Influence of Preharvest Gibberellic Acid Treatments on Postharvest Quality of Minimally Processed Leaf Lettuce and Rocket



Abstract
:1. Introduction
2. Materials and Methods
2.1. Leafy Vegetable Cultivation
2.2. Minimal Processing and Cold Storage
2.3. Statistics and Principal Component Analysis
3. Results
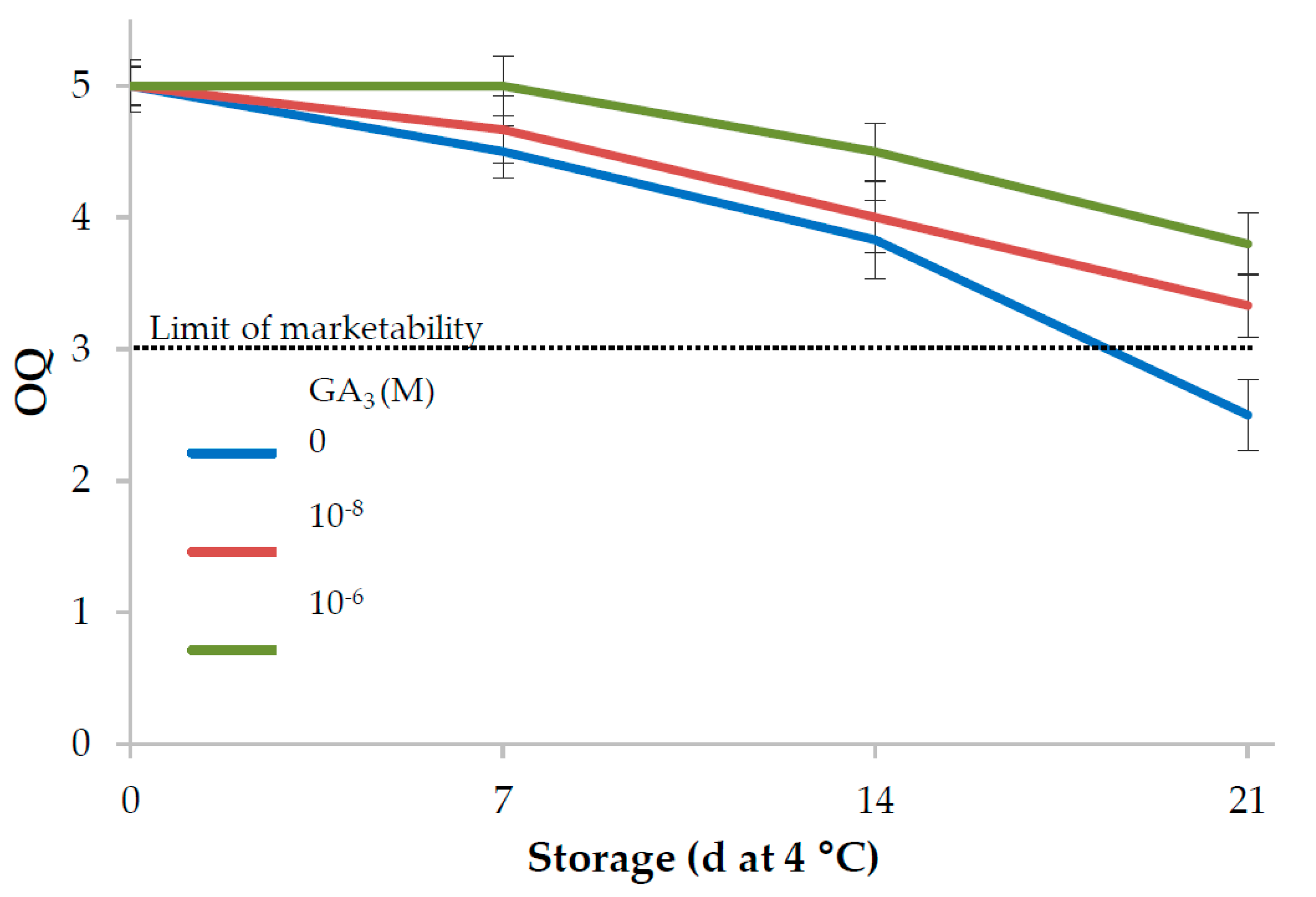
3.1. Leaf Lettuce
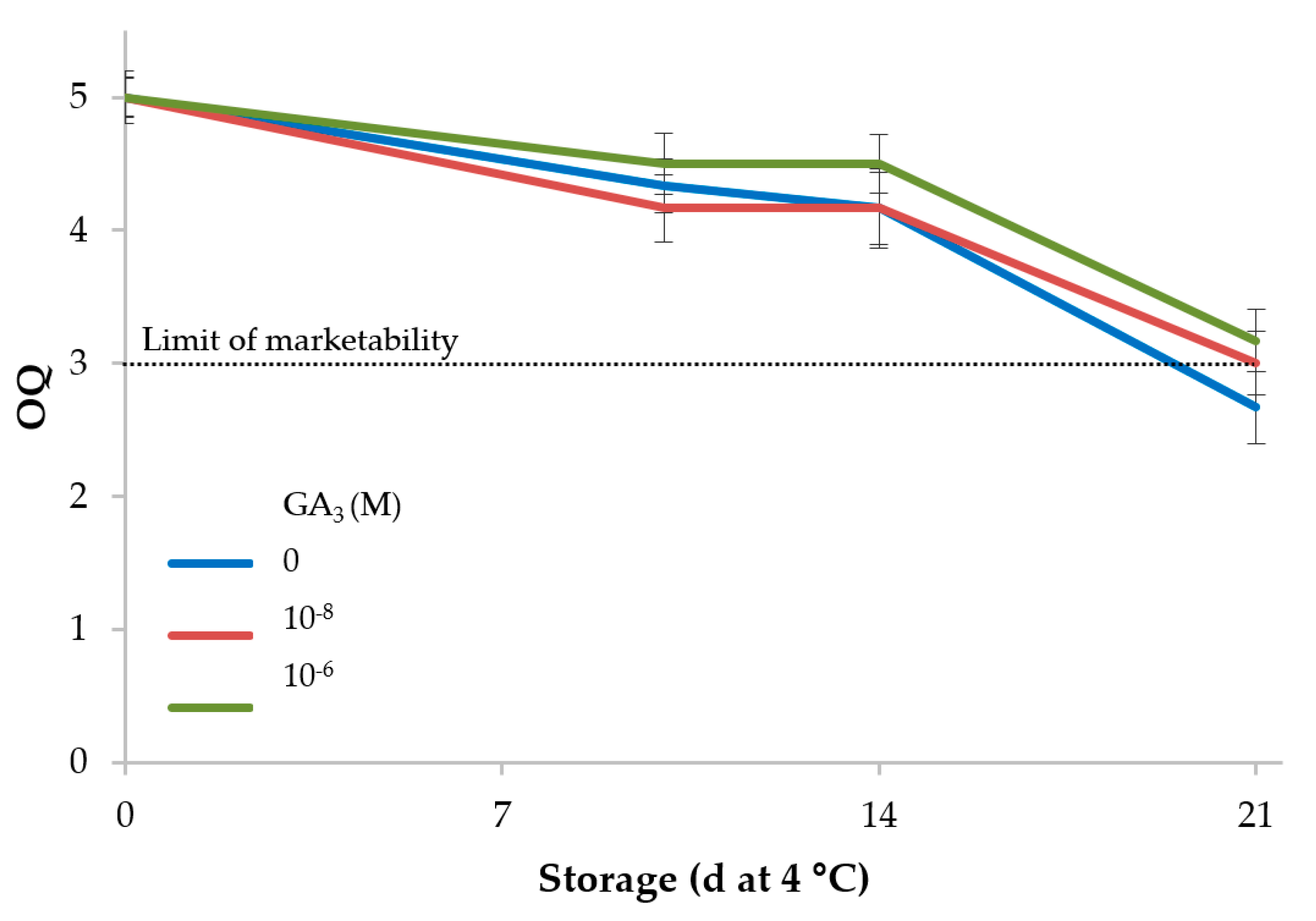
3.2. Rocket
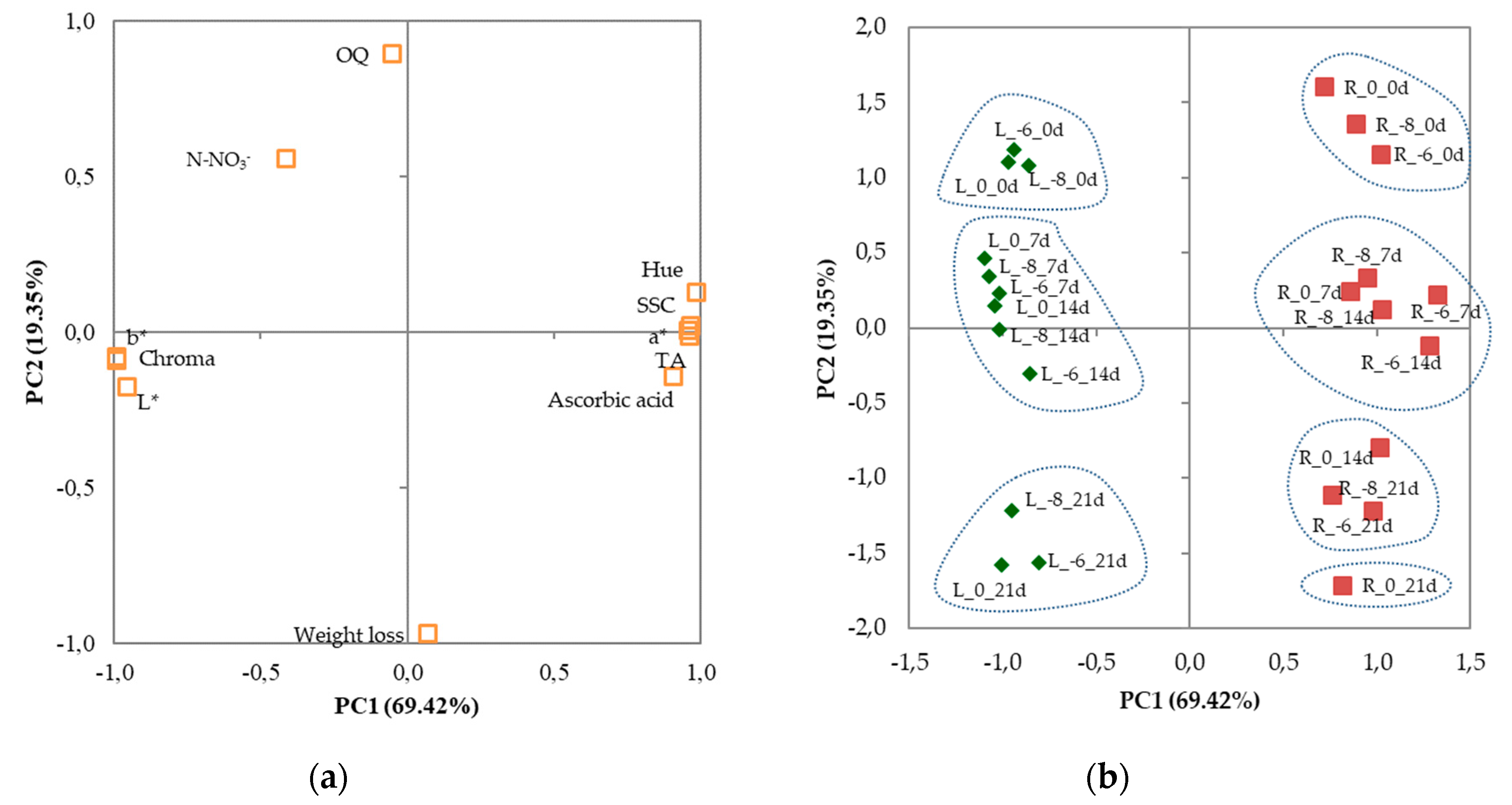
3.3. Principal Components Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chakraborty, I.; Chattopadhyay, A. Pre- and Post-Harvest Losses in Vegetables. In Advances in Postharvest Technologies of Vegetable Crops; CRC Press: Boca Raton, FL, USA, 2018; pp. 25–87. [Google Scholar]
- Lynch, M.F.; Tauxe, R.V.; Hedberg, C.W. The growing burden of foodborne outbreaks due to contaminated fresh produce: Risks and opportunities. Epidemiol. Infect. 2009, 137, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Settanni, L.; Miceli, A.; Francesca, N.; Cruciata, M.; Moschetti, G. Microbiological investigation of Raphanus sativus L. grown hydroponically in nutrient solutions contaminated with spoilage and pathogenic bacteria. Int. J. Food Microbiol. 2013, 160, 344–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomasi, N.; Pinton, R.; Costa, L.D.; Cortella, G.; Terzano, R.; Mimmo, T.; Scampicchio, M.; Cesco, S. New ‘solutions’ for floating cultivation system of ready-to-eat salad: A review. Trends Food Sci. Technol. 2015, 46, 267–276. [Google Scholar] [CrossRef]
- Khan, I.; Tango, C.N.; Miskeen, S.; Lee, B.H.; Oh, D.H. Hurdle technology: A novel approach for enhanced food quality and safety—A review. Food Control 2017, 73, 1426–1444. [Google Scholar] [CrossRef]
- Soerjomataram, I.; Oomen, D.; Lemmens, V.; Oenema, A.; Benetou, V.; Trichopoulou, A.; Coebergh, J.W.; Barendregt, J.; De Vries, E. Increased consumption of fruit and vegetables and future cancer incidence in selected European countries. Eur. J. Cancer 2010, 46, 2563–2580. [Google Scholar] [CrossRef]
- Karam, M.C.; Petit, J.; Zimmer, D.; Djantou, E.B.; Scher, J. Effects of drying and grinding in production of fruit and vegetable powders: A review. J. Food Eng. 2016, 188, 32–49. [Google Scholar] [CrossRef]
- Francesca, N.; Guarcello, R.; Craparo, V.; Moschetti, G.; Settanni, L.; Gaglio, R. Microbial Ecology of Retail Ready-to-Eat Escarole and Red Chicory Sold in Palermo City, Italy. J. Food Qual. Hazards Control 2019, 6, 45–52. [Google Scholar] [CrossRef]
