Improvement of Phenylpropanoid Production with Elicitor Treatments in Pimpinella brachycarpa Nakai


Abstract
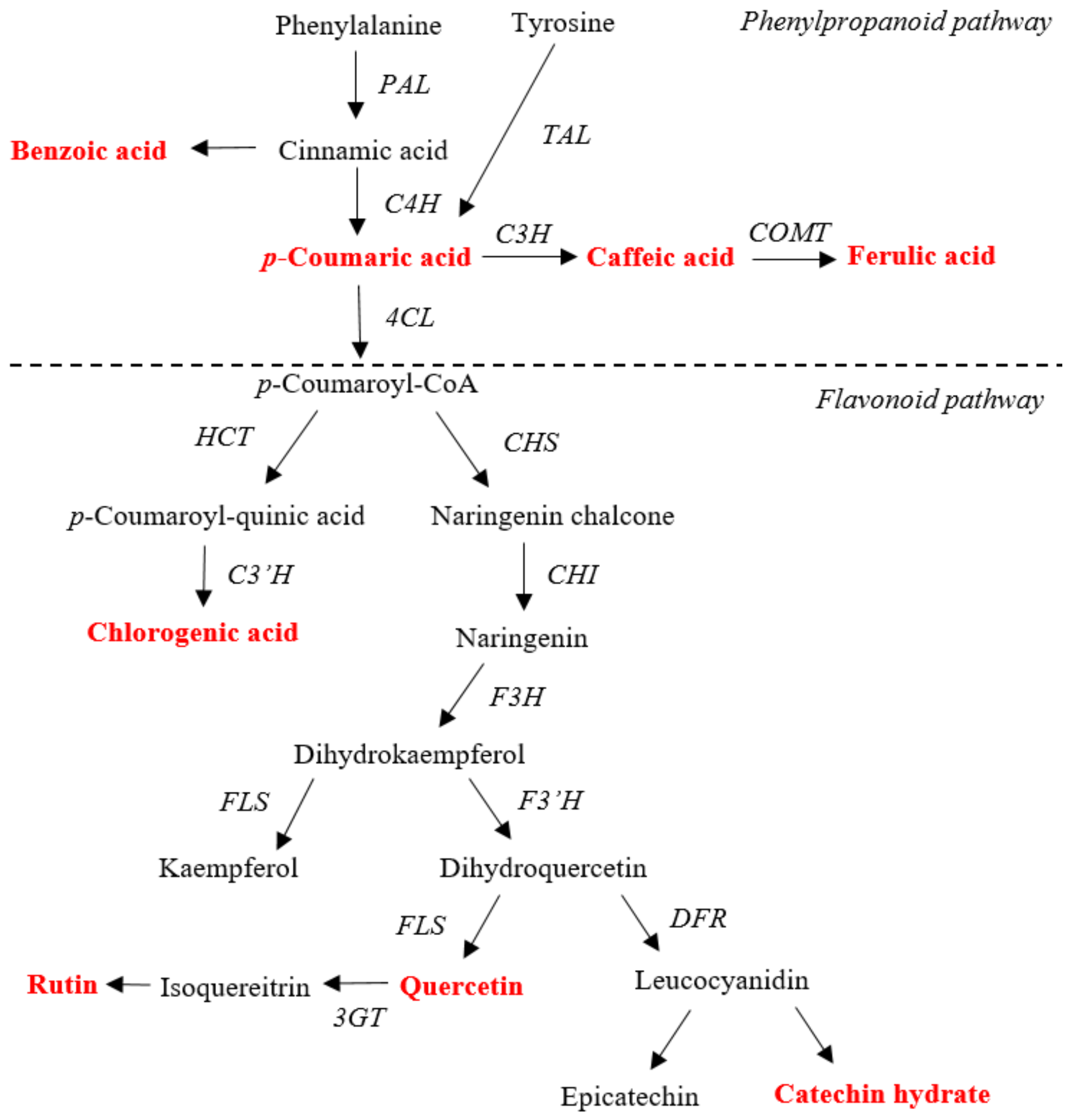
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Elicitor Treatment
2.2. Extraction Procedure of Phenolic Compounds
2.3. HPLC Analysis of Phenolic Compounds
2.4. Statistical Analysis
3. Results
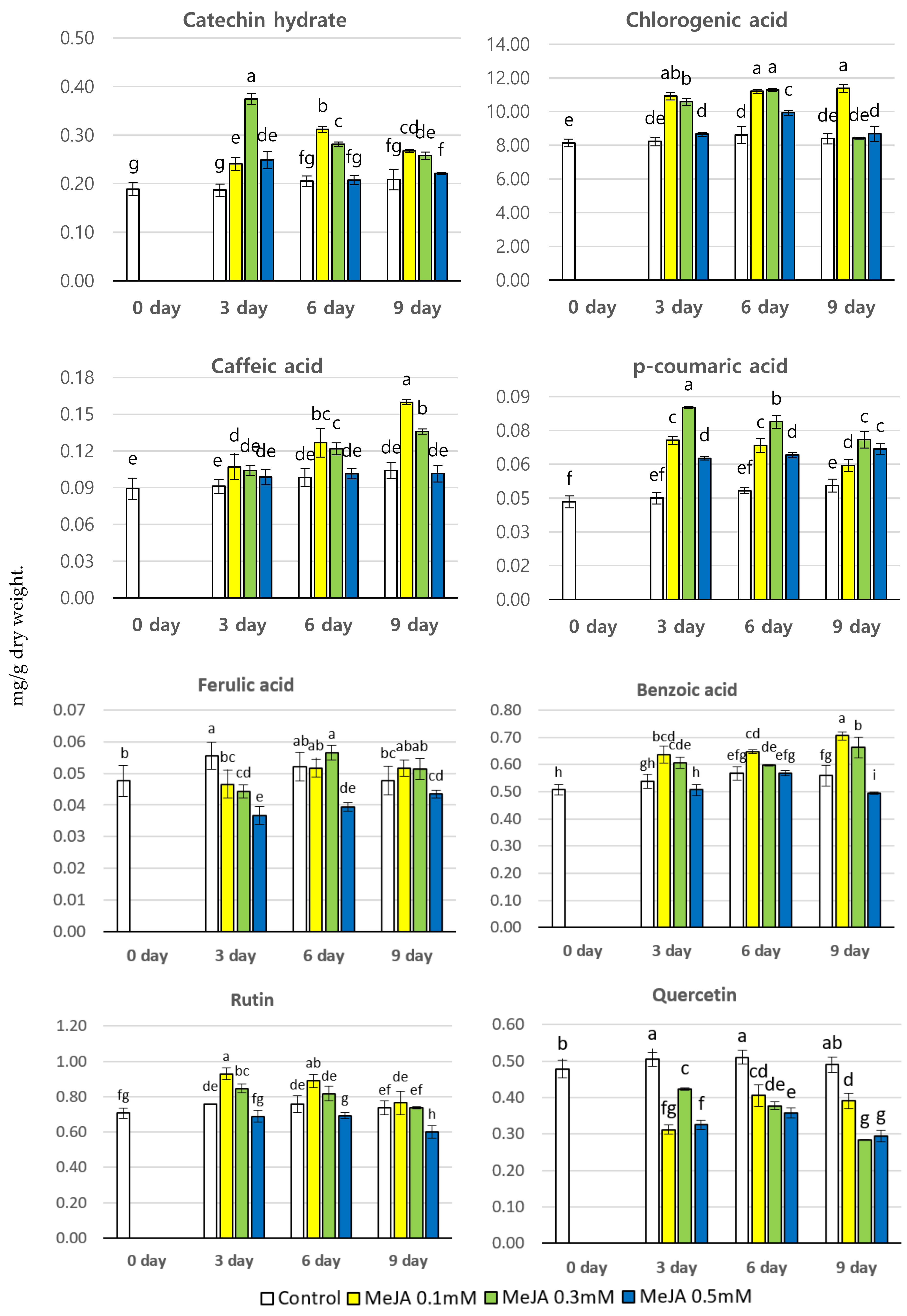
3.1. Effect of MeJA Treatment on The Phenolic Content (mg/g dry wt.) of P. brachycarpa Nakai Samples
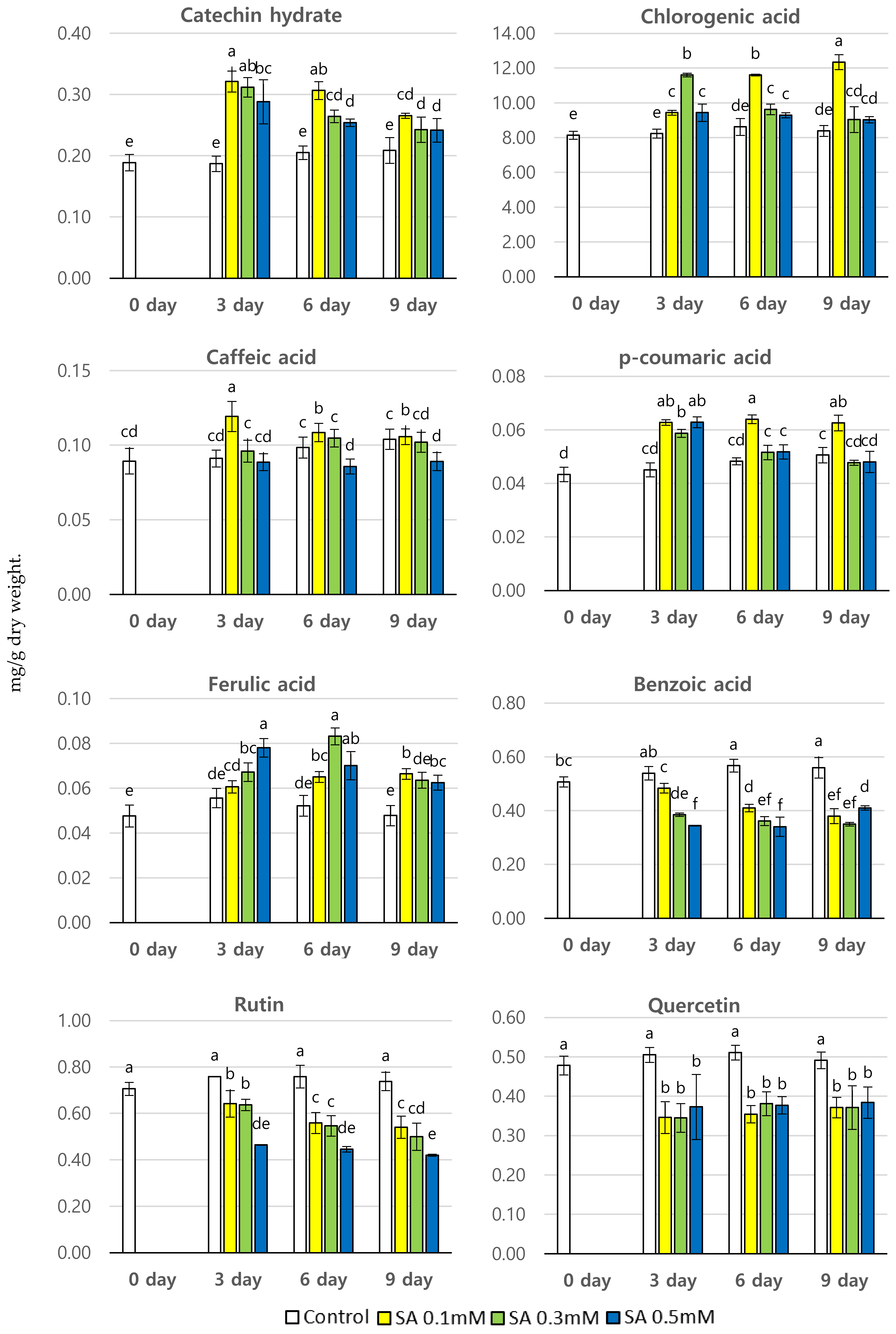
3.2. Effect of SA Treatment on The Phenolic Content (mg/g dry wt.) of P. brachycarpa Nakai Samples
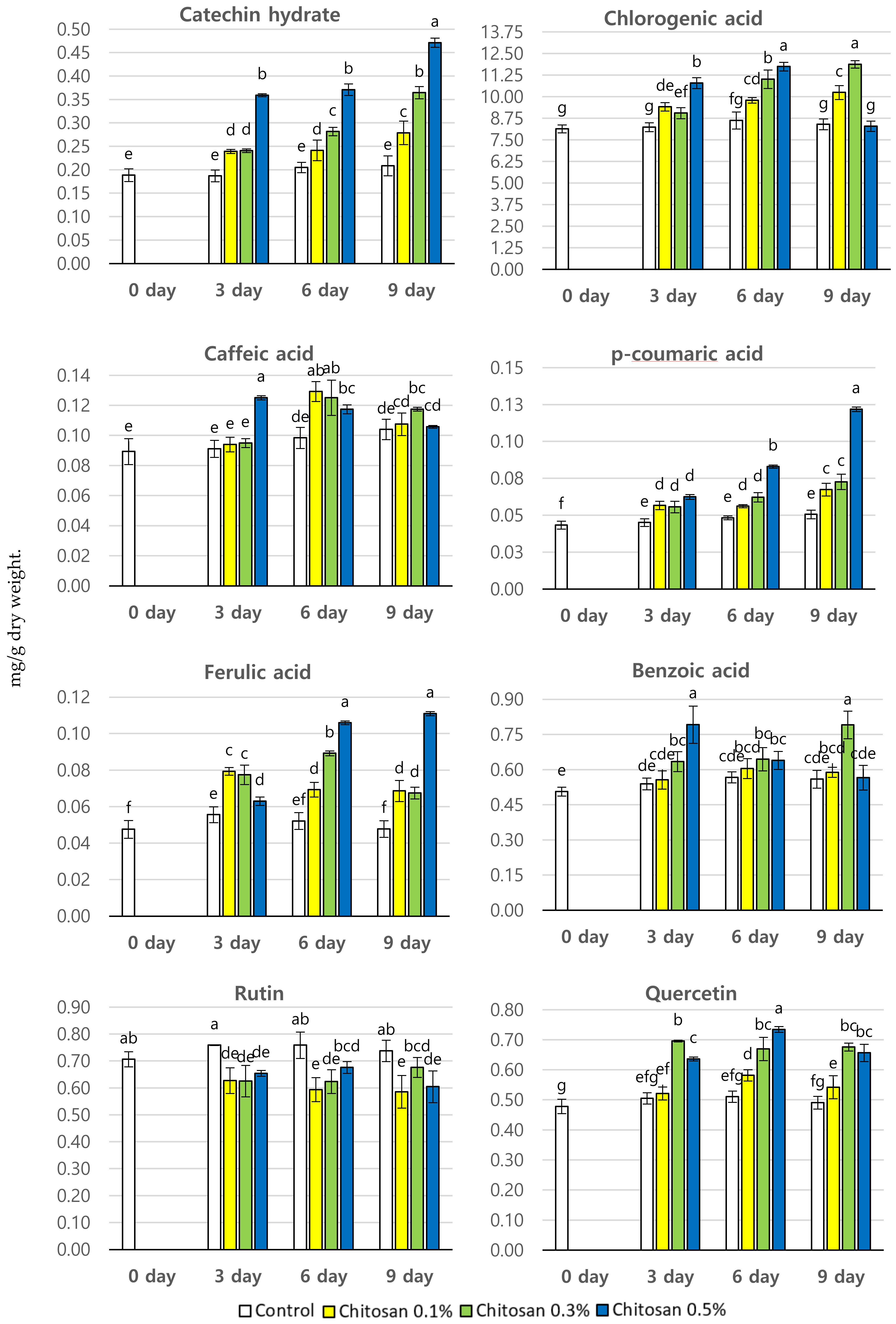
3.3. Effect of Chitosan Treatment on The Phenolic Content (mg/g dry wt.) of P. brachycarpa Nakai Samples
3.4. Comparison of The Effect of MeJA, SA, and Chitosan Treatments on The Phenolic Content (mg/g dry wt.) of P. brachycarpa Nakai Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lee, C. Coloured Flora of Korean Medicinal Herbs; Koy-Hak Publishing, Co.: Seoul, Korea, 1998; pp. 430–436. [Google Scholar]
- Ahn, S.-M.; Kim, M.-S.; Jung, I.-C.; Sohn, H.-Y. Antibacterial, antioxidative and anti-proliferative activity against human colorectal cell of Pimpinella brachycarpa. Korean J. Food Preserv. 2011, 18, 590–596. [Google Scholar]
- Lee, J.-J.; Choo, M.-H.; Lee, M.-Y. Physicochemical compositions of Pimpinella brachycarpa. J. Korean Soc. Food Sci. Nutr. 2007, 36, 327–331. [Google Scholar]
- Oh, S.-S.; Lee, J.-M.; Choi, N.-S. Change of biologically functional compounds of Pimpinella brachycarpa (Chamnamul) by blanching conditions. J. Korean Soc. Food Cult. 2001, 16, 388–397. [Google Scholar]
- Lee, S.Y.; Shin, Y.J.; Lee, K.R. Two new sesquiterpenes from the aerial parts of Pimpinella brachycarpa Nakai. Notes 2013, 34, 2215. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-J.; Choo, M.-H.; Lee, M.-Y. Effect of Pimpinella brachycarpa extract on lipid metabolism in rats fed high cholesterol diet. J. Korean Soc. Food Sci. Nutr. 2006, 35, 1151–1158. [Google Scholar]
