
Exploring MicroRNAs Associated with Pomegranate Pistil Development: An Identification and Analysis Study

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Sequencing and Data Analysis
2.2.1. Library Preparation and Sequencing
2.2.2. Comparison and Analysis of Raw Data
2.2.3. Identification of Conservative miRNA and Novel miRNA
2.2.4. miRNA Expression and Differential Analysis
2.2.5. Prediction and Enrichment Analysis of miRNA Target Genes
2.2.6. Correlation Analysis of Sequencing Results
2.2.7. qRT-PCR Verification of Sequencing Results
3. Results and Analysis
3.1. Sequencing Results
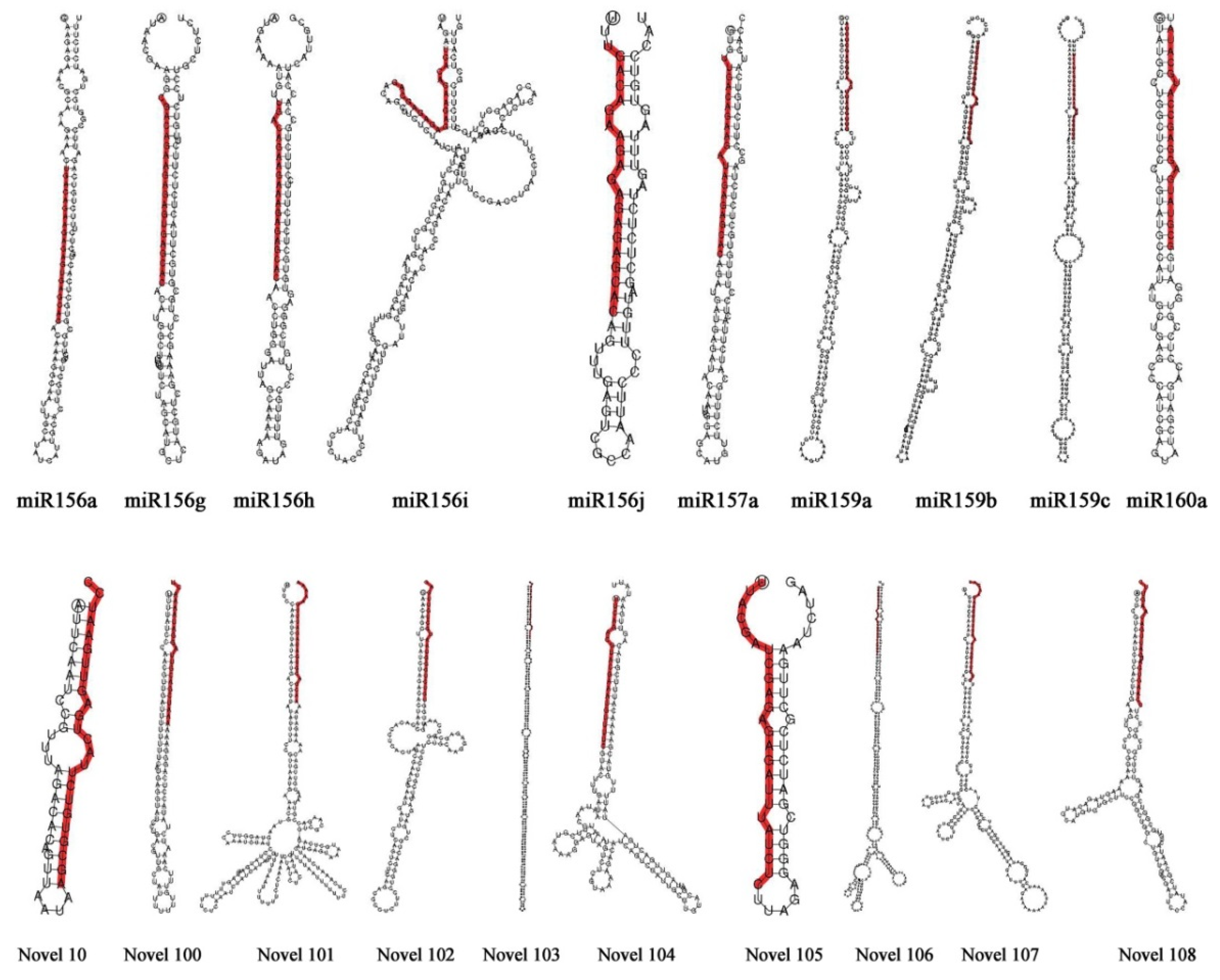
3.2. Identification and Analysis of Conserved miRNAs and Novel miRNAs
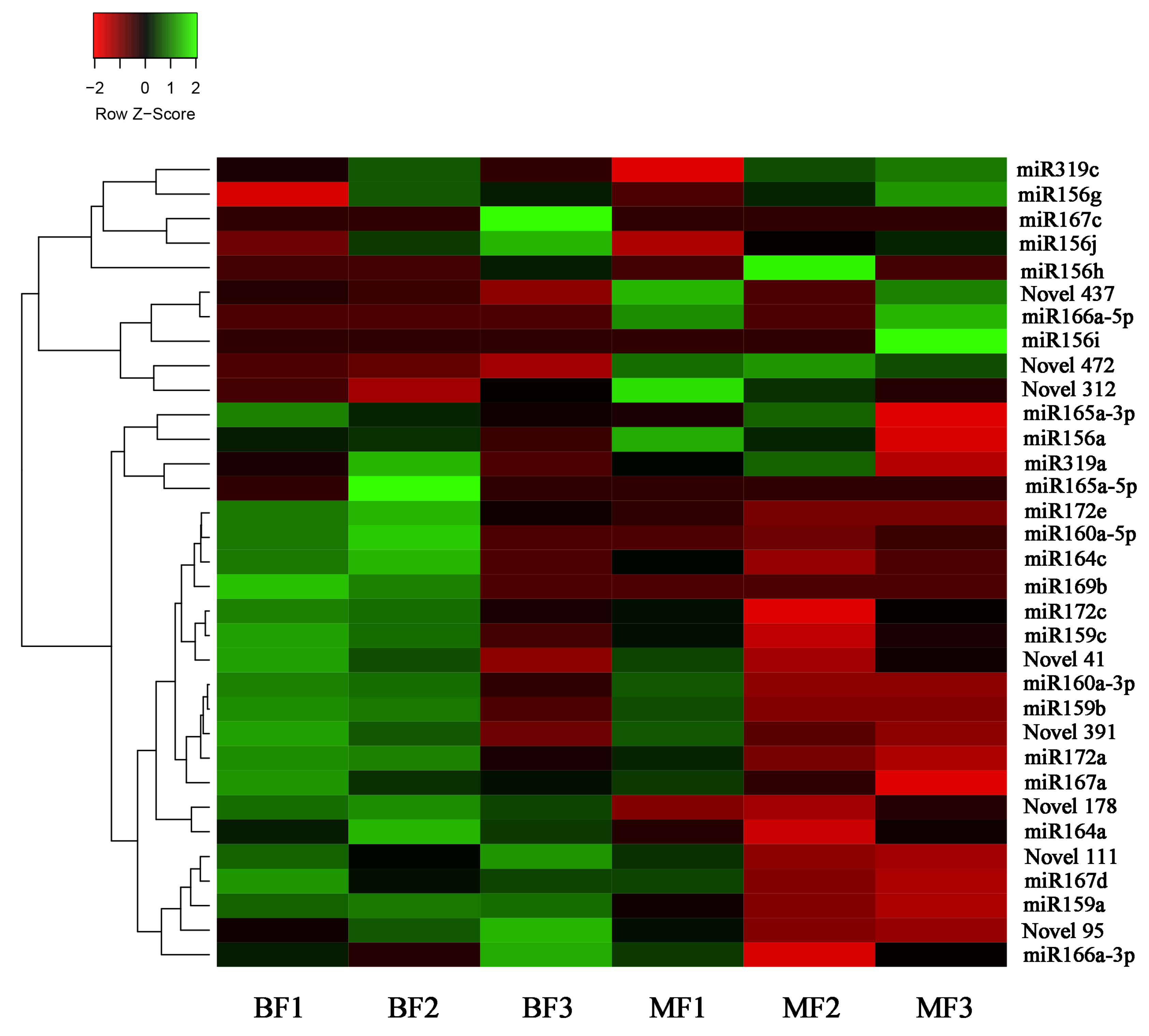
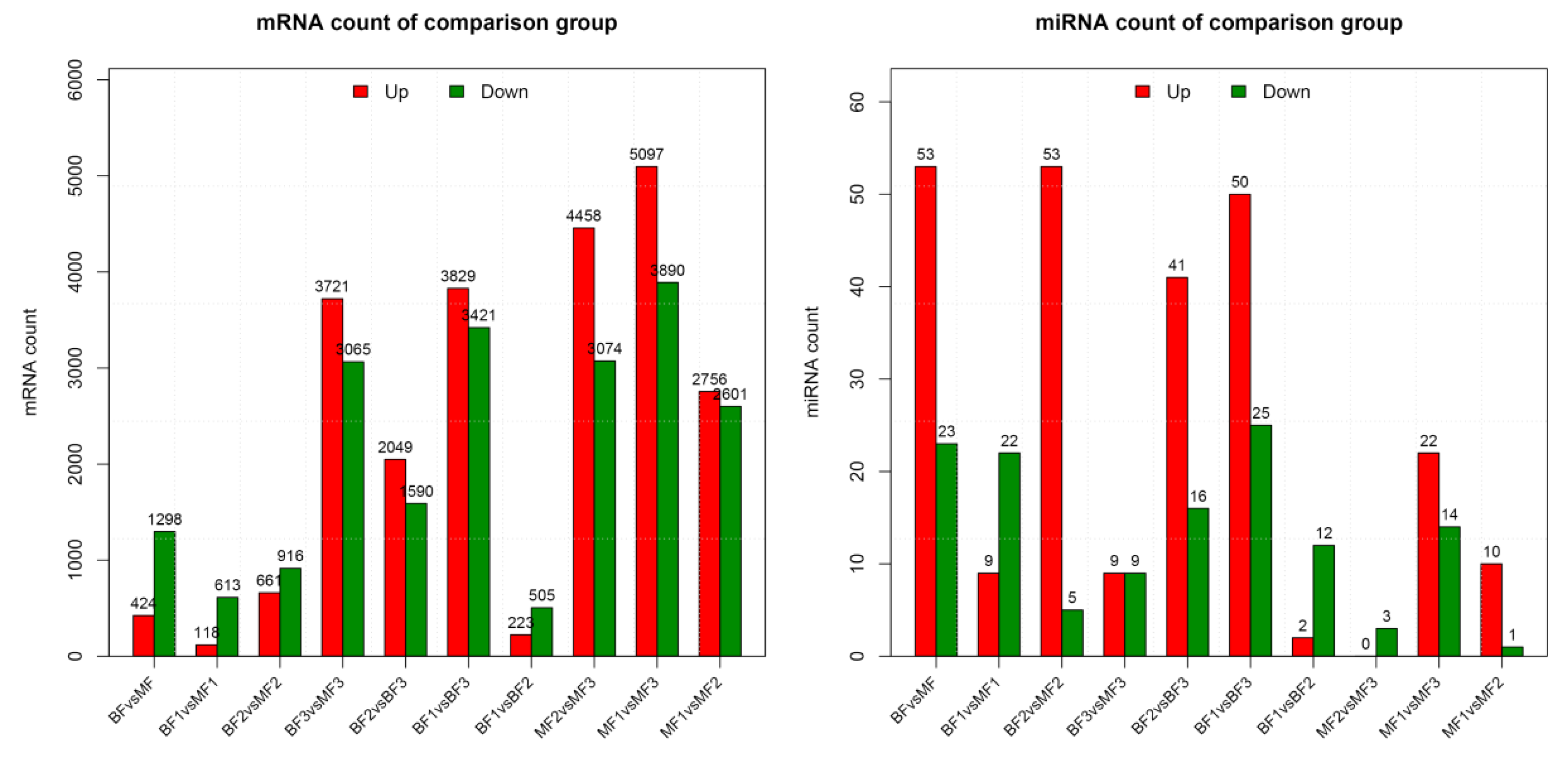
3.3. miRNA Differential Expression Analysis
3.4. miRNA Target Gene Prediction
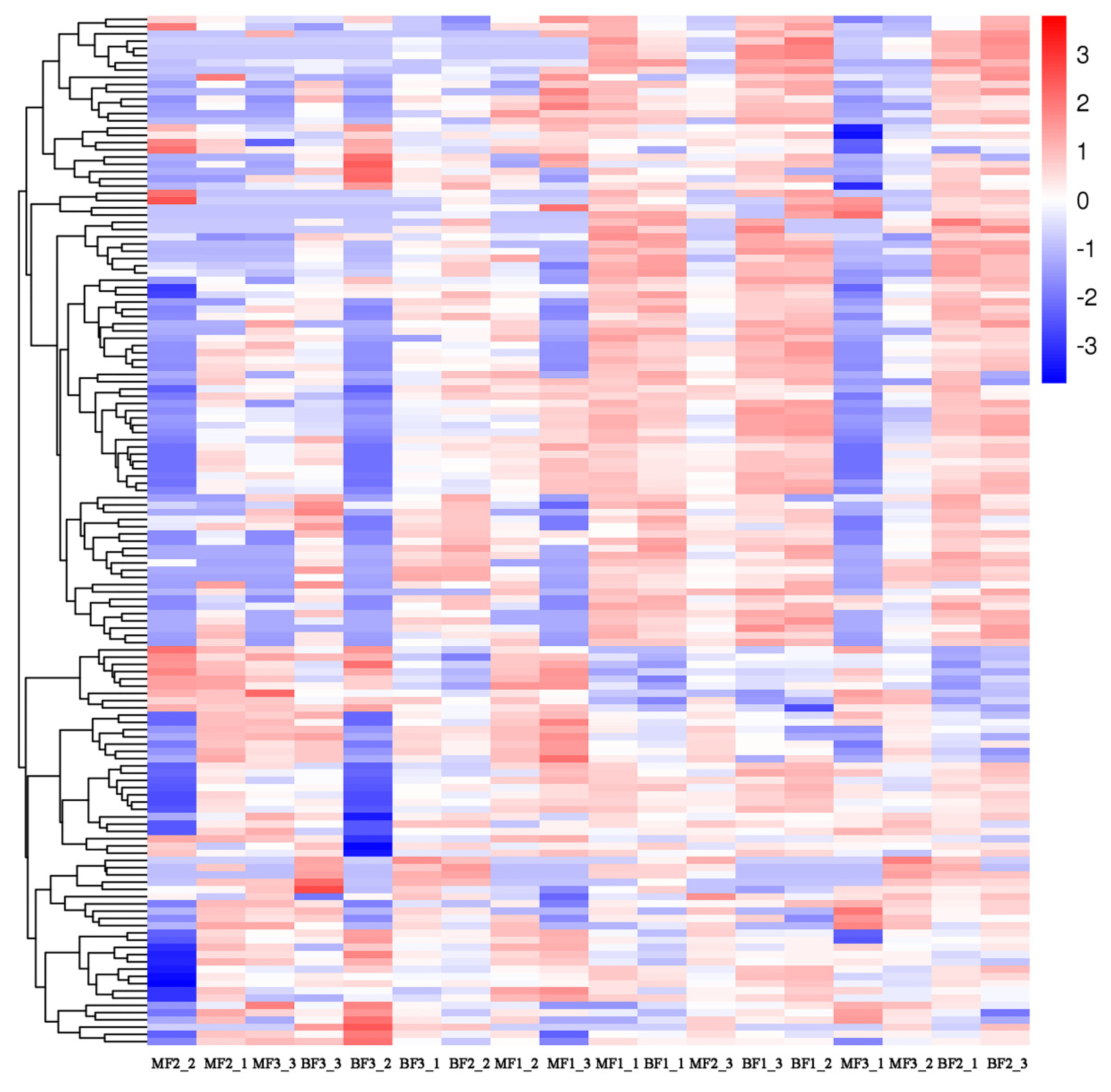
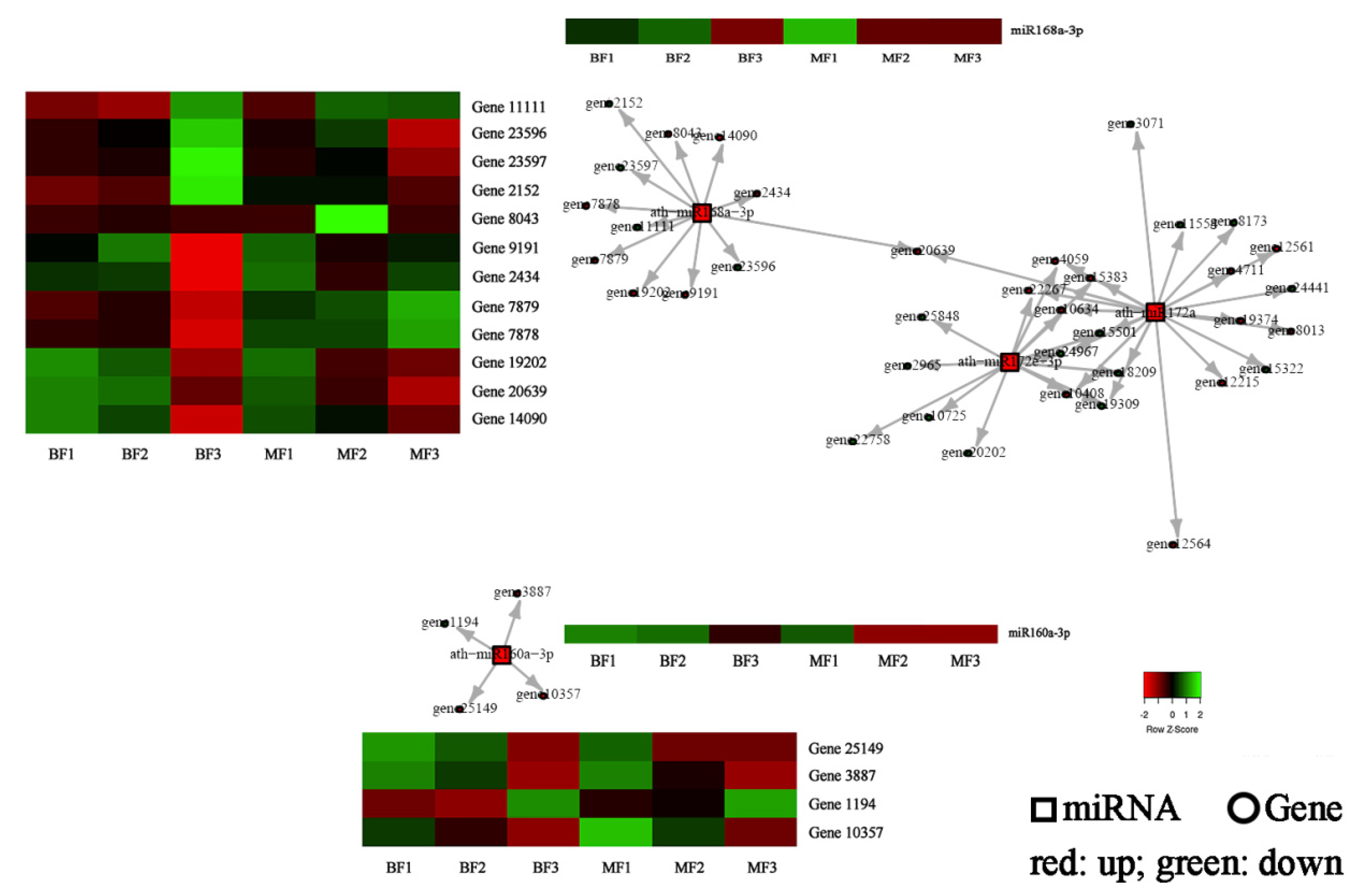
3.5. Correlation Analysis of miRNAs and mRNAs
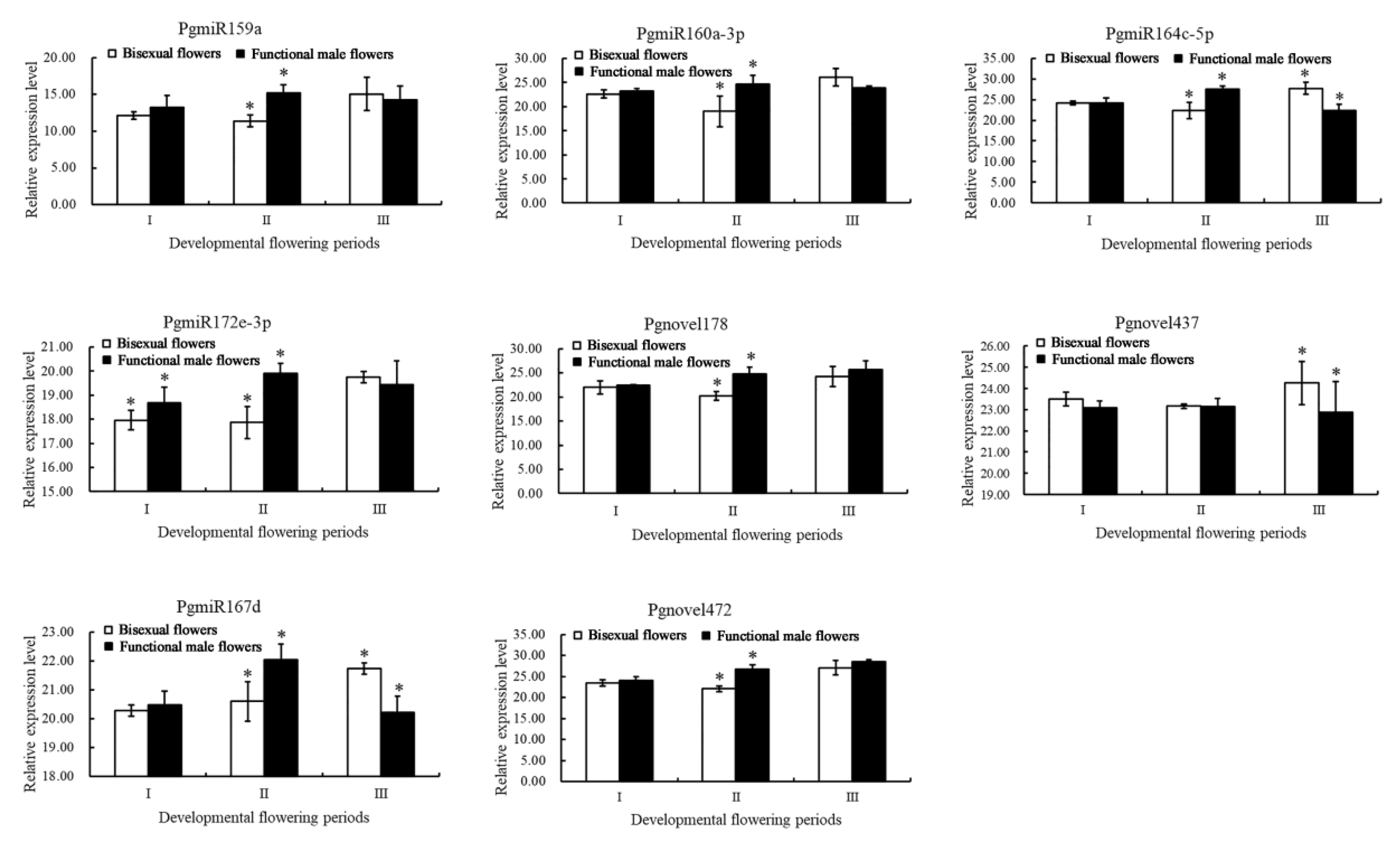
3.6. qRT-PCR Validation of Differential Transcripts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Origin, Biogenesis, and Activity of Plant MicroRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed]
- Arribas-Hernández, L.; Kielpinski, L.J.; Brodersen, P. mRNA decay of most Arabidopsis miRNA targets requires slicer activity of AGO1. Plant Physiol. 2016, 171, 2620–2632. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.K. microRNAs databases: Developmental methodologies, structural and functional annotations. Interdiscip. Sci. 2017, 9, 357–377. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, B.J.; Weinstein, E.G.; Rhoades, M.W.; Bartel, B.; Bartel, D.P. MicroRNAs in plants. Genes Dev. 2002, 16, 1616–1626. [Google Scholar] [CrossRef]
- Khraiwesh, B.; Zhu, J.K.; Zhu, J.H. Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochim. Biophys. Acta(BBA)-ene Regul. Mech. 2012, 1819, 137–148. [Google Scholar] [CrossRef]