- Miceli, A.; Settanni, L. Influence of agronomic practices and pre-harvest conditions on the attachment and development of Listeria monocytogenes in vegetables. Ann. Microbiol. 2019, 69, 185–199. [Google Scholar] [CrossRef]
- Putnik, P.; Bursać Kovačević, D.; Herceg, K.; Roohinejad, S.; Greiner, R.; Bekhit, A.E.D.A.; Levaj, B. Modelling the shelf-life of minimally-processed fresh-cut apples packaged in a modified atmosphere using food quality parameters. Food Control 2011, 81, 55–64. [Google Scholar] [CrossRef]
- Mir, S.A.; Shah, M.A.; Mir, M.M.; Dar, B.; Greiner, R.; Roohinejad, S. Microbiological contamination of ready-to-eat vegetable salads in developing countries and potential solutions in the supply chain to control microbial pathogens. Food Control 2018, 85, 235–244. [Google Scholar] [CrossRef]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337. [Google Scholar] [CrossRef] [PubMed]
- Kummu, M.; De Moel, H.; Porkka, M.; Siebert, S.; Varis, O.; Ward, P. Lost food, wasted resources: Global food supply chain losses and their impacts on freshwater, cropland, and fertiliser use. Sci. Total Environ. 2012, 438, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.F.; D’Odorico, P.; Rulli, M.C. Moderating diets to feed the future. Earth’s Future 2014, 2, 559–565. [Google Scholar] [CrossRef]
- Davis, K.F.; Gephart, J.A.; Emery, K.A.; Leach, A.M.; Galloway, J.N.; D’Odorico, P. Meeting future food demand with current agricultural resources. Glob. Environ. Chang. 2016, 39, 125–132. [Google Scholar] [CrossRef]
- Mueller, N.D.; Gerber, J.S.; Johnston, M.; Ray, D.K.; Ramankutty, N.; Foley, J.A. Closing yield gaps through nutrient and water management. Nature 2012, 490, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Basra, A. Plant Growth Regulators in Agriculture and Horticulture: Their Role and Commercial Uses; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Shah, S.H.; Ahmad, I. Samiullah Effect of gibberellic acid spray on growth.; nutrient uptake and yield attributes during various growth stages of black cumin (Nigella sativa L.). Asian J. Plant Sci. 2006, 5, 881–884. [Google Scholar]
- Hedden, P.; Sponsel, V. A Century of Gibberellin Research. J. Plant Growth Regul. 2015, 34, 740–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azuma, T.; Ueno, S.; Uchida, N.; Yasuda, T. Gibberellin-induced elongation and osmoregulation in internodes of floating rice. Physiol. Plant. 1997, 99, 517–522. [Google Scholar] [CrossRef]
- Bhaskar, S.; Vasantha Kumar, T.; Srivastava, H.C. Influence of growth regulators on production of herbage and oil in patchouli (Pogostemon patchouli). Ind. Perfum. 1997, 41, 98–101. [Google Scholar]
