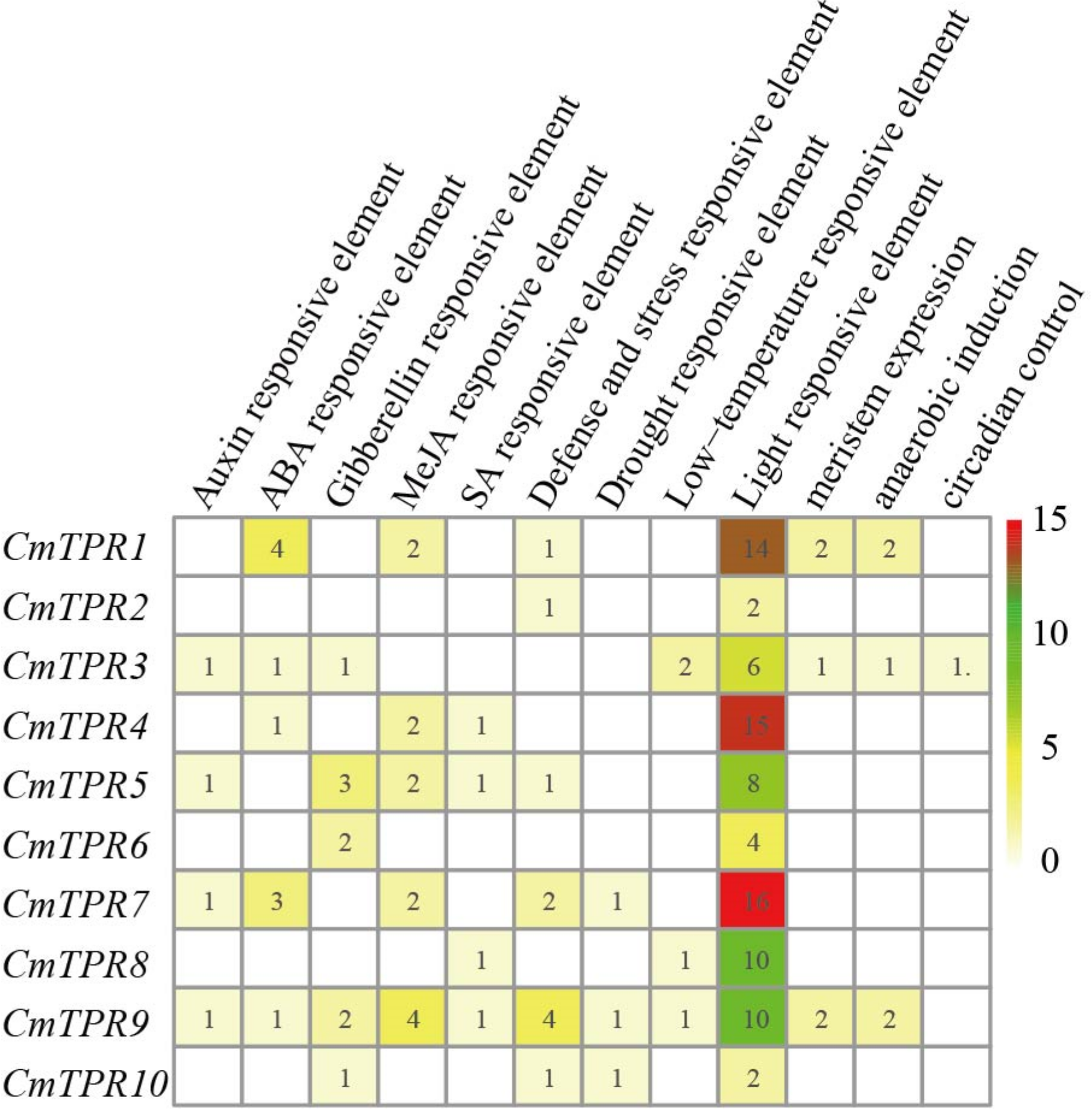
3.6. Analysis of Promoter Cis-Regulator Elements and GO Terms in Melon
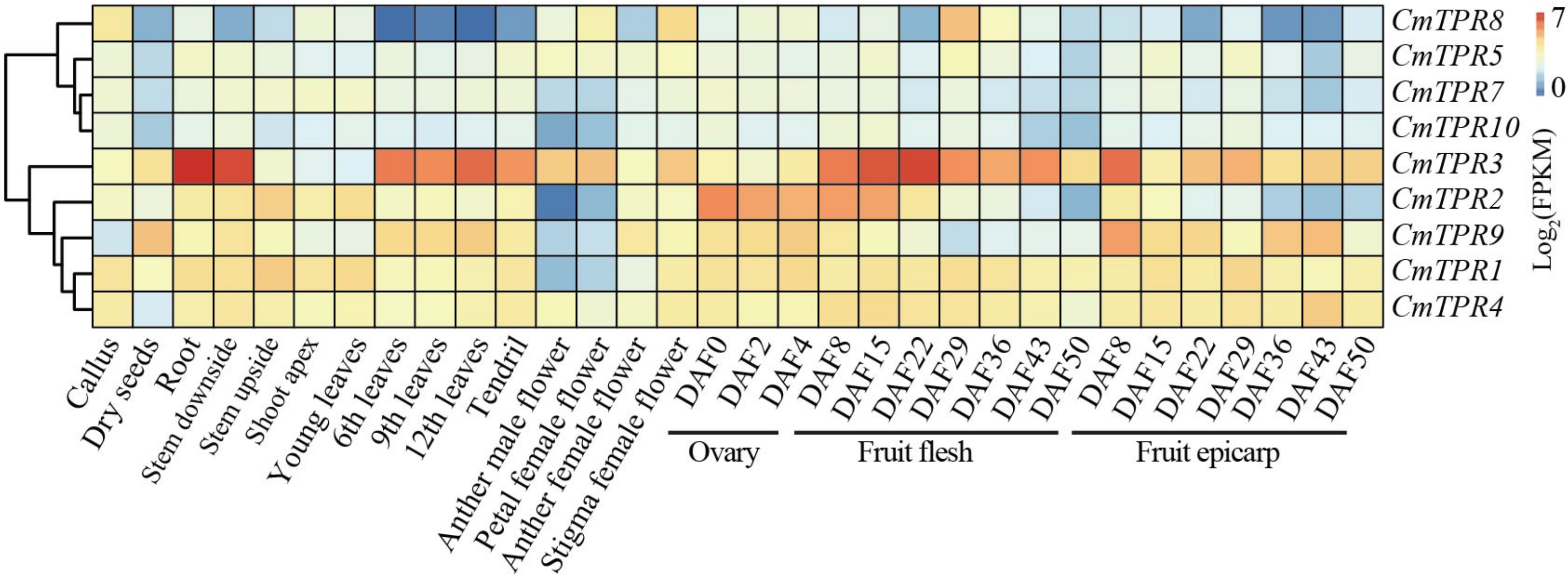
3.7. Tissue-Specific Expression of CmTPR Genes in Melon
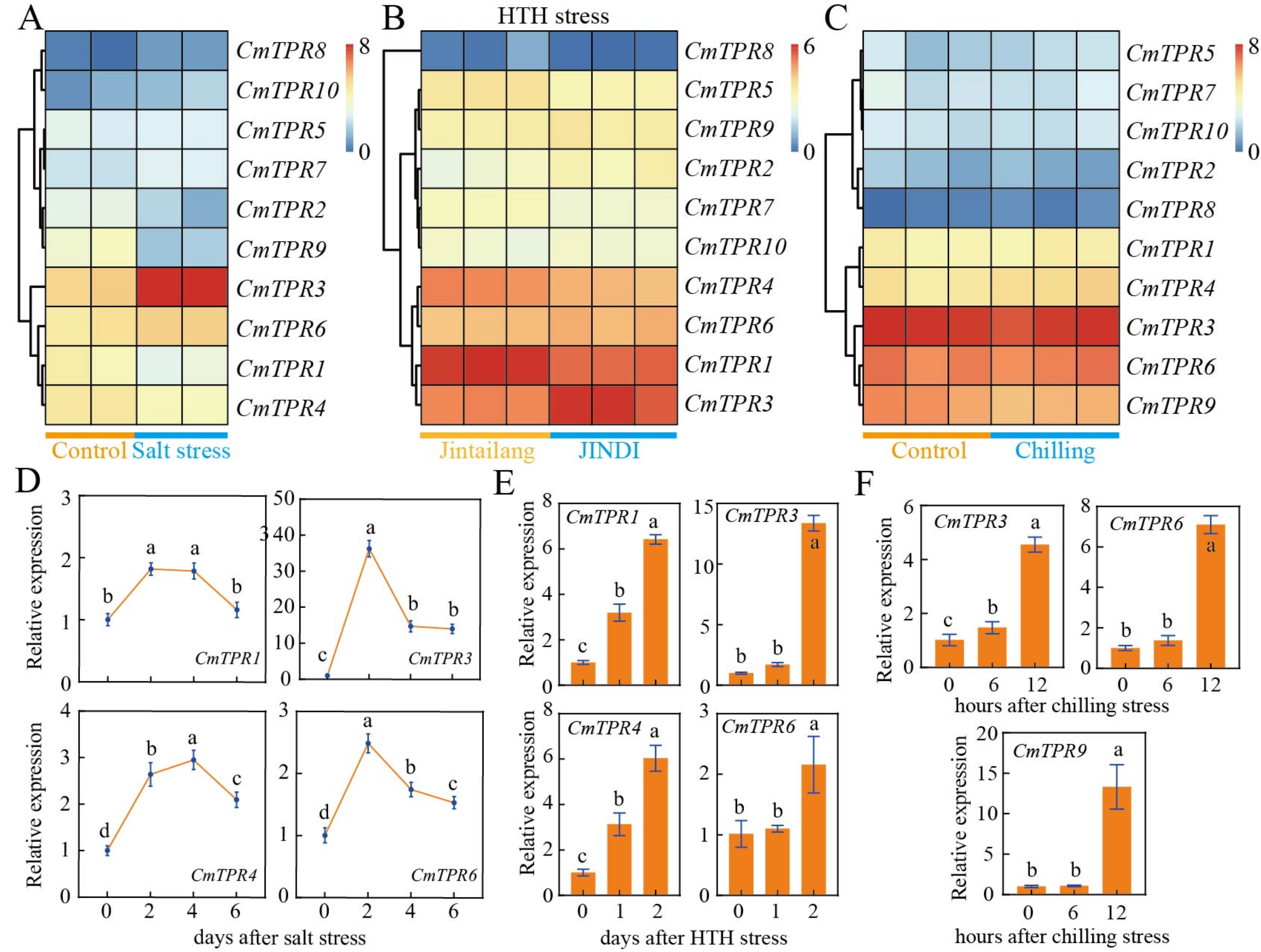
3.8. CmTPR Genes Expression in Response to Multiple Stresses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Xin, T.; Tian, H.; Ma, Y.; Wang, S.; Yang, L.; Li, X.; Zhang, M.; Chen, C.; Wang, H.; Li, H.; et al. Targeted creating new mutants with compact plant architecture using CRISPR/Cas9 genome editing by an optimized genetic transformation procedure in cucurbit plants. Hortic. Res. 2022, 9, uhab086. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Lian, Q.; Zhang, Z.; Fu, Q.; He, Y.; Ma, S.; Ruggieri, V.; Monforte, A.J.; Wang, P.; Julca, I.; et al. A comprehensive genome variation map of melon identifies multiple domestication events and loci influencing agronomic traits. Nat. Genet. 2019, 51, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Davoudi, M.; Song, M.; Zhang, M.; Chen, J.; Lou, Q. Long-distance control of pumpkin rootstock over cucumber scion under drought stress as revealed by transcriptome sequencing and mobile mRNAs identifications. Hortic. Res. 2022, 9, uhab033. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Guo, Y.; Yan, J.; Zhang, Z.; Yuan, L.; Wei, C.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X.; et al. The role of watermelon caffeic acid O-methyltransferase (ClCOMT1) in melatonin biosynthesis and abiotic stress tolerance. Hortic. Res. 2021, 8, 210. [Google Scholar] [CrossRef]
- Zhao, Z.; Dong, Y.; Wang, J.; Zhang, G.; Zhang, Z.; Zhang, A.; Wang, Z.; Ma, P.; Li, Y.; Zhang, X.; et al. Comparative transcriptome analysis of melon (Cucumis melo L.) reveals candidate genes and pathways involved in powdery mildew resistance. Sci. Rep. 2022, 12, 4936. [Google Scholar] [CrossRef] [PubMed]
- Sikorski, R.S.; Boguski, M.S.; Goebl, M.; Hieter, P. A repeating amino acid motif in CDC23 defines a family of proteins and a new relationship among genes required for mitosis and RNA synthesis. Cell 1990, 60, 307–317. [Google Scholar] [CrossRef]
- Wei, K.; Han, P. Comparative functional genomics of the TPR gene family in Arabidopsis, rice and maize. Mol. Breed. 2017, 37, 152. [Google Scholar] [CrossRef]