- Petrella, R.; Cucinotta, M.; Mendes, M.A.; Underwood, C.J.; Colombo, L. The emerging role of small RNAs in ovule development, a kind of magic. Plant Reprod. 2021, 34, 335–351. [Google Scholar] [CrossRef]
- Araki, S.; Le, N.T.; Koizumi, K.; Villar-Briones, A.; Nonomura, K.-I.; Endo, M.; Inoue, H.; Saze, H.; Komiya, R. miR2118-dependent U-rich phasiRNA production in rice anther wall development. Nat. Commun. 2020, 11, 3115. [Google Scholar] [CrossRef]
- Wu, M.F.; Tian, Q.; Reed, J.W. Arabidopsis microRNA167 controls patterns of ARF6 and ARF8 expression, and regulates both female and male reproduction. Development 2006, 133, 4211–4218. [Google Scholar] [CrossRef]
- Tan, C.; Zhang, Z.M.; Liu, H.J.; Gao, J.; Rong, Y.Z.; Pan, G.T. Advances on microRNAs regulated flower development of higher plant. J. Agric. Biotechnol. 2011, 19, 938–952. [Google Scholar]
- Schwab, R.; Palatnik, J.F.; Riester, M.; Schommer, C.; Schmid, M.; Weigel, D. Specific effects of microRNAs on the plant transcriptome. Dev. Cell 2005, 8, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.; Saedler, H.; Huijser, P. A new family of DNA binding proteins includes putative transcriptional regulators of theAntirrhinum majus floral meristem identity geneSQUAMOSA. Mol. Genet. Genom. 1996, 250, 7–16. [Google Scholar] [CrossRef]
- Sun, C.; Zhao, Q.; Liu, D.D.; You, C.X.; Hao, Y.J. Ectopic expression of the apple Md-miRNA156h gene regulates flower and fruit development in Arabidopsis. Plant Cell 2013, 112, 343–351. [Google Scholar] [CrossRef]
- Yang, C.W. MiR164 is Required for Tomato Flower Initiation and Fruit Development. Master’s Thesis, Chongqing University, Chongqing, China, 2012. [Google Scholar]
- Guo, C.; Jiang, Y.; Shi, M.; Wu, X.; Wu, G. ABI5 acts downstream of miR159 to delay vegetative phase change in Arabidopsis. New Phytol. 2021, 231, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Y.; Yan, M.; Liu, C.; Yuan, Z. BELL1 interacts with CRABS CLAW and INNER NO OUTER to regulate ovule and seed development in pomegranate. Plant Physiol. 2023, 191, 1066–1083. [Google Scholar] [CrossRef] [PubMed]