- Brock, T.G. Combined effects of hormones and light during growth promotion in primary leaves of Phaseolus vulgaris. Can. J. Bot. 1993, 71, 501–505. [Google Scholar] [CrossRef]
- Emongor, V.E. Effect of benzyladenine and gibberellins on growth, yield and yield components of common bean (Phaseolus vulgaris). UNISWA. Res. J. Agric. Sci. Technol. 2002, 6, 65–72. [Google Scholar] [CrossRef]
- Gupta, V.N.; Datta, S.K. Influence of gibberellic acid on growth and flowering in chrysanthemum (Chrysanthemum morifolium Ramat) cv. Jayanti. Ind. J. Plant Physiol. 2001, 6, 420–422. [Google Scholar]
- Khan, N.A. Samiullah Comparative effect of modes of gibberellic acid application on photosynthetic biomass distribution and productivity of rapeseed-mustard. Physiol. Mol. Biol. Plants 2003, 9, 141–145. [Google Scholar]
- Richards, D.E.; King, K.E.; Ait-Ali, T.; Harberd, N.P. How gibberellin regulates plant growth and development: A molecular genetic analysis of gibberellin signaling. Annu. Rev. Plant Biol. 2001, 52, 67–88. [Google Scholar] [CrossRef] [PubMed]
- Takei, K.; Takahashi, T.; Sugiyama, T.; Yamaya, T.; Sakakibara, H. Multiple routes communicating nitrogen availability from roots to shoots: A signal transduction pathway mediated by cytokinin. J. Exp. Bot. 2002, 53, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Wareing, P.F.; Phillips, I.D.J. Growth and Differentiation in Plants, 3rd ed.; Pergamon Press: Oxford, UK, 1981. [Google Scholar]
- Yang, Y.Z.; Lin, D.C.; Guo, Z.Y. Promotion of fruit development in cucumber (Cucumis sativus) by thidiazuron. Sci. Hortic. 1992, 50, 47–51. [Google Scholar] [CrossRef]
- Aharoni, N.; Dvir, O.; Chalupowicz, D.; Aharon, Z. Coping with postharvest physiology of fresh culinary herbs. Acta Hortic. 1993, 344, 69–78. [Google Scholar] [CrossRef]
- Ding, Y.; Sheng, J.; Li, S.; Nie, Y.; Zhao, J.; Zhu, Z.; Wang, Z.; Tang, X. The role of gibberellins in the mitigation of chilling injury in cherry tomato (Solanum lycopersicum L.) fruit. Postharvest Biol. Technol. 2015, 101, 88–95. [Google Scholar] [CrossRef]
- Valero, D.; Serrano, M.; Burló, F.; Carbonell, A.; Burgos, L.; Riquelme, F.; Martinez-Romero, D.; Martínez-Romero, D. Exogenous Polyamines and Gibberellic Acid Effects on Peach (Prunus persica L.) Storability Improvement. J. Food Sci. 2000, 65, 288–294. [Google Scholar]
- Yu, T.; Wu, P.G.; Qi, J.J.; Zheng, X.D.; Jiang, F.; Zhan, X. Improved control of postharvest blue mold rot in pear fruit by a combination of Cryptococcus laurentii and gibberellic acid. Biol. Control 2006, 39, 128–134. [Google Scholar] [CrossRef]