- Chang, K. Cookery study of Pimpinella brachycarpa for Practical Utilization. Ph.D. Thesis, Duksung Women’s University, Seoul, Korea, 2003. [Google Scholar]
- Kwon, C.; Kwon, Y.; Kim, Y.; Kwon, G.; Jin, I.; Ryu, G.; Sohn, H. Inhibitory activities of edible and medicinal herbs against human thrombin. J. Life Sci. 2004, 14, 509–513. [Google Scholar]
- Choi, M.-H.; Kim, G.-H. A study on quality characteristics of Pimpinella brachycarpa Kimchi during storage at different temperatures. J. Korean Soc. Food Sci. Nutr. 2002, 31, 45–49. [Google Scholar]
- Park, S.; Jeong, G. The Use and Efficacy of Korean Folk Vegetables; Seowon Publishing, Co.: Daegu, Korea, 1995; pp. 204–205. [Google Scholar]
- Lattanzio, V. Phenolic Compounds: Introduction 50. Nat. Prod. 2013, 1543–1580. [Google Scholar] [CrossRef]
- Tossi, V.; Amenta, M.; Lamattina, L.; Cassia, R. Retracted: Nitric oxide enhances plant ultraviolet-B protection up-regulating gene expression of the phenylpropanoid biosynthetic pathway. Plant Cell Environ. 2011, 34, 909–921. [Google Scholar]
- Singh, A.; Sarma, B.K.; Upadhyay, R.S.; Singh, H.B. Compatible rhizosphere microbes mediated alleviation of biotic stress in chickpea through enhanced antioxidant and phenylpropanoid activities. Microbiol. Res. 2013, 168, 33–40. [Google Scholar]
- Critchfield, J.W.; Butera, S.T.; Folks, T.M. Inhibition of HIV activation in latently infected cells by flavonoid compounds. AIDS Res. Hum. Retrovir. 1996, 12, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Ferrazzano, G.F.; Amato, I.; Ingenito, A.; De Natale, A.; Pollio, A. Anti-cariogenic effects of polyphenols from plant stimulant beverages (cocoa, coffee, tea). Fitoterapia 2009, 80, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.-C.; Hsu, C.-L.; Yen, G.-C. Anti-inflammatory effects of phenolic compounds isolated from the fruits of Artocarpus heterophyllus. J. Agric. Food Chem. 2008, 56, 4463–4468. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef]