- Wen, M.; Shen, Y.; Shi, S.; Tang, T. miREvo: An integrative microRNA evolutionary analysis platform for next-generation sequencing experiments. BMC Bioinform. 2012, 13, 140. [Google Scholar] [CrossRef]
- Friedländer, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2011, 40, 37–52. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, J.; Li, Z.; Li, X.; Hu, X.; Huang, Y.; Zhao, X.; Liang, C.; Wang, Y.; Sun, L.; et al. Integrated Profiling of MicroRNAs and mRNAs: MicroRNAs Located on Xq27.3 Associate with Clear Cell Renal Cell Carcinoma. PLoS ONE 2010, 5, e15224. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Wu, H.J.; Ma, Y.-K.; Chen, T.; Wang, M.; Wang, X.J. PsRobot: A web-based plant small RNA meta-analysis toolbox. Nucleic Acids Res. 2012, 40, 22–28. [Google Scholar] [CrossRef]
- Chen, C.; Ridzon, D.A.; Broomer, A.J.; Zhou, Z.; Lee, D.H.; Nguyen, J.T.; Barbisin, M.; Xu, N.L.; Mahuvakar, V.R.; Andersen, M.R.; et al. Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Res. 2005, 33, e179. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Hajkova, P.; Barton, S.C.; Lao, K.; Surani, M.A. MicroRNA expression profiling of single whole embryonic stem cells. Nucleic Acids Res. 2006, 34, e9. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Mi, S.; Cai, T.; Hu, Y.; Chen, Y.; Hodges, E.; Ni, F.; Wu, L.; Li, S.; Zhou, H.; Long, C.; et al. Sorting of small RNAs into Arabidopsis Argonaute complexes is directed by the 5′ terminal nucleotide. Cell 2008, 133, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Jones-Rhoades, M.W.; Barte, D.P. Computational identification of plant microRNAs and their targets, including a stress-induced miRNA. Mol. Cell 2004, 14, 787–799. [Google Scholar] [CrossRef]