- Goebl, M.; Yanagida, M. The TPR snap helix: A novel protein repeat motif from mitosis to transcription. Trends Biochem. Sci. 1991, 16, 173–177. [Google Scholar] [CrossRef]
- Haucke, V.; Horst, M.; Schatz, G.; Lithgow, T. The Mas20p and Mas70p subunits of the protein import receptor of yeast mitochondria interact via the tetratricopeptide repeat motif in Mas20p: Evidence for a single hetero-oligomeric receptor. EMBO J. 1996, 15, 1231–1237. [Google Scholar] [CrossRef]
- Tsukahara, F.; Urakawa, I.; Hattori, M.; Hirai, M.; Ohba, K.; Yoshioka, T.; Sakaki, Y.; Muraki, T. Molecular characterization of the mouse mtprd gene, a homologue of human TPRD: Unique gene expression suggesting its critical role in the pathophysiology of Down syndrome. J. Biochem. 1998, 123, 1055–1063. [Google Scholar] [CrossRef]
- Cerveny, L.; Straskova, A.; Dankova, V.; Hartlova, A.; Ceckova, M.; Staud, F.; Stulik, J. Tetratricopeptide repeat motifs in the world of bacterial pathogens: Role in virulence mechanisms. Infect. Immun. 2013, 81, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Groshong, A.M.; Fortune, D.E.; Moore, B.P.; Spencer, H.J.; Skinner, R.A.; Bellamy, W.T.; Blevins, J.S. BB0238, a presumed tetratricopeptide repeat-containing protein, is required during Borrelia burgdorferi mammalian infection. Infect. Immun. 2014, 82, 4292–4306. [Google Scholar] [CrossRef]
- Hirano, T.; Kinoshita, N.; Morikawa, K.; Yanagida, M. Snap helix with knob and hole: Essential repeats in S. pombe nuclear protein nuclear. Cell 1990, 60, 319–328. [Google Scholar] [CrossRef]
- D’Andrea, L.D.; Regan, L. TPR proteins: The versatile helix. Trends Biochem. Sci. 2003, 28, 655–662. [Google Scholar] [CrossRef]
- Goodarzi, M.O.; Xu, N.; Cui, J.; Guo, X.; Chen, Y.I.; Azziz, R. Small glutamine-rich tetratricopeptide repeat-containing protein alpha (SGTA), a candidate gene for polycystic ovary syndrome. Hum. Reprod. 2008, 23, 1214–1219. [Google Scholar] [CrossRef] [PubMed]
- Grizot, S.; Fieschi, F.; Dagher, M.C.; Pebay-Peyroula, E. The active N-terminal region of p67phox. Structure at 1.8 A resolution and biochemical characterizations of the A128V mutant implicated in chronic granulomatous disease. J. Biol. Chem. 2001, 276, 21627–21631. [Google Scholar] [CrossRef] [PubMed]
- Connarn, J.N.; Assimon, V.A.; Reed, R.A.; Tse, E.; Southworth, D.R.; Zuiderweg, E.R.; Gestwicki, J.E.; Sun, D. The molecular chaperone Hsp70 activates protein phosphatase 5 (PP5) by binding the tetratricopeptide repeat (TPR) domain. J. Biol. Chem. 2014, 289, 2908–2917. [Google Scholar] [CrossRef]