- Cook, N.C.; Samman, S. Flavonoids—chemistry, metabolism, cardioprotective effects, and dietary sources. J. Nutr. Biochem. 1996, 7, 66–76. [Google Scholar] [CrossRef]
- Cuong, D.M.; Kwon, S.J.; Jeon, J.; Park, Y.J.; Park, J.S.; Park, S.U. Identification and characterization of phenylpropanoid biosynthetic genes and their accumulation in bitter melon (Momordica charantia). Molecules 2018, 23, 469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.R.; Ravishankar, G. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar]
- Gadzovska, S.; Maury, S.; Delaunay, A.; Spasenoski, M.; Joseph, C.; Hagege, D. Jasmonic acid elicitation of Hypericum perforatum L. cell suspensions and effects on the production of phenylpropanoids and naphtodianthrones. Plant Cell Tissue Organ Cult. 2007, 89, 1–13. [Google Scholar] [CrossRef]
- Chakraborty, M.; Karun, A.; Mitra, A. Accumulation of phenylpropanoid derivatives in chitosan-induced cell suspension culture of Cocos nucifera. J. Plant Physiol. 2009, 166, 63–71. [Google Scholar] [CrossRef]
- Guo, J.; Wang, M.-H. Ultraviolet A-specific induction of anthocyanin biosynthesis and PAL expression in tomato (Solanum lycopersicum L.). Plant Growth Regul. 2010, 62, 1–8. [Google Scholar] [CrossRef]
- Brechner, M.L.; Albright, L.D.; Weston, L.A. Effects of UV-B on secondary metabolites of St. John’s Wort (Hypericum perforatum L.) grown in controlled environments. Photochem. Photobiol. 2011, 87, 680–684. [Google Scholar] [CrossRef]
- Flores, H.E.; Curtis, W.R. Approaches to Understanding and Manipulating the Biosynthetic Potential of Plant Roots a. Ann. N. Y. Acad. Sci. USA 1992, 665, 188–209. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.J.; Chang, H.N.; Liu, J.R.; Jung, K.H. Production and secretion of indole alkaloids in hairy root cultures of Catharanthus roseus: Effects of in situ adsorption, fungal elicitation and permeabilization. J. Ferment. Bioeng. 1994, 78, 229–234. [Google Scholar]