- Jones-Rhoades, M.W.; Bartel, D.P.; Bartel, B. MicroRNAs and their regulatory roles in plants. Annu. Rev. Plant Biol. 2006, 57, 19–53. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Y.; Zhao, X.; Yan, M.; Ren, Y.; Yuan, Z. ARF6s identification and function analysis provide insights into flower development of Punica granatum L. Front. Plant Sci. 2022, 13, 833747. [Google Scholar] [CrossRef]
- Kurihara, Y.; Watanabe, Y. Arabidopsis micro-RNA biogenesis through Dicer-like 1 protein functions. Proc. Natl. Acad. Sci. USA 2004, 101, 12753–12758. [Google Scholar] [CrossRef]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef]
- Allen, R.S.; Li, J.; Stahle, M.I.; Dubroué, A.; Gubler, F.; Millar, A.A. Genetic analysis reveals functional redundancy and the major target genes of Arabidopsis miR159 family. Proc. Natl. Acad. Sci. USA 2007, 104, 16371–16376. [Google Scholar] [CrossRef] [PubMed]
- Aukerman, M.J.; Sakai, H. Regulation of flowering time and floral organ identity by a microRNA and its APETALA2-like target genes. Plant Cell 2003, 15, 2730–2741. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.W.; Yang, X.H.; Li, F.; Deng, Y.T. Advances in miRNA-mediated growth and development regulation in horticultural crops. Acta Hortic. Sin. 2022, 49, 1145–1161. [Google Scholar]
- Palatnik, J.F.; Wollmann, H.; Schommer, C.; Schwab, R.; Boisbouvier, J.; Rodriguez, R.; Warthmann, N.; Allen, E.; Dezulian, T.; Huson, D.; et al. Sequence and expression differences underlie functional specialization of Arabidopsis MicroRNAs miR159 and miR319. Dev. Cell 2007, 13, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Poethig, R.S. Temporal regulation of shoot development in Arabidopsis thaliana by Mir156 and its target SPL3. Development 2006, 133, 3539–3547. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.-W.; Weigel, D.; Poethig, R.S. The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Tu, L.; Wang, L.; Hu, H.; Xu, J.; Zhang, X. MicroRNA 157-targeted SPL genes regulate floral organ size and ovule production in cotton. BMC Plant Biol. 2017, 17, 7. [Google Scholar] [CrossRef]