- Yan, J.; Wang, J.; Li, Q.; Hwang, J.R.; Patterson, C.; Zhang, H. AtCHIP, a U-box-containing E3 ubiquitin ligase, plays a critical role in temperature stress tolerance in Arabidopsis. Plant Physiol. 2003, 132, 861–869. [Google Scholar] [CrossRef]
- Rosado, A.; Schapire, A.L.; Bressan, R.A.; Harfouche, A.L.; Hasegawa, P.M.; Valpuesta, V.; Botella, M.A. The Arabidopsis tetratricopeptide repeat-containing protein TTL1 is required for osmotic stress responses and abscisic acid sensitivity. Plant Physiol. 2006, 142, 1113–1126. [Google Scholar] [CrossRef]
- Zhou, X.; Zheng, Y.; Cai, Z.; Wang, X.; Liu, Y.; Yu, A.; Chen, X.; Liu, J.; Zhang, Y.; Wang, A. Identification and Functional Analysis of Tomato TPR Gene Family. Int. J. Mol. Sci. 2021, 22, 758. [Google Scholar] [CrossRef]
- Shan, Q.; Liu, M.; Li, R.; Shi, Q.; Li, Y.; Gong, B. γ-Aminobutyric acid (GABA) improves pesticide detoxification in plants. Sci. Total Environ. 2022, 835, 155404. [Google Scholar] [CrossRef]
- Gao, H.; Suo, X.; Zhao, L.; Ma, X.; Cheng, R.; Wang, G.; Zhang, H. Molecular evolution, diversification, and expression assessment of MADS gene family in Setaria italica, Setaria viridis, and Panicum virgatum. Plant Cell Rep. 2023, 42, 1003–1024. [Google Scholar] [CrossRef]
- Li, J.; Liu, J.; Wang, G.; Cha, J.Y.; Li, G.; Chen, S.; Li, Z.; Guo, J.; Zhang, C.; Yang, Y.; et al. A chaperone function of NO CATALASE ACTIVITY1 is required to maintain catalase activity and for multiple stress responses in Arabidopsis. Plant Cell 2015, 27, 908–925. [Google Scholar] [CrossRef]
- Greenboim-Wainberg, Y.; Maymon, I.; Borochov, R.; Alvarez, J.; Olszewski, N.; Ori, N.; Eshed, Y.; Weiss, D. Cross talk between gibberellin and cytokinin: The Arabidopsis GA response inhibitor SPINDLY plays a positive role in cytokinin signaling. Plant Cell 2005, 17, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Xu, W.; Hu, Y.; Huang, J.; Zhao, Y.; Zhang, L.; Huang, C.H.; Ma, H. Phylotranscriptomics in Cucurbitaceae Reveal Multiple Whole-Genome Duplications and Key Morphological and Molecular Innovations. Mol. Plant 2020, 13, 1117–1133. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mas, J.; Benjak, A.; Sanseverino, W.; Bourgeois, M.; Mir, G.; González, V.M.; Hénaff, E.; Câmara, F.; Cozzuto, L.; Lowy, E.; et al. The genome of melon (Cucumis melo L.). Proc. Natl. Acad. Sci. USA 2012, 109, 11872–11877. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Deng, G.; Lian, J.; Garraway, J.; Niu, Y.; Hu, Z.; Yu, J.; Zhang, M. The Chromosome-Scale Genome of Melon Dissects Genetic Architecture of Important Agronomic Traits. iScience 2020, 23, 101422. [Google Scholar] [CrossRef]