- Bhagyalakshmi, N.; Bopanna, K. Elicitation and immobilization of cell cultures for enhanced synthesis of pharmaceutical compounds. In Role of Biotechnology in Medicinal and Aromatic Plants; Ukaaz Publications: Hyderabad, India, 1998; Volume 1, pp. 305–325. [Google Scholar]
- Singh, G. Elicitation—Manipulating and enhancing secondary metabolite production. In Plant Cell and Tissue Culture for the Production of Food Ingredients; Springer: Berlin/Heidelberg, Germany, 1999; pp. 101–111. [Google Scholar]
- Uddin, M.R.; Park, K.W.; Kim, Y.K.; Park, S.U.; Pyon, J.Y. Enhancing sorgoleone levels in grain sorghum root exudates. J. Chem. Ecol. 2010, 36, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.R.; Park, W.T.; Kim, Y.K.; Pyon, J.Y.; Park, S.-U. Effects of auxins on sorgoleone accumulation and genes for sorgoleone biosynthesis in sorghum roots. J. Agric. Food Chem. 2011, 59, 12948–12953. [Google Scholar] [CrossRef]
- Uddin, M.R.; Thwe, A.A.; Kim, Y.B.; Park, W.T.; Chae, S.C.; Park, S.U. Effects of jasmonates on sorgoleone accumulation and expression of genes for sorgoleone biosynthesis in sorghum roots. J. Chem. Ecol. 2013, 39, 712–722. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Park, N.I.; Xu, H.; Woo, S.-H.; Park, C.H.; Park, S.U. Differential expression of flavonoid biosynthesis genes and accumulation of phenolic compounds in common buckwheat (Fagopyrum esculentum). J. Agric. Food Chem. 2010, 58, 12176–12181. [Google Scholar] [CrossRef]
- Uddin, M.R.; Li, X.; Park, W.T.; Kim, Y.B.; Kim, S.J.; Kim, Y.S.; Lee, M.Y.; Park, C.H.; Park, S.U. Phenolic compound content in different organs of Korean common buckwheat cultivars. Asian J. Chem. 2013, 25, 424. [Google Scholar] [CrossRef]
- Uddin, M.R.; Li, X.; Kim, Y.B.; Chae, S.C.; Kim, S.-J.; Park, S.U. Phenolic compounds in different organs of tartary buckwheat (Fagopyrum tataricum Gaertn.) cultivars. Aust. J. Crop Sci. 2013, 7, 1861–1865. [Google Scholar]
- Kim, Y.K.; Li, X.; Xu, H.; Park, N.I.; Uddin, M.R.; Pyon, J.Y.; Park, S.U. Production of phenolic compounds in hairy root culture of tartary buckwheat (Fagopyrum tataricum Gaertn). J. Crop Sci. Biotechnol. 2009, 12, 53–57. [Google Scholar]