- Robert, E.C. Introduction: Nature, Occurrence and Functioning of Plant Hormones. In Biochemistry and Molecular Biology of Plant Hormones; Hooykaas, P.J.J., Hall, M.A., Libbenga, K.R., Eds.; Elsevier: Amsterdam, The Netherlands; New York, NY, USA; London, UK, 1999; pp. 3–22. [Google Scholar]
- Janick, J. Horticultural Science; Freeman & Co.: San Francisco, CA, USA, 1979. [Google Scholar]
- Khan, M.M.A.; Gautam, C.; Mohammad, F.; Siddiqui, M.H.; Naeem, M.; Khan, M.N. Effect of gibberellic acid spray on performance of tomato. Turk. J. Biol. 2006, 30, 11–16. [Google Scholar]
- Khan, N.A.; Ansari, H.R. Samiullah Effect of gibberellic acid spray during ontogeny of mustard on growth, nutrient uptake and yield characteristics. J. Agron. Crop Sci. 1998, 181, 61–73. [Google Scholar] [CrossRef]
- King, R.W.; Evans, L.T. Gibberellins and flowering of grasses and cereals: Prizing open the lid of the “florigen black box”. Annu. Rev. Plant Biol. 2003, 54, 307–328. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, R.K.; Prabhakar, B.S. Effect of gibberellin on rice yield response to nitrogen applied at heading, and quality of seeds. Plant Soil 1980, 55, 153–156. [Google Scholar] [CrossRef]
- Pal, P.; Yadav, K.; Kumar, K.; Singh, N. Effect of Gibberellic Acid and Potassium Foliar Sprays on Productivity and Physiological and Biochemical Parameters of Parthenocarpic Cucumber cv. ‘Seven Star F1’. J. Hortic. Res. 2016, 24, 93–100. [Google Scholar] [CrossRef]
- Singh, S.P. Response of tomatoes to growth substances—A review. Adv. Hort. Forest. 1995, 4, 73–84. [Google Scholar]
- Miceli, A.; Moncada, A.; Sabatino, L.; Vetrano, F. Effect of Gibberellic Acid on Growth, Yield, and Quality of Leaf Lettuce and Rocket Grown in a Floating System. Agronomy 2019, 9, 382. [Google Scholar] [CrossRef]
- Sonneveld, C.; Voogt, W. Plant Nutrition of Greenhouse Crops; Springer Science and Business Media LLC: Berlin, Germany, 2009. [Google Scholar]
- Goto, E.; Both, A.; Albright, L.; Langhans, R.; Leed, A. Effect of dissolved oxygen concentration on lettuce growth in floating hydroponics. Acta Hortic. 1996, 440, 205–210. [Google Scholar] [CrossRef]
- McGuire, R.G. Reporting of Objective Color Measurements. HortScience 1992, 27, 1254–1255. [Google Scholar] [CrossRef] [Green Version]
- Merck Reagenti, Prodotti Chimici e Attrezzature per il Laboratorio. Available online: http://www.merckmillipore.com/chemicals (accessed on 28 May 2019).