- Yang, L.; Koo, D.H.; Li, Y.; Zhang, X.; Luan, F.; Havey, M.J.; Jiang, J.; Weng, Y. Chromosome rearrangements during domestication of cucumber as revealed by high-density genetic mapping and draft genome assembly. Plant J. 2012, 71, 895–906. [Google Scholar] [CrossRef]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef]
- Wu, S.; Wang, X.; Reddy, U.; Sun, H.; Bao, K.; Gao, L.; Mao, L.; Patel, T.; Ortiz, C.; Abburi, V.L.; et al. Genome of ‘Charleston Gray’, the principal American watermelon cultivar, and genetic characterization of 1365 accessions in the U.S. National Plant Germplasm System watermelon collection. Plant Biotechnol. J. 2019, 17, 2246–2258. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, J.; Sun, H.; Salse, J.; Lucas, W.J.; Zhang, H.; Zheng, Y.; Mao, L.; Ren, Y.; Wang, Z.; et al. The draft genome of watermelon (Citrullus lanatus) and resequencing of 20 diverse accessions. Nat. Genet. 2013, 45, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Shamimuzzaman, M.; Sun, H.; Salse, J.; Sui, X.; Wilder, A.; Wu, Z.; Levi, A.; Xu, Y.; Ling, K.S.; et al. The bottle gourd genome provides insights into Cucurbitaceae evolution and facilitates mapping of a Papaya ring-spot virus resistance locus. Plant J. 2017, 92, 963–975. [Google Scholar] [CrossRef]
- Sun, H.; Wu, S.; Zhang, G.; Jiao, C.; Guo, S.; Ren, Y.; Zhang, J.; Zhang, H.; Gong, G.; Jia, Z.; et al. Karyotype Stability and Unbiased Fractionation in the Paleo-Allotetraploid Cucurbita Genomes. Mol. Plant 2017, 10, 1293–1306. [Google Scholar] [CrossRef] [PubMed]
- Montero-Pau, J.; Blanca, J.; Bombarely, A.; Ziarsolo, P.; Esteras, C.; Martí-Gómez, C.; Ferriol, M.; Gómez, P.; Jamilena, M.; Mueller, L.; et al. De novo assembly of the zucchini genome reveals a whole-genome duplication associated with the origin of the Cucurbita genus. Plant Biotechnol. J. 2018, 16, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Xu, Y.; Wang, J.; Liu, W.; Zhou, Q.; Luo, S.; Huang, W.; He, X.; Li, Q.; Peng, Q.; et al. The wax gourd genomes offer insights into the genetic diversity and ancestral cucurbit karyotype. Nat. Commun. 