- Kim, Y.B.; Kim, J.K.; Uddin, M.R.; Xu, H.; Park, W.T.; Tuan, P.A.; Li, X.; Chung, E.; Lee, J.-H.; Park, S.U. Metabolomics analysis and biosynthesis of rosmarinic acid in Agastache rugosa Kuntze treated with methyl jasmonate. PLoS ONE 2013, 8, e64199. [Google Scholar] [CrossRef] [Green Version]
- Won, O.; Uddin, M.; Park, K.; Pyon, J.; Park, S. Phenolic compounds in sorghum leaf extracts and their effects on weed control. Allelopath. J. 2013, 31, 147–156. [Google Scholar]
- Im, A.; Kim, Y.-H.; Uddin, M.; Lee, H.W.; Chae, S.W.; Kim, Y.H.; Jung, W.S.; Kang, B.J.; Mun, C.S.; Lee, M.-Y. Scutellaria baicalensis extracts and flavonoids protect rat L6 cells from antimycin A-induced mitochondrial dysfunction. Evid. Based Complementary Altern. Med. 2012, 2012, 517965. [Google Scholar] [CrossRef] [Green Version]
- Park, W.T.; Kim, J.K.; Park, S.; Lee, S.-W.; Li, X.; Kim, Y.B.; Uddin, M.R.; Park, N.I.; Kim, S.-J.; Park, S.U. Metabolic profiling of glucosinolates, anthocyanins, carotenoids, and other secondary metabolites in kohlrabi (Brassica oleracea var. gongylodes). J. Agric. Food Chem. 2012, 60, 8111–8116. [Google Scholar]
- Chen, H.; Seguin, P.; Archambault, A.; Constan, L.; Jabaji, S. Gene expression and isoflavone concentrations in soybean sprouts treated with chitosan. Crop Sci. 2009, 49, 224–236. [Google Scholar]
- Park, W.T.; Kim, Y.B.; Seo, J.M.; Kim, S.-J.; Chung, E.; Lee, J.-H.; Park, S.U. Accumulation of anthocyanin and associated gene expression in radish sprouts exposed to light and methyl jasmonate. J. Agric. Food Chem. 2013, 61, 4127–4132. [Google Scholar] [CrossRef]
- Skrzypczak-Pietraszek, E.; Słota, J.; Pietraszek, J. The influence of L-phenylalanine, methyl jasmonate and sucrose concentration on the accumulation of phenolic acids in Exacum affine Balf. f. ex Regel shoot culture. Acta Biochim. Pol. 2014, 61, 61. [Google Scholar] [CrossRef]