- Hooley, R. Gibberellins: Perception, transduction and responses. Plant Mol. Biol. 1994, 26, 1529–1555. [Google Scholar] [CrossRef]
- Da Silva Vieira, M.R.; Citadini, V.; Lima, G.P.P.; de Souza, A.V.; de Souza Alves, L. Use of gibberellin in floriculture. Afr. J. Biotechnol. 2010, 9, 9118–9121. [Google Scholar]
- Khan, N.A.; Mir, R.; Khan, M.; Javid, S.; Samiullah. Effects of gibberellic acid spray on nitrogen yield efficiency of mustard grown with different nitrogen levels. Plant Growth Regul. 2002, 38, 243–247. [Google Scholar] [CrossRef]
- Shah, S.H. Effects of salt stress on mustard as affected by gibberellic acid application. Gen. Appl. Plant Physiol. 2007, 33, 97–106. [Google Scholar]
- Gelmesa, D.; Abebie, B.; Desalegn, L. Effects of Gibberellic acid and 2, 4-dichlorophenoxyacetic acid spray on fruit yield and quality of tomato (Lycopersicon esculentum Mill.). J. Plant Breed. Crop Sci. 2010, 2, 316–324. [Google Scholar]
- Ludford, P.M. Postharvest Hormone Changes in Vegetables and Fruit. In Plant Hormones; Davies, P.J., Ed.; Kluwer Academic Publishers: Dordrecht, NL, USA, 1995. [Google Scholar]
- Tang, X.M.; Liu, C.; Zhong, R.C.; Jiang, J.; Han, Z.Q.; He, L.Q.; Li, Z.; Xiong, F.Q.; Tang, R.H. Comparative studies on the chemical regulation of peanuts by paclobutrazol, mepiquat chloride and chlorocholine chloride. J. South. Agric. 2011, 42, 603–605. [Google Scholar]
- Xie, B.T.; Wang, Q.M.; Zhang, H.Y.; Li, A.X.; Hou, F.Y.; Wang, B.Q.; Dong, S.X.; Zhang, L.M. The effect of plant growth regulators on the yield and phytohormone concentration in sweet potato. Acta Agric. Boreali Sin. 2016, 31, 155–161. [Google Scholar]
- Zhang, Y. Research status of plant growth regulators on crop regulation. Mod. Agric. 2016, 5, 31–34. [Google Scholar]
- Han, H.Y.; Wang, F.A.; Chen, B.; Li, B.A.; Zhang, W.F.; Tian, X.L.; Li, Z. Effect of nitrogen fertilizer on plant growth and yield formation of cotton applied with fortified DPC. J. China Agric. Univ. 2017, 22, 12–20. [Google Scholar]
- Zhang, M.; Liu, M.; Zhang, Y.; Ji, Y.; Zhao, M.; Wu, Z. Effect of different plant growth regulator added in nutrient solution on growth and development of summer tomato seedling. North Hortic. 2017, 6, 8–13. [Google Scholar]
- Khan, N.A. Effect of gibberellic acid on carbonic anhydrase, photosynthesis, growth and yield of mustard. Biol. Plant. 1996, 38, 145–147. [Google Scholar] [CrossRef]
- Sültemeyer, D.; Schmidt, C.; Fock, H.P. Carbonic anhydrases in higher plants and aquatic microorganisms. Physiol. Plant. 1993, 88, 179–190. [Google Scholar] [CrossRef]
- Khan, N.A. Variation in carbonic anhydrase activity and its relationship with photosynthesis and dry mass of mustard. Photosynthetica 1994, 30, 317–320. [Google Scholar]
- Roura, S.I.; Davidovich, L.A.; Del Valle, C.E. Postharvest changes in fresh Swiss chard (Beta vulgaris. type cycla) under different storage conditions. J. Food Qual. 2000, 23, 137–147. [Google Scholar] [CrossRef]
- Roura, S.I.; Davidovich, L.A.; Del Valle, C.E. Quality loss in minimally processed Swiss chard related to amount of damaged area. Lebensm. Wiss. Technol. 2000, 33, 53–59. [Google Scholar] [CrossRef]
- Hodges, D.M.; Toivonen, P.M. Quality of fresh-cut fruits and vegetables as affected by exposure to abiotic stress. Postharvest Biol. Technol. 2008, 48, 155–162. [Google Scholar] [CrossRef]
- Toivonen, P.M.A.; DeEll, J.R. Physiology of fresh-cut fruits and vegetables. In Physiology of Fresh-Cut Fruits and Vegetables: Science, Technology, and Market; Lamikanra, O., Ed.; CRC Press: Boca Raton, FL, USA, 2002; pp. 91–123. [Google Scholar]
- Alfonzo, A.; Gaglio, R.; Miceli, A.; Francesca, N.; Di Gerlando, R.; Moschetti, G.; Settanni, L. Shelf life evaluation of fresh-cut red chicory subjected to different minimal processes. Food Microbiol. 2018, 73, 298–304. [Google Scholar] [CrossRef]
- Miceli, A.; Gaglio, R.; Francesca, N.; Ciminata, A.; Moschetti, G.; Settanni, L. Evolution of shelf life parameters of ready-to-eat escarole (Cichorium endivia var. latifolium) subjected to different cutting operations. Sci. Hortic. 2019, 247, 175–183. [Google Scholar] [CrossRef]
- Miceli, A.; Miceli, C. Effect of Nitrogen Fertilization on the Quality of Swiss Chard at Harvest and during Storage as Minimally Processed Produce. J. Food Qual. 2014, 37, 125–134. [Google Scholar] [CrossRef]