2019, 10, 5158. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Zhang, R.; Yang, X.; Zhu, C.; Li, H.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X. Comparative Analysis of Calcium-Dependent Protein Kinase in Cucurbitaceae and Expression Studies in Watermelon. Int. J. Mol. Sci. 2019, 20, 2527. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Yano, R.; Ariizumi, T.; Nonaka, S.; Kawazu, Y.; Zhong, S.; Mueller, L.; Giovannoni, J.J.; Rose, J.K.C.; Ezura, H. Comparative genomics of muskmelon reveals a potential role for retrotransposons in the modification of gene expression. Commun. Biol. 2020, 3, 432. [Google Scholar] [CrossRef]
- Wang, L.M.; Zhang, L.D.; Chen, J.B.; Huang, D.F.; Zhang, Y.D. Physiological analysis and transcriptome comparison of two muskmelon (Cucumis melo L.) cultivars in response to salt stress. Genet. Mol. Res. 2016, 15, gmr.15038738. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.; Rehman, A.; Li, P.; Chang, L.; Zhang, Y.; Niu, Q. Physiological and Transcriptomic Analysis Reveals the Responses and Difference to High Temperature and Humidity Stress in Two Melon Genotypes. Int. J. Mol. Sci. 2022, 23, 734. [Google Scholar] [CrossRef]
- Diao, Q.; Cao, Y.; Fan, H.; Zhang, Y. Transcriptome analysis deciphers the mechanisms of exogenous nitric oxide action on the response of melon leaves to chilling stress. Biol. Plant. 2020, 64, 465–472. [Google Scholar] [CrossRef]
- Liu, P.; Wang, S.; Wang, X.; Yang, X.; Li, Q.; Wang, C.; Chen, C.; Shi, Q.; Ren, Z.; Wang, L. Genome-wide characterization of two-component system (TCS) genes in melon (Cucumis melo L.). Plant Physiol. Biochem. 2020, 151, 197–213. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Kherawat, B.S.; Singh, A.; Dey, P.; Routray, S.; Mohapatra, C.; Saha, D.; Ram, C.; Siddique, K.H.M.; Kumar, A.; et al. Genome-Wide Analysis and Characterization of the Proline-Rich Extensin-like Receptor Kinases (PERKs) Gene Family Reveals Their Role in Different Developmental Stages and Stress Conditions in Wheat (Triticum aestivum L.). Plants 2022, 11, 496. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Guan, Y.; Weng, Y.; Liao, B.; Tong, L.; Hao, Z.; Chen, J.; Shi, J.; Cheng, T. Genome-wide identification of the NAC gene family and its functional analysis in Liriodendron. BMC Plant Biol. 2023, 23, 415. [Google Scholar] [CrossRef]