- Duan, Z.X. Expression Pattern and Functional Analysis of microRNA Peu-miR156j and Peu-miR169 from Populus euphratica. Master’s Thesis, Beijing Forestry University, Beijing, China, 2012. [Google Scholar]
- Chen, G.; Li, J.; Liu, Y.; Zhang, Q.; Gao, Y.; Fang, K.; Cao, Q.; Qin, L.; Xing, Y. Roles of the GA-mediated SPL gene family and miR156 in the floral development of Chinese chestnut (Castanea mollissima). Int. J. Mol. Sci. 2019, 20, 1577. [Google Scholar] [CrossRef]
- Liu, X.D.; Zhang, H.; Zhao, Y.; Feng, Z.Y.; Li, Q.; Yang, H.Q.; Luan, S.; Li, J.M.; He, Z.H. Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 15485–15490. [Google Scholar] [CrossRef]
- Wu, S.C. Functional Characterization of mir160 in Cotton Ovule Development. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2016. [Google Scholar]
- Bai, Y.H.; Wang, W.R.; Dong, T.Y.; Guan, L.; Su, Z.W.; Jia, H.F.; Fang, J.G.; Wang, C. vvi-miR160s in mediating vvARF18 response to gibberellin regulation of grape seed development. Sci. Agric. Sin. 2020, 53, 1890–1903. [Google Scholar]
- Hendelman, A.; Buxdorf, K.; Stav, R.; Kravchik, M.; Arazi, T. Inhibition of lamina outgrowth following Solanum lycopersicum auxin response factor 10 (SlARF10) derepression. Plant Mol. Biol. 2012, 78, 561–576. [Google Scholar] [CrossRef] [PubMed]
- Damodharan, S.; Zhao, D.Z.; Arazi, T. A common mi RNA160-based mechanism regulates ovary patterning, floral organ abscission and lamina outgrowth in tomato. Plant J. 2016, 86, 458–471. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lai, T.; Song, H.; Xu, X. MiR164 is involved in delaying senescence of strawberry (Fragaria ananassa) fruit by negatively regulating NAC transcription factor genes under low temperature. Russ. J. Plant Physiol. 2017, 64, 251–259. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, M.; Gan, C.; Ren, Y.; Zhao, X.; Yuan, Z. Riboflavin application delays senescence and relieves decay in harvested strawberries during cold storage by improving antioxidant system. LWT-Food Sci. Technol. 2023, 182, 114810. [Google Scholar] [CrossRef]