- Miceli, A.; Romano, C.; Moncada, A.; D’Anna, F.; Vetrano, F. Effect of cold storage on the quality of minimally processed cauliflower. Carpath. J. Food Sci. Technol. 2015, 7, 70–74. [Google Scholar]
- Watada, A.E.; Qi, L. Quality of fresh-cut produce. Postharv. Biol. Technol. 1999, 15, 201–205. [Google Scholar] [CrossRef]
- Demirsoy, L.; Bilgener, S. The effect of chemical applications on cuticular and epidermal properties of some sweet cherry cultivars with respect to fruit cracking susceptibility. Turk. J. Agric. For. 2000, 24, 541–550. [Google Scholar]
- Sams, C.E. Preharvest factors affecting postharvest texture. Postharvest Biol. Technol. 1999, 15, 249–254. [Google Scholar] [CrossRef]
- Gang, C.; Li, J.; Chen, Y.; Wang, Y.; Li, H.; Pan, B.; Odeh, I. Synergistic effect of chemical treatments on storage quality and chilling injury of honey peaches. J. Food Process. Preserv. 2015, 39, 1108–1117. [Google Scholar] [CrossRef]
- Miceli, A.; Vetrano, F.; Romano, C. Effect of hot air treatment on minimally processed cauliflower. Acta Hortic. 2013, 1005, 309–314. [Google Scholar] [CrossRef]
- Lers, A.; Jiang, W.; Lomaniec, E.; Aharoni, N. Gibberellic Acid and CO2 Additive Effect in Retarding Postharvest Senescence of Parsley. J. Food Sci. 1998, 63, 66–68. [Google Scholar] [CrossRef]
- Gaglio, R.; Miceli, A.; Sardina, M.T.; Francesca, N.; Moschetti, G.; Settanni, L. Evaluation of microbiological and physico-chemical parameters of retail ready-to-eat mono-varietal salads. J. Food Process. Preserv. 2019, 43, e13955. [Google Scholar] [CrossRef]
- Jacob, R.A.; Sotoudeh, G. Vitamin C Function and Status in Chronic Disease. Nutr. Clin. Care 2002, 5, 66–74. [Google Scholar] [CrossRef]
- Eheart, M.S.; Odland, D. Storage of fresh broccoli and green beans. J. Am. Diet. Assoc. 1972, 60, 402–406. [Google Scholar]
- Esteve, M.J.; Farré, R.; Frígola, A.; Clemente, G. Changes in Ascorbic Acid Content of Green Asparagus during the Harvesting Period and Storage. J. Agric. Food Chem. 1995, 43, 2058–2061. [Google Scholar] [CrossRef]
- Howard, L.; Wong, A.; Perry, A.; Klein, B. β-Carotene and Ascorbic Acid Retention in Fresh and Processed Vegetables. J. Food Sci. 1999, 64, 929–936. [Google Scholar] [CrossRef]
- Wu, Y.; Perry, A.K.; Klein, B.P. Vitamin C and B-carotene in fresh and frozen green beans and broccoli in a simulated system. J. Food Qual. 1992, 15, 87–96. [Google Scholar] [CrossRef]
- El-Hamahmy, M.A.; Elsayed, A.I.; Odero, D.C. Physiological effects of hot water dipping, chitosan coating and gibberellic acid on shelf-life and quality assurance of sugar snap peas (Pisum sativum L. var. macrocarpon). Food Packag. Shelf Life 2017, 11, 58–66. [Google Scholar] [CrossRef]
- Lee, S.K.; Kader, A.A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Stefanelli, D.; Goodwin, I.; Jones, R. Minimal nitrogen and water use in horticulture: Effects on quality and content of selected nutrients. Food Res. Int. 2010, 43, 1833–1843. [Google Scholar] [CrossRef]
- Mozafar, A. Plant Vitamins. Agronomic, Physiological and Nutritional Aspects; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- European Union Commission Regulation (EC) No. 1258/2011 of 2 December 2011 amending Regulation (EC) No. 1881/2006 as regards maximum levels for nitrates in foodstuffs. Off. J. Eur. Union 2011, L320, 15–17.
- Chanda, S.V.; Sood, C.R.; Reddy, V.S.; Singh, Y.D. Influence of plant growth regulators on some enzymes of nitrogen assimilation in mustard seedlings. J. Plant Nutr. 1998, 21, 1765–1777. [Google Scholar] [CrossRef]
- Moncada, A.; Miceli, A.; Sabatino, L.; Iapichino, G.; D’Anna, F.; Vetrano, F. Effect of Molybdenum Rate on Yield and Quality of Lettuce, Escarole, and Curly Endive Grown in a Floating System. Agronomy 2018, 8, 171. [Google Scholar] [CrossRef]
- La Scalia, G.; Aiello, G.; Miceli, A.; Nasca, A.; Alfonzo, A.; Settanni, L. Effect of vibration on the quality of strawberry fruits caused by simulated transport. J. Food Process Eng. 2016, 39, 140–156. [Google Scholar] [CrossRef]
- Ihl, M.; Shene, C.; Scheuermann, E.; Bifani, V. Correlation for pigment content through color determination using tristimulus values in a green leafy vegetable, Swiss chard. J. Sci. Food Agric. 1994, 66, 527–531. [Google Scholar] [CrossRef]
- Madeira, A.C.; Ferreira, A.; De Varennes, A.; Vieira, M.I. SPAD Meter Versus Tristimulus Colorimeter to Estimate Chlorophyll Content and Leaf Color in Sweet Pepper. Commun. Soil Sci. Plant Anal. 2003, 34, 2461–2470. [Google Scholar] [CrossRef]