- Li, G.L.; Tang, L.L.; He, Y.H.; Xu, Y.Y.; Bendahmane, A.; Garcia-Mas, J.; Lin, T.; Zhao, G.G. The haplotype-resolved T2T reference genome highlights structural variation underlying agronomic traits of melon. Hortic. Res. 2023, 10, uhad182. [Google Scholar] [CrossRef]
- Tang, H.; Bowers, J.E.; Wang, X.; Ming, R.; Alam, M.; Paterson, A.H. Synteny and collinearity in plant genomes. Science 2008, 320, 486–488. [Google Scholar] [CrossRef]
- Li, H.; Wen, X.; Huang, X.; Wei, M.; Chen, H.; Yu, Y.; Dai, S. Genome-Wide Identification and Characterization of TCP Gene Family Members in Melastoma candidum. Molecules 2022, 27, 9036. [Google Scholar] [CrossRef]
- Xu, P.; Wang, Y.; Sun, F.; Wu, R.; Du, H.; Wang, Y.; Jiang, L.; Wu, X.; Wu, X.; Yang, L.; et al. Long-read genome assembly and genetic architecture of fruit shape in the bottle gourd. Plant J. 2021, 107, 956–968. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Wang, Q.; Zheng, Y.; Guo, J.; Yuan, S.; Fu, A.; Bai, C.; Zhao, X.; Zheng, S.; Wen, C.; et al. Cucurbitaceae genome evolution, gene function, and molecular breeding. Hortic. Res. 2022, 9, uhab057. [Google Scholar] [CrossRef] [PubMed]
- Eickbush, T.H. Mobile introns: Retrohoming by complete reverse splicing. Curr. Biol. 1999, 9, R11–R14. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.; Xu, L.; Chai, P.; Peng, J.; Devarkar, S.C.; Pyle, A.M. Structures of a mobile intron retroelement poised to attack its structured DNA target. Science 2022, 378, 627–634. [Google Scholar] [CrossRef]
- Wen, Z.; Li, M.; Meng, J.; Miao, R.; Liu, X.; Fan, D.; Lv, W.; Cheng, T.; Zhang, Q.; Sun, L. Genome-Wide Identification of the MAPK and MAPKK Gene Families in Response to Cold Stress in Prunus mume. Int. J. Mol. Sci. 2023, 24, 8829. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Jia, X.; Yang, Z.; Fu, Q.; Yang, H.; Xu, X. Genome-Wide Identification of PEBP Gene Family in Solanum lycopersicum. Int. J. Mol. Sci. 2023, 24, 9185. [Google Scholar] [CrossRef]
- Lu, P.; Magwanga, R.O.; Guo, X.; Kirungu, J.N.; Lu, H.; Cai, X.; Zhou, Z.; Wei, Y.; Wang, X.; Zhang, Z.; et al. Genome-Wide Analysis of Multidrug and Toxic Compound Extrusion (MATE) Family in Gossypium raimondii and Gossypium arboreum and Its Expression Analysis Under Salt, Cadmium, and Drought Stress. G3-Genes Genomes Genet. 2018, 8, 2483–2500. [Google Scholar] [CrossRef]