- Ru, P.; Xu, L.; Ma, H.; Huang, H. Plant fertility defects induced by the enhanced expression of microRNA167. Cell Res. 2006, 16, 457–465. [Google Scholar] [CrossRef]
- Jung, J.H.; Seo, Y.H.; Seo, P.J.; Reyes, J.L.; Yun, J.; Chua, N.-H.; Park, C.-M. The GIGANTEA-regulated microRNA172 mediates photoperiodic flowering Independent of CONSTANS in Arabidopsis. Plant Cell 2007, 19, 2736–2748. [Google Scholar] [CrossRef]
- Grigorova, B.; Mara, C.; Hollender, C.; Sijacic, P.; Chen, X.; Liu, Z. LEUNIG and SEUSS co-repressors regulate miR172 expression in Arabidopsis flowers. Development 2011, 138, 2451–2456. [Google Scholar] [CrossRef]
- Zhu, Q.-H.; Upadhyaya, N.M.; Gubler, F.; Helliwell, C.A. Over-expression of miR172 causes loss of spikelet determinacy and floral organ abnormalities in rice (Oryza sativa). BMC Plant Biol. 2009, 9, 149. [Google Scholar] [CrossRef]
- Yao, J.; Xu, J.; Cornille, A.; Tomes, S.; Karunairetnam, S.; Luo, Z.; Bassett, H.; Whitworth, C.; Rees-George, J.; Ranatunga, C.; et al. A microRNA allele that emerged prior to apple domestication may underlie fruit size evolution. Plant J. 2015, 84, 417–427. [Google Scholar] [CrossRef]
- Gattolin, S.; Cirilli, M.; Pacheco, I.; Ciacciulli, A.; Linge, C.D.S.; Mauroux, J.; Lambert, P.; Cammarata, E.; Bassi, D.; Pascal, T.; et al. Deletion of the miR172 target site in a TOE-type gene is a strong candidate variant for dominant double-flower trait in Rosaceae. Plant J. 2018, 96, 358–371. [Google Scholar] [CrossRef]
- Sui, M.J.; Yan, H.J.; Wang, Z.Z.; Qiu, X.Q.; Jian, H.Y.; Wang, Q.G.; Chen, M.; Zhang, H.; Tang, K.X. Identification of microRNA associated with flower organ development in Rosa chinensis ‘Viridiflora’. Plant Sci. J. 2019, 37, 37–46. [Google Scholar]
- Varkonyi-Gasic, E.; Lough, R.H.; Moss, S.M.A.; Wu, R.; Hellens, R.P. Kiwifruit floral gene APETALA2 is alternatively spliced and accumulates in aberrant indeterminate flowers in the absence of miR172. Plant Mol. Biol. 2012, 78, 417–429. [Google Scholar] [CrossRef]