- Sun, Z.; Hou, S.; Yang, W.; Han, Y. Exogenous application of salicylic acid enhanced the rutin accumulation and influenced the expression patterns of rutin biosynthesis related genes in Fagopyrum tartaricum Gaertn leaves. Plant Growth Regul. 2012, 68, 9–15. [Google Scholar] [CrossRef]
- Mandal, S.; Gupta, C. Inducing cell wall-bound phenolic compounds by elicitors in eggplant (Solanum melongena). Appl. Biochem. Microbiol. 2016, 52, 650–656. [Google Scholar] [CrossRef]
- Coste, A.; Vlase, L.; Halmagyi, A.; Deliu, C.; Coldea, G. Effects of plant growth regulators and elicitors on production of secondary metabolites in shoot cultures of Hypericum hirsutum and Hypericum maculatum. Plant Cell Tissue Organ Cult. 2011, 106, 279–288. [Google Scholar] [CrossRef]
- Khan, W.; Prithiviraj, B.; Smith, D.L. Chitosan and chitin oligomers increase phenylalanine ammonia-lyase and tyrosine ammonia-lyase activities in soybean leaves. J. Plant Physiol. 2003, 160, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Yeo, H.J.; Park, Y.E.; Chun, S.W.; Chung, Y.S.; Lee, S.Y.; Park, S.U. Influence of chitosan, salicylic acid and jasmonic acid on phenylpropanoid accumulation in germinated buckwheat (Fagopyrum esculentum Moench). Foods 2019, 8, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, J.; Zhao, X.; Li, Z. iTRAQ-Based Quantitative Proteomic Analysis of the Arabidopsis Mutant opr3-1 in Response to Exogenous MeJA. Int. J. Mol. Sci. 2020, 21, 571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayat, Q.; Hayat, S.; Irfan, M.; Ahmad, A. Effect of exogenous salicylic acid under changing environment: A review. Environ. Exp. Bot. 2010, 68, 14–25. [Google Scholar] [CrossRef]
- Jogaiah, S.; Satapute, P.; De Britto, S.; Konappa, N.; Udayashankar, C. Exogenous priming of chitosan induces upregulation of phytohormones and resistance against cucumber powdery mildew disease is correlated with localized biosynthesis of defense enzymes. Int. J. Biol. Macromol. 2020, 162, 1825–1838. [Google Scholar] [CrossRef]