- Chen, S.; Zhong, K.; Li, Y.; Bai, C.; Xue, Z.; Wu, Y. Evolutionary Analysis of the Melon (Cucumis melo L.) GH3 Gene Family and Identification of GH3 Genes Related to Fruit Growth and Development. Plants 2023, 12, 1382. [Google Scholar] [CrossRef]
- Schapire, A.L.; Valpuesta, V.; Botella, M.A. TPR Proteins in Plant Hormone Signaling. Plant Signal Behav. 2006, 1, 229–230. [Google Scholar] [CrossRef]
- Gray, W.M.; Muskett, P.R.; Chuang, H.W.; Parker, J.E. Arabidopsis SGT1b is required for SCF(TIR1)-mediated auxin response. Plant Cell 2003, 15, 1310–1319. [Google Scholar] [CrossRef]
- Qin, F.; Kodaira, K.S.; Maruyama, K.; Mizoi, J.; Tran, L.S.; Fujita, Y.; Morimoto, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. SPINDLY, a negative regulator of gibberellic acid signaling, is involved in the plant abiotic stress response. Plant Physiol. 2011, 157, 1900–1913. [Google Scholar] [CrossRef]
- Niu, Y.; Chen, M.; Chen, X.; Ma, Y.; Xu, Z.; Li, L. Characterization of Ethylene Receptors and Their Interactions with GmTPR—A Novel Tetratricopeptide Repeat Protein (TPR) in Soybean (Glycine max L.). J. Integr. Agric. 2013, 12, 571–581. [Google Scholar] [CrossRef]
- Lakhssassi, N.; Doblas, V.G.; Rosado, A.; Esteban Del Valle, A.; Posé, D.; Jimenez, A.J.; Castillo, A.G.; Valpuesta, V.; Borsani, O.; Botella, M.A. The Arabidopsis thaliana TETRATRICO PEPTIDE THIOREDOXIN-LIKE gene family is required for osmotic stress tolerance and male sporogenesis. Plant Physiol. 2012, 158, 1252–1266. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, Y.; Qi, J.; Chi, Y.; Fan, B.; Yu, J.Q.; Chen, Z. E3 ubiquitin ligase CHIP and NBR1-mediated selective autophagy protect additively against proteotoxicity in plant stress responses. PLOS Genet. 2014, 10, e1004478. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | No. of Genes | No. of Chr. | No. of Amino Acids | Molecular Weight | Theoretical pI | Total Number of Atoms | Instability Index | Aliphatic Index | GRAVY |
|---|---|---|---|---|---|---|---|---|---|
| Cucumis melo L. cv. DHL92 | 10 | 7 | 270–1388 | 30,756.79–153,512.86 | 4.24–8.76 | 4320–21,188 | 23.64–62.19 | 62.6–94.1 | −0.755–0.148 |
| Cucumis melo subsp. agrestis | 10 | 8 | 281–1373 | 32,007.34–151,908.17 | 5.31–8.76 | 4500–20,967 | 27.2–50.84 | 62.87–94.1 | −0.754–0.148 |
| Cucumis sativus L. cv. Gy14 | 8 | 5 | 319–1383 | 34,813.22–152,300.88 | 4.79–8.76 | 4784–21,068 | 23.31–52.69 | 66.83–94.68 | −0.68–0.11 |