- Aharoni, N.; Back, A.; Ben-Yehushua, S.; Richmond, A.E. Exogenous gibberellic acid and the cytokinin isopentenyladenine retardants of senescence in romaine lettuce. J. Am. Soc. Hortic. Sci. 1975, 100, 4–6. [Google Scholar]
- Aharoni, N. Interrelationship between ethylene and growth regulators in the senescence of lettuce leaf discs. J. Plant Growth Regul. 1989, 8, 309–317. [Google Scholar] [CrossRef]
- Misra, A.N.; Biswal, U.C. Effect of phytohormones on chlorophyll degradation during aging of chloroplasts in vivo and in vitro. Protoplasma 1980, 105, 1–8. [Google Scholar] [CrossRef]
- Chin, T.Y.; Beevers, L. Changes in endogenous growth regulators in Nasturtium leaves during senescence. Planta 1970, 92, 178–188. [Google Scholar] [CrossRef]
- Li, J.R.; Yu, K.; Wei, J.R.; Ma, Q.; Wang, B.Q.; Yu, D. Gibberellin retards chlorophyll degradation during senescence of Paris polyphylla. Biol. Plant. 2010, 54, 395–399. [Google Scholar] [CrossRef]
- Van Doorn, W.G.; Van Lieburg, M.J. Interaction between the effects of phytochrome and gibberellic acid on the senescence of Alstroemeria pelegrina leaves. Physiol. Plant. 1993, 89, 182–186. [Google Scholar] [CrossRef]
- Cefola, M.; Carbone, V.; Minasi, P.; Pace, B. Phenolic profiles and postharvest quality changes of fresh-cut radicchio (Cichorium intybus L.): Nutrient value in fresh vs. stored leaves. J. Food Compos. Anal. 2016, 51, 76–84. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Yield | GA3 (M) | ||
|---|---|---|---|
| 0 | 10−8 | 10−6 | |
| Leaf Lettuce | |||
| Crop yield (kg m−2) | 4.7b z | 5.2b | 7.0a |
| Minimal processing yield (%) | 84.6a | 86.6a | 85.0a |
| Minimal processing yield (kg m−2) | 4.0b | 4.5b | 5.9a |
| Rocket | |||
| Crop yield (kg m−2) | 2.9b | 3.3ab | 4.1a |
| Minimal processing yield (%) | 70.6a | 71.3a | 71.4a |
| Minimal processing yield (kg m−2) | 2.0b | 2.3ab | 2.9a |
| Treatments | Weight Loss (g 100 g−1 FW) | TSS (°Brix) | TAy (mg 100 g−1 FW) | Ascorbic Acid (mg 100 g−1 FW) | N-NO3− (mg kg−1 FW) | |
|---|---|---|---|---|---|---|
| GA3 (M) | ||||||
| 0 | 0.83 z | 3.1 | 29.7 | 59.6 | 2599.7a | |
| 10−8 | 0.84 | 3.1 | 27.9 | 57.4 | 2337.2a | |
| 10−6 | 0.93 | 3.1 | 29.5 | 56.9 | 1848.8b | |
| Storage (d at 4 °C) | ||||||
| 0 | 2.8b | 28.5 | 63.2 | 2374.8 | ||
| 7 | 0.50b | 3.2ab | 27.8 | 49.8 | 2265.6 | |
| 14 | 0.70b | 3.0ab | 31.1 | 59.8 | 2410.0 | |
| 21 | 1.41a | 3.3a | 28.8 | 59.0 | 1997.2 | |
| GA3 × Storage | ||||||
| 0 | 0 | 2.9 | 30.7 | 78.5 | 2690.3 | |
| 0 | 10 | 0.40 | 3.3 | 28.5 | 52.5 | 2495.0 |
| 0 | 14 | 0.52 | 2.9 | 29.8 | 52.5 | 2665.0 |
| 0 | 21 | 1.59 | 3.2 | 29.8 | 55.0 | 2548.3 |
| 10−8 | 0 | 3.1 | 28.8 | 53.0 | 2328.8 | |
| 10−8 | 10 | 0.57 | 3.2 | 26.9 | 49.0 | 2376.7 |
| 10−8 | 14 | 0.77 | 2.8 | 28.8 | 65.0 | 2640.0 |
| 10−8 | 21 | 1.19 | 3.2 | 26.9 | 62.5 | 2003.3 |
| 10−6 | 0 | 2.5 | 25.9 | 58.0 | 2105.3 | |
| 10−6 | 10 | 0.53 | 3.1 | 27.9 | 48.0 | 1925.0 |
| 10−6 | 14 | 0.82 | 3.3 | 34.6 | 62.0 | 1925.0 |
| 10−6 | 21 | 1.45 | 3.4 | 29.8 | 59.5 | 1440.0 |
| GA3 | ns x | ns | ns | ns | ** | |
| Storage | *** | * | ns | ns | ns | |
| GA3 × Storage | ns | ns | ns | ns | ns | |
| Treatments | L* | a* | b* | Chroma | Hue | OQ | |
|---|---|---|---|---|---|---|---|
| GA3 (M) | |||||||
| 0 | 56.4 z | −21.7 | 39.4a | 44.4 | 118.9 | 4.0b | |
| 10−8 | 55.6 | −21.5 | 38.9ab | 44.4 | 119.0 | 4.3ab | |
| 10−6 | 55.3 | −21.7 | 38.7b | 45.0 | 119.2 | 4.6a | |
| Storage (d at 4 °C) | |||||||
| 0 | 52.2 | −21.8ab | 38.2c | 44.0b | 119.8a | 5.0a | |
| 7 | 57.0 | −22.0a | 39.4bc | 45.2a | 119.2b | 4.7a | |
| 14 | 57.0 | −21.5bc | 38.8ab | 44.3ab | 119.0b | 4.1b | |
| 21 | 56.7 | −21.3c | 39.6a | 45.0a | 118.2c | 3.2c | |
| GA3 × Storage | |||||||
| 0 | 0 | 54.8a | −21.7 | 39.0 | 44.1 | 119.1 | 5.0 |
| 0 | 10 | 57.7b | −22.1 | 39.6 | 45.1 | 119.1 | 4.5 |
| 0 | 14 | 56.7ab | −21.6 | 39.0 | 43.7 | 119.1 | 3.8 |
| 0 | 21 | 56.3ab | −21.4 | 40.0 | 44.8 | 118.2 | 2.5 |
| 10−8 | 0 | 51.0c | −22.0 | 38.2 | 43.1 | 120.0 | 5.0 |
| 10−8 | 10 | 56.7ab | −21.8 | 39.4 | 45.1 | 119.0 | 4.7 |
| 10−8 | 14 | 57.1ab | −21.2 | 38.2 | 44.7 | 119.0 | 4.0 |
| 10−8 | 21 | 57.6a | −21.1 | 39.6 | 44.8 | 118.1 | 3.3 |
| 10−6 | 0 | 50.9c | −21.7 | 37.2 | 44.7 | 120.3 | 5.0 |
| 10−6 | 10 | 56.7ab | −22.1 | 39.3 | 45.4 | 119.3 | 5.0 |
| 10−6 | 14 | 57.3ab | −21.5 | 39.1 | 44.6 | 118.9 | 4.5 |
| 10−6 | 21 | 56.2ab | −21.3 | 39.4 | 45.4 | 118.4 | 3.8 |
| GA3 | * x | ns | * | ns | ns | *** | |
| Storage | *** | *** | *** | *** | *** | *** | |
| GA3 × Storage | ** | ns | ns | ns | ns | ns | |
| Treatments | Weight Loss (g 100 g−1 FW) | TSS (°Brix) | TA y (mg 100 g−1 FW) | Ascorbic Acid (mg 100 g−1 FW) | N-NO3− (mg kg−1 FW) | |
|---|---|---|---|---|---|---|
| GA3 (M) | ||||||
| 0 | 1.29a z | 5.6 | 48.8 | 220.5b | 2171.1a | |
| 10−8 | 0.82b | 5.5 | 47.7 | 209.7b | 2156.1a | |
| 10−6 | 0.79b | 5.8 | 48.7 | 257.5a | 1777.4b | |
| Storage (d at 4 °C) | ||||||
| 0 | 5.5bc | 45.3b | 204.0b | 2383.2a | ||
| 7 | 0.63b | 6.1a | 51.7a | 158.3b | 2044.9ab | |
| 14 | 0.98ab | 5.7ab | 53.4a | 300.0a | 1919.9ab | |
| 21 | 1.29a | 5.1c | 43.3b | 254.7a | 1791.6b | |
| GA3 × Storage | ||||||
| 0 | 0 | 5.8 | 46.1 | 173.0 | 2714.9 | |
| 0 | 10 | 0.66 | 6.2 | 48.7 | 144.0 | 2079.9 |
| 0 | 14 | 1.54 | 5.3 | 55.1 | 296.0 | 1939.9 |
| 0 | 21 | 1.66 | 5.1 | 45.5 | 269.0 | 1949.9 |
| 10−8 | 0 | 5.2 | 44.8 | 221.0 | 2364.9 | |
| 10−8 | 10 | 0.63 | 6.0 | 51.2 | 128.0 | 2224.9 |
| 10−8 | 14 | 0.71 | 5.4 | 53.8 | 282.0 | 2199.9 |
| 10−8 | 21 | 1.11 | 5.3 | 41.0 | 208.0 | 1834.9 |
| 10−6 | 0 | 5.4 | 44.8 | 218.0 | 2069.9 | |
| 10−6 | 10 | 0.60 | 6.2 | 55.1 | 203.0 | 1829.9 |
| 10−6 | 14 | 0.68 | 6.5 | 51.2 | 322.0 | 1619.9 |
| 10−6 | 21 | 1.09 | 5.0 | 43.5 | 287.0 | 1589.9 |
| GA3 | ** x | ns | ns | ** | * | |
| Storage | *** | *** | *** | *** | * | |
| GA3 × Storage | ns | ns | ns | ns | ns | |
| Treatments | L* | a* | b* | Chroma | Hue | OQ | |
|---|---|---|---|---|---|---|---|
| GA3 (M) | |||||||
| 0 | 45.2a z | −19.5b | 26.5a | 32.9a | 126.6 | 4.0 | |
| 10−8 | 44.4b | −19.1b | 25.7ab | 32.0ab | 126.9 | 4.1 | |
| 10−6 | 44.4b | −18.6a | 24.8b | 31.0b | 127.1 | 4.3 | |
| Storage (d at 4 °C) | |||||||
| 0 | 43.7c | −19.0ab | 25.2b | 31.6b | 127.2a | 5.0a | |
| 7 | 44.5bc | −18.7a | 24.9b | 31.2b | 127.1a | 4.3b | |
| 14 | 45.0ab | −19.4b | 25.8ab | 32.3ab | 127.2a | 4.3b | |
| 21 | 45.5a | −19.2b | 26.7a | 33.0a | 126.0b | 2.9c | |
| GA3 × Storage | |||||||
| 0 | 0 | 44.7 | −19.4 | 26.2 | 32.6 | 126.8 | 5.0 |
| 0 | 10 | 45.5 | −19.3 | 26.1 | 32.4 | 126.6 | 4.3 |
| 0 | 14 | 45.2 | −19.9 | 26.5 | 33.1 | 127.1 | 4.2 |
| 0 | 21 | 45.5 | −19.6 | 27.2 | 33.6 | 125.9 | 2.7 |
| 10−8 | 0 | 43.2 | −19.0 | 25.1 | 31.5 | 127.4 | 5.0 |
| 10−8 | 10 | 44.0 | −18.9 | 25.1 | 31.4 | 127.1 | 4.2 |
| 10−8 | 14 | 44.8 | −19.5 | 25.6 | 32.2 | 127.4 | 4.2 |
| 10−8 | 21 | 45.7 | −19.3 | 26.9 | 33.1 | 125.9 | 3.0 |
| 10−6 | 0 | 43.1 | −18.6 | 24.3 | 30.6 | 127.5 | 5.0 |
| 10−6 | 10 | 44.0 | −18.0 | 23.5 | 29.6 | 127.6 | 4.5 |
| 10−6 | 14 | 45.0 | −19.0 | 25.3 | 31.6 | 127.1 | 4.5 |
| 10−6 | 21 | 45.3 | −18.8 | 26.1 | 32.2 | 126.1 | 3.2 |
| GA3 | * x | *** | *** | *** | ns | ns | |
| Storage | *** | *** | ** | ** | *** | *** | |
| GA3 × Storage | ns | ns | ns | ns | ns | ns | |
| Variables | PC1 | PC2 |
|---|---|---|
| Weight loss | 0.071 | −0.968 |
| TSS | 0.965 | 0.021 |
| TA | 0.958 | 0.006 |
| Ascorbic acid | 0.910 | −0.144 |
| N-NO3- | −0.413 | 0.557 |
| L* | −0.955 | −0.174 |
| a* | 0.964 | −0.011 |
| b* | −0.992 | −0.090 |
| Chroma | −0.991 | −0.080 |
| Hue | 0.986 | 0.129 |
| OQ | −0.053 | 0.894 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miceli, A.; Vetrano, F.; Sabatino, L.; D’Anna, F.; Moncada, A. Influence of Preharvest Gibberellic Acid Treatments on Postharvest Quality of Minimally Processed Leaf Lettuce and Rocket. Horticulturae 2019, 5, 63. https://doi.org/10.3390/horticulturae5030063
Miceli A, Vetrano F, Sabatino L, D’Anna F, Moncada A. Influence of Preharvest Gibberellic Acid Treatments on Postharvest Quality of Minimally Processed Leaf Lettuce and Rocket. Horticulturae. 2019; 5(3):63. https://doi.org/10.3390/horticulturae5030063
Chicago/Turabian StyleMiceli, Alessandro, Filippo Vetrano, Leo Sabatino, Fabio D’Anna, and Alessandra Moncada. 2019. "Influence of Preharvest Gibberellic Acid Treatments on Postharvest Quality of Minimally Processed Leaf Lettuce and Rocket" Horticulturae 5, no. 3: 63. https://doi.org/10.3390/horticulturae5030063
APA StyleMiceli, A., Vetrano, F., Sabatino, L., D’Anna, F., & Moncada, A. (2019). Influence of Preharvest Gibberellic Acid Treatments on Postharvest Quality of Minimally Processed Leaf Lettuce and Rocket. Horticulturae, 5(3), 63. https://doi.org/10.3390/horticulturae5030063