Salinity-Induced Physiological Responses of Three Putative Salt Tolerant Citrus Rootstocks



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth and Salt Treatment
2.2. Chlorophyll and Gas Exchange Measurements
2.3. Malondialdehyde Determination
2.4. Total Phenolic Content (TPC) Determination
2.5. Proline Content Determination
2.6. Sodium and Chloride Determination
2.7. Experimental Design and Data Analysis
3. Results and Discussion
3.1. Morphological Changes in Citrus Rootstocks in Response to Salinity Stress
3.2. Effect of Salinity Stress in Physiological Changes in Citrus Rootstocks
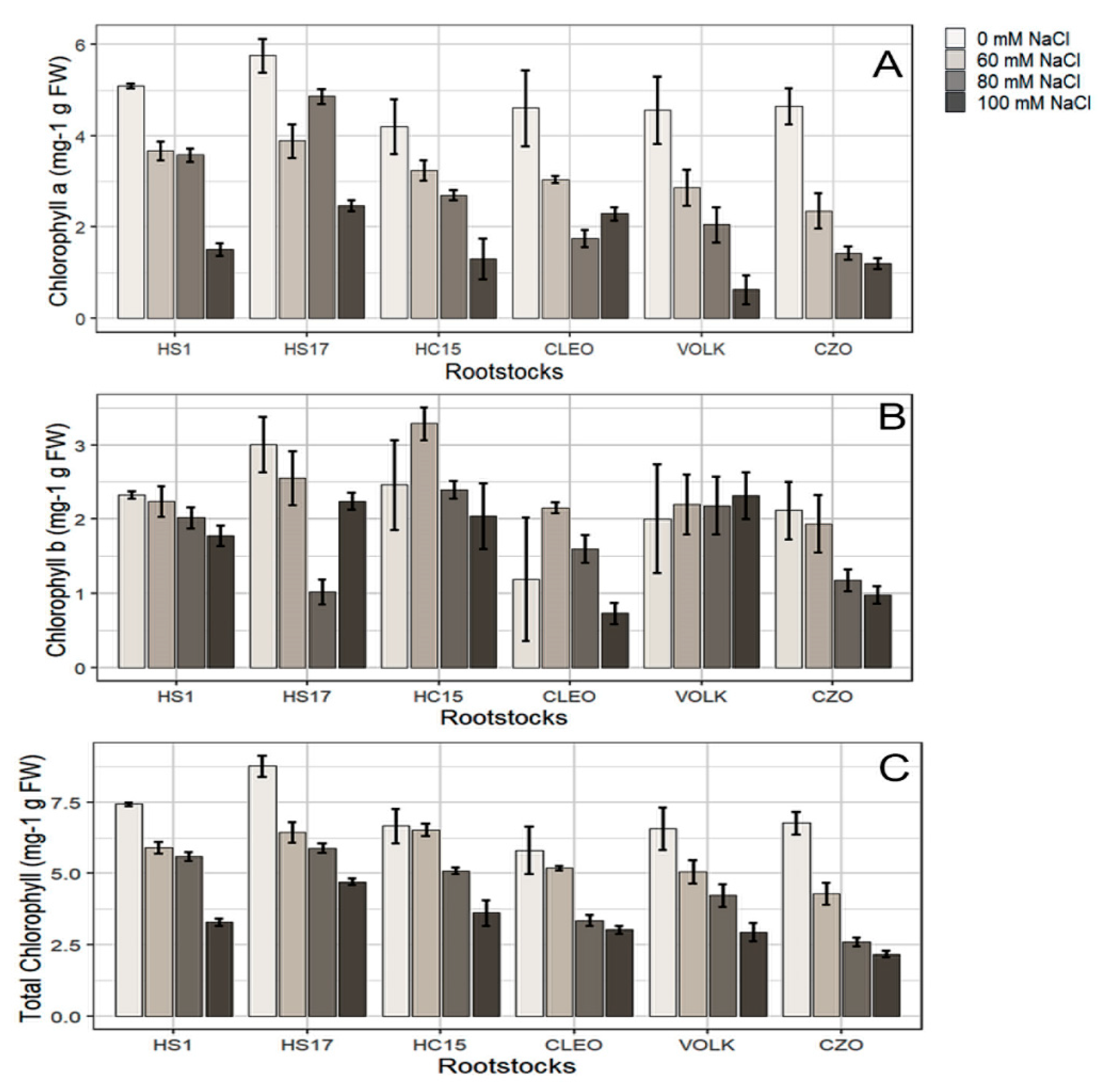
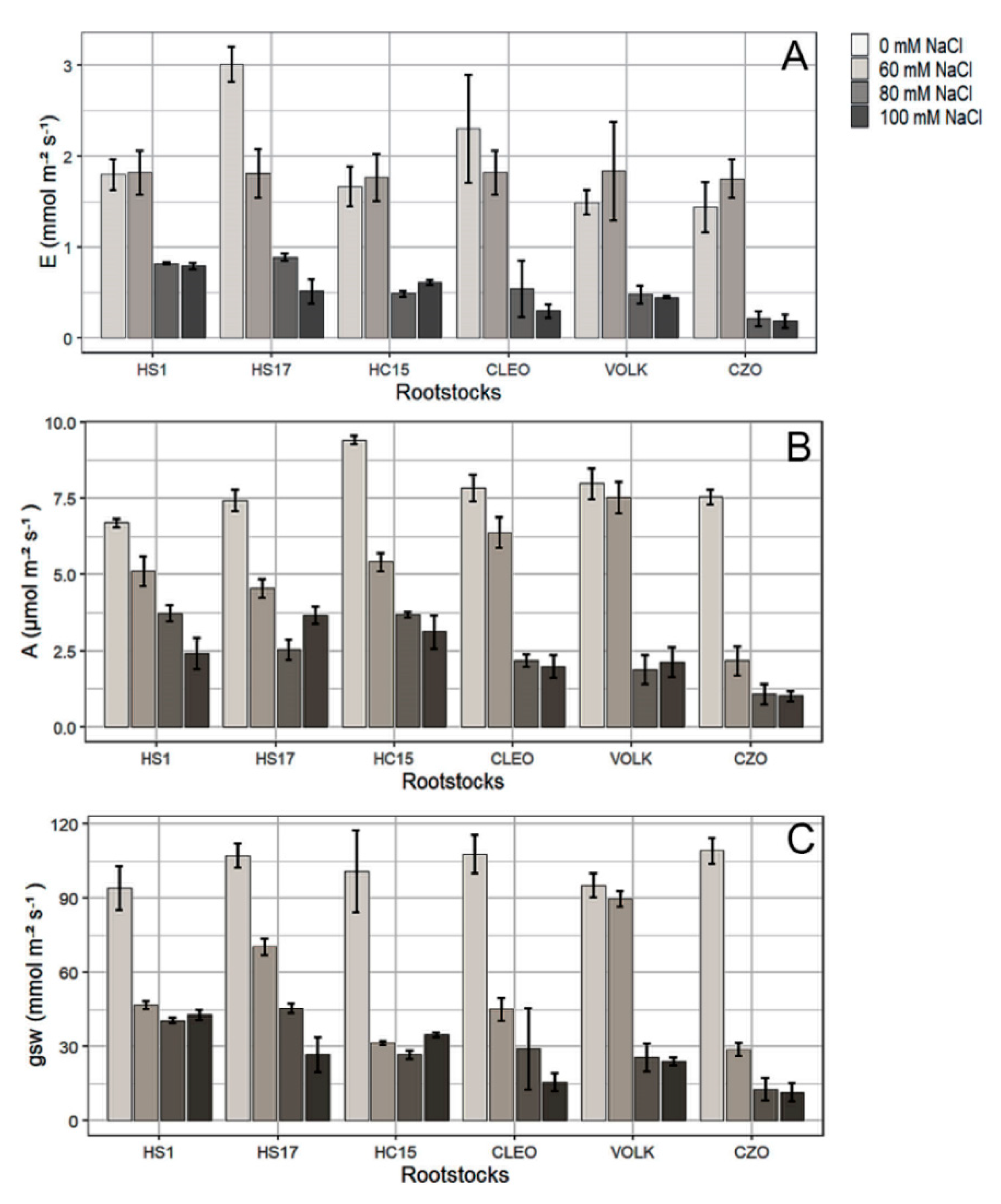
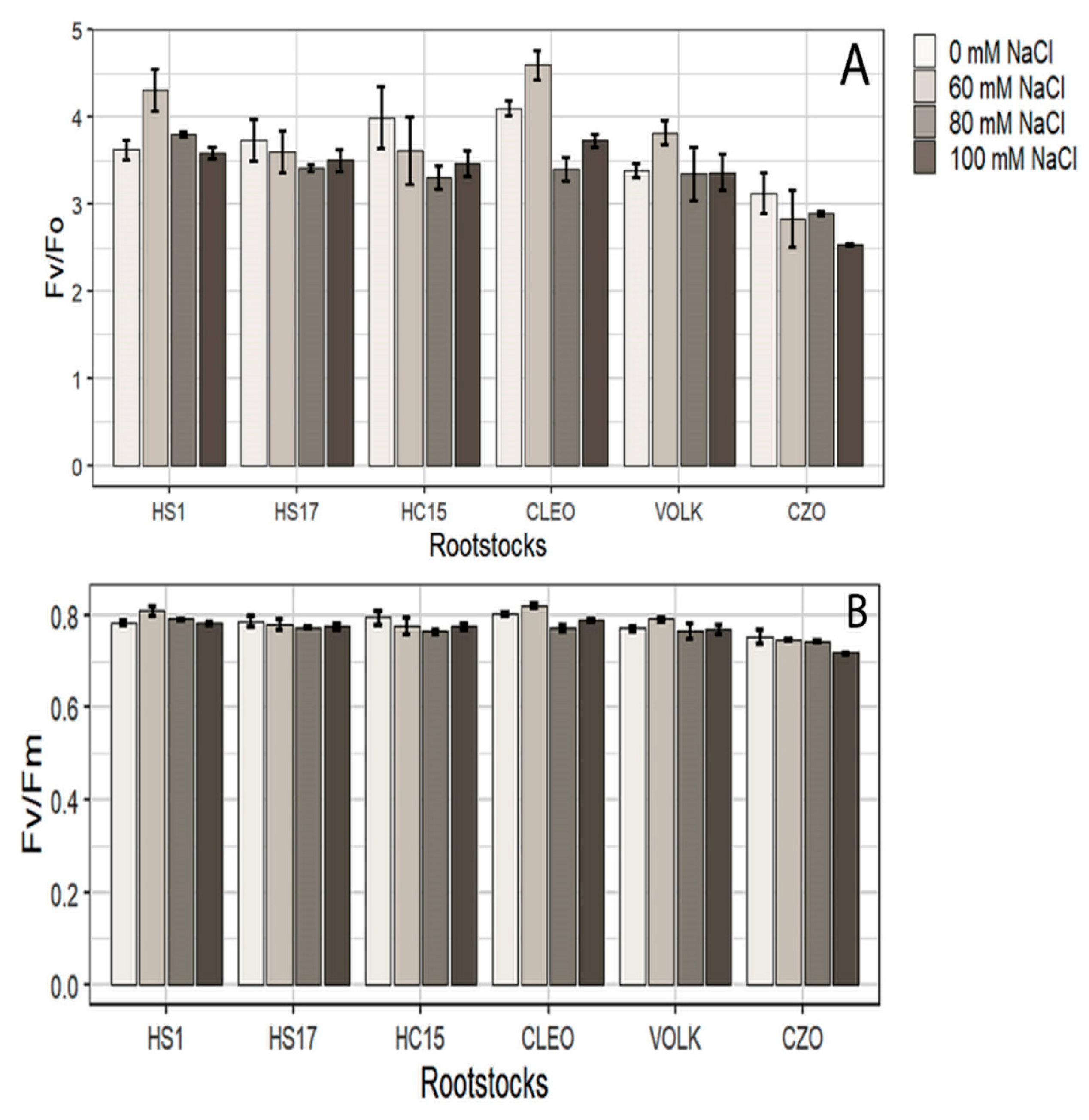
3.2.1. Photosynthesis and Chlorophyll Content