| Cucumis. sativus L. var. sativus var. 9930 | 12 | 7 | 281–1368 | 32,050.38–150,585.73 | 5.41–8.76 | 4499–20,818 | 23.31–50.92 | 64.42–92.32 | −0.7–0.144 |
| Citrullus lanatus subsp. vulgaris cv. Charleston Gray | 14 | 7 | 269–1435 | 29,612.28–157,680 | 4.8–8.74 | 4078–21,757 | 22.73–61.26 | 62.85–90.38 | −0.687–0.186 |
| Citrullus lanatus subsp. vulgaris cv. 97103 | 18 | 9 | 269–1358 | 29,612.28–152,201.83 | 4.8–8.59 | 4078–20,992 | 22.8–61.26 | 63.36–90.22 | −0.687–0.195 |
| Lagenaria siceraria var. USVL1VR-Ls | 10 | 6 | 272–1384 | 29,403.12–151,284.88 | 4.63–8.95 | 4060–20,858 | 21.74–55.51 | 58.29–96.64 | −0.68–0.015 |
| Cucurbita moschata var. Rifu | 19 | 13 | 245–1376 | 27,266.81–151,323.17 | 4.62–9.19 | 3817–20,874 | 24.78–67.92 | 62.64–92.94 | −0.667–0.11 |
| Cucurbita maxima var. Rimu | 17 | 11 | 198–1375 | 21,697.96–150,981.06 | 4.02–9.17 | 2965–20,841 | 22.02–56.92 | 63.8–88.25 | −0.702–0.258 |
| Cucurbita pepo subsp. pepo | 16 | 12 | 307–1372 | 32,914.14–150,709.35 | 4.89–9.3 | 4514–20,762 | 21.76–60.48 | 61.83–90.18 | −0.687–0.272 |
| Benincasa hispida var. B227 | 10 | 8 | 253–1370 | 27,776.22–149,982.2 | 4.7–7.1 | 3847–20,668 | 22.72–60.66 | 63.77–91.71 | −0.683–0.167 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Meng, Y.; Ding, F.; Yang, K.; Wang, C.; Zhang, H.; Jin, H. Comparative Analysis of TPR Gene Family in Cucurbitaceae and Expression Profiling under Abiotic Stress in Cucumis melo L. Horticulturae 2024, 10, 83. https://doi.org/10.3390/horticulturae10010083
Wang S, Meng Y, Ding F, Yang K, Wang C, Zhang H, Jin H. Comparative Analysis of TPR Gene Family in Cucurbitaceae and Expression Profiling under Abiotic Stress in Cucumis melo L. Horticulturae. 2024; 10(1):83. https://doi.org/10.3390/horticulturae10010083
Chicago/Turabian StyleWang, Shuoshuo, Yuchen Meng, Fei Ding, Kuo Yang, Chuang Wang, Hengjia Zhang, and Han Jin. 2024. "Comparative Analysis of TPR Gene Family in Cucurbitaceae and Expression Profiling under Abiotic Stress in Cucumis melo L." Horticulturae 10, no. 1: 83. https://doi.org/10.3390/horticulturae10010083
APA StyleWang, S., Meng, Y., Ding, F., Yang, K., Wang, C., Zhang, H., & Jin, H. (2024). Comparative Analysis of TPR Gene Family in Cucurbitaceae and Expression Profiling under Abiotic Stress in Cucumis melo L. Horticulturae, 10(1), 83. https://doi.org/10.3390/horticulturae10010083