- Farag, M.A.; Huhman, D.V.; Dixon, R.A.; Sumner, L.W. Metabolomics reveals novel pathways and differential mechanistic and elicitor-specific responses in phenylpropanoid and isoflavonoid biosynthesis in Medicago truncatula cell cultures. Plant Physiol. 2008, 146, 387–402. [Google Scholar] [CrossRef] [Green Version]
- Kubes, J.; Skalicky, M.; Tumova, L.; Martin, J.; Hejnak, V.; Martinkova, J. Vanadium elicitation of Trifolium pratense L. cell culture and possible pathways of produced isoflavones transport across the plasma membrane. Plant Cell Rep. 2019, 38, 657–671. [Google Scholar] [CrossRef]
- Reichling, J.; Merkel, B.; Hofmeister, P. Studies on the Biological Activites of Rare Phenylpropanoids of the Genus Pimpinella. J. Nat. Prod. 1991, 54, 1416–1418. [Google Scholar] [CrossRef]
- Delazar, A.; Biglari, F.; Esnaashari, S.; Nazemiyeh, H.; Talebpour, H.; Nahar, L.; Sarker, S.D. GC-MS analysis of the essential oils, and the isolation of phenylpropanoid derivatives from the aerial parts of Pimpinella aurea. Phytochemistry 2006, 67, 2176–2181. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HPLC | Futecs HPLC system NS-4000 (Futecs Co. Ltd., Daejeon, Korea) | ||
|---|---|---|---|
| Column | OptimaPak C18-51002546 (250 mm × 4.6 mm I.d., particle size 5 µm) (RStech Co. Ltd., Daejeon, Korea) | ||
| Mobile phase | Solvent A (0.15% Acetic acid water) Solvent B (Methanol) | ||
| Flow rate | 1.0 mL/min | ||
| Column temperature | 30 °C | ||
| Injection Volume | 10 μL | ||
| UV wavelength | 280 nm | ||
| Gradient elution | Time (min) | Solvent A (%) | Solvent B (%) |
| 0–1 | 95 | 5 | |
| 1–9 | 85 | 15 | |
| 9–24 | 80 | 20 | |
| 24–54 | 70 | 30 | |
| 54–65 | 55 | 45 | |
| 65–75 | 44 | 56 | |
| 75–80 | 40 | 60 | |
| 80–90 | 20 | 80 | |
| 90–105 | 95 | 5 | |
| Treatment | Total | Treatment | Total | Treatment | Total | |
|---|---|---|---|---|---|---|
| 0 Days | Control | 10.20 ± 0.20 g 1 | Control | 10.20 ± 0.20 f | Control | 10.20 ± 0.20 i |
| 3 Days | Control | 10.41 ± 0.30 fg | Control | 10.41 ± 0.30 ef | Control | 10.41 ± 0.30 hi |
| MeJA 0.1 | 13.26 ± 0.18 bc | SA 0.1 | 11.47 ± 0.01 c | Chitosan 0.1% | 11.58 ± 0.34 de | |
| MeJA 0.3 | 13.06 ± 0.28 c | SA 0.3 | 13.50 ± 0.15 b | Chitosan 0.3% | 11.47 ± 0.33 ef | |
| MeJA 0.5 | 10.63 ± 0.12 fg | SA 0.5 | 11.14 ± 0.57 cd | Chitosan 0.5% | 13.48 ± 0.30 b | |
| 6 Days | Control | 10.86 ± 0.42 e | Control | 10.86 ± 0.42 cdef | Control | 10.86 ± 0.42 g |
| MeJA 0.1 | 13.71 ± 0.14 ab | SA 0.1 | 13.47 ± 0.07 b | Chitosan 0.1% | 12.07 ± 0.23 cd | |
| MeJA 0.3 | 13.62 ± 0.10 ab | SA 0.3 | 11.42 ± 0.33 c | Chitosan 0.3% | 13.51 ± 0.60 b | |
| MeJA 0.5 | 11.96 ± 0.13 d | SA 0.5 | 10.92 ± 0.13 cde | Chitosan 0.5% | 14.47 ± 0.23 a | |
| 9 Days | Control | 10.59 ± 0.30 fg | Control | 10.59 ± 0.30 def | Control | 10.59 ± 0.30 hi |
| MeJA 0.1 | 13.78 ± 0.27 a | SA 0.1 | 14.14 ± 0.44 a | Chitosan 0.1% | 12.48 ± 0.34 c | |
| MeJA 0.3 | 10.63 ± 0.02 fg | SA 0.3 | 10.71 ± 0.86 def | Chitosan 0.3% | 14.63 ± 0.33 a | |
| MeJA 0.5 | 10.50 ± 0.47 fg | SA 0.5 | 10.68 ± 0.16 def | Chitosan 0.5% | 10.92 ± 0.31 fg | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, N.S.; Jung, D.H.; Jung, C.R.; Jeon, K.S.; Park, H.W.; Park, S.U. Improvement of Phenylpropanoid Production with Elicitor Treatments in Pimpinella brachycarpa Nakai. Horticulturae 2020, 6, 108. https://doi.org/10.3390/horticulturae6040108
Kim NS, Jung DH, Jung CR, Jeon KS, Park HW, Park SU. Improvement of Phenylpropanoid Production with Elicitor Treatments in Pimpinella brachycarpa Nakai. Horticulturae. 2020; 6(4):108. https://doi.org/10.3390/horticulturae6040108
Chicago/Turabian StyleKim, Nam Su, Dae Hui Jung, Chung Ryul Jung, Kwon Seok Jeon, Hong Woo Park, and Sang Un Park. 2020. "Improvement of Phenylpropanoid Production with Elicitor Treatments in Pimpinella brachycarpa Nakai" Horticulturae 6, no. 4: 108. https://doi.org/10.3390/horticulturae6040108
APA StyleKim, N. S., Jung, D. H., Jung, C. R., Jeon, K. S., Park, H. W., & Park, S. U. (2020). Improvement of Phenylpropanoid Production with Elicitor Treatments in Pimpinella brachycarpa Nakai. Horticulturae, 6(4), 108. https://doi.org/10.3390/horticulturae6040108