- Zhang, X.; Henderson, I.R.; Lu, C.; Green, P.J.; Jacobsen, S.E. Role of RNA polymerase IV in plant small RNA metabolism. Proc. Natl. Acad. Sci. USA 2007, 104, 4536–4541. [Google Scholar] [CrossRef]
- Sieber, P.; Gheyselinck, J.; Gross-Hardt, R.; Laux, T.; Grossniklaus, U.; Schneitz, K. Pattern formation during early ovule development in Arabidopsis thaliana. Dev. Biol. 2004, 273, 321–334. [Google Scholar] [CrossRef]
- Hashimoto, K.; Miyashima, S.; Sato-Nara, K.; Yamada, T.; Nakajima, K. Functionally diversified members of the miR165/6 gene family regulate ovule morphogenesis in Arabidopsis thaliana. Plant Cell Physiol. 2018, 59, 1017–1026. [Google Scholar] [CrossRef]
- Chen, L.N. Identification of microRNAs Involved in Pomegranate Female Sterility and Functional Analysis on the Regulation of Pg-miR166a-3p on Pistil Development. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2020. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | GC Content (%) | Q20/Q30 (%) | Total sRNA | Mapped sRNA |
|---|---|---|---|---|---|---|
| BF1_1 | 17,367,419 | 17,051,495 (98.18%) | 49.96 | 99.04/96.85 | 14,117,927 | 94.72% |
| BF1_2 | 13,704,093 | 13,340,571 (97.35%) | 50.11 | 99.39/97.63 | 8,884,127 | 94.02% |
| BF1_3 | 13,401,810 | 12,885,356 (96.15%) | 50.21 | 99.29/97.67 | 7,375,939 | 95.61% |
| BF2_1 | 21,426,062 | 20,858,435 (97.35%) | 49.25 | 99.31/97.59 | 17,745,249 | 94.55% |
| BF2_2 | 13,406,968 | 12,787,925 (95.38%) | 49.50 | 98.48/95.09 | 10,792,373 | 96.84% |
| BF2_3 | 14,948,932 | 14,681,792 (98.21%) | 49.16 | 99.05/96.87 | 11,500,997 | 94.93% |
| BF3_1 | 21,966,339 | 19,930,652 (90.73%) | 50.56 | 99.15/97.22 | 14,925,094 | 96.23% |
| BF3_2 | 18,298,780 | 15,436,529 (84.36%) | 50.91 | 99.24/97.59 | 5,175,907 | 97.24% |
| BF3_3 | 21,665,923 | 20,317,773 (93.78%) | 50.01 | 98.07/94.05 | 18,571,396 | 95.28% |
| MF1_1 | 21,655,951 | 21,171,252 (97.76%) | 48.37 | 99.41/97.73 | 16,395,117 | 92.75% |
| MF1_2 | 23,257,849 | 22,788,789 (97.98%) | 49.29 | 99.41/97.76 | 19,202,540 | 93.99% |
| MF1_3 | 13,748,317 | 13,525,096 (98.38%) | 49.36 | 99.33/97.61 | 10,193,022 | 94.36% |
| MF2_1 | 14,418,240 | 13,551,334 (93.99%) | 51.05 | 99.43/97.80 | 6,959,440 | 95.06% |
| MF2_2 | 12,606,612 | 12,326,514 (97.78%) | 50.73 | 98.21/94.51 | 6,948,964 | 93.62% |
| MF2_3 | 18,135,526 | 17,633,277 (97.23%) | 49.33 | 99.11/97.08 | 14,981,807 | 96.68% |
| MF3_1 | 21,572,493 | 18,175,437 (84.25%) | 49.52 | 99.13/97.20 | 15,960,976 | 94.39% |
| MF3_2 | 19,010,757 | 17,214,821 (90.55%) | 51.60 | 98.91/96.99 | 10,989,846 | 97.12% |
| MF3_3 | 17,985,939 | 16,567,891 (92.12%) | 53.63 | 99.31/97.74 | 4,197,113 | 96.47% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Huang, J.; Li, M.; Ren, H.; Jiao, J.; Wan, R.; Liu, Y.; Wang, M.; Shi, J.; Zhang, K.; et al. Exploring MicroRNAs Associated with Pomegranate Pistil Development: An Identification and Analysis Study. Horticulturae 2024, 10, 85. https://doi.org/10.3390/horticulturae10010085
Zhao Y, Huang J, Li M, Ren H, Jiao J, Wan R, Liu Y, Wang M, Shi J, Zhang K, et al. Exploring MicroRNAs Associated with Pomegranate Pistil Development: An Identification and Analysis Study. Horticulturae. 2024; 10(1):85. https://doi.org/10.3390/horticulturae10010085
Chicago/Turabian StyleZhao, Yujie, Jingyi Huang, Ming Li, Hongfang Ren, Jian Jiao, Ran Wan, Yu Liu, Miaomiao Wang, Jiangli Shi, Kunxi Zhang, and et al. 2024. "Exploring MicroRNAs Associated with Pomegranate Pistil Development: An Identification and Analysis Study" Horticulturae 10, no. 1: 85. https://doi.org/10.3390/horticulturae10010085
APA StyleZhao, Y., Huang, J., Li, M., Ren, H., Jiao, J., Wan, R., Liu, Y., Wang, M., Shi, J., Zhang, K., Hao, P., Song, S., Bai, T., & Zheng, X. (2024). Exploring MicroRNAs Associated with Pomegranate Pistil Development: An Identification and Analysis Study. Horticulturae, 10(1), 85. https://doi.org/10.3390/horticulturae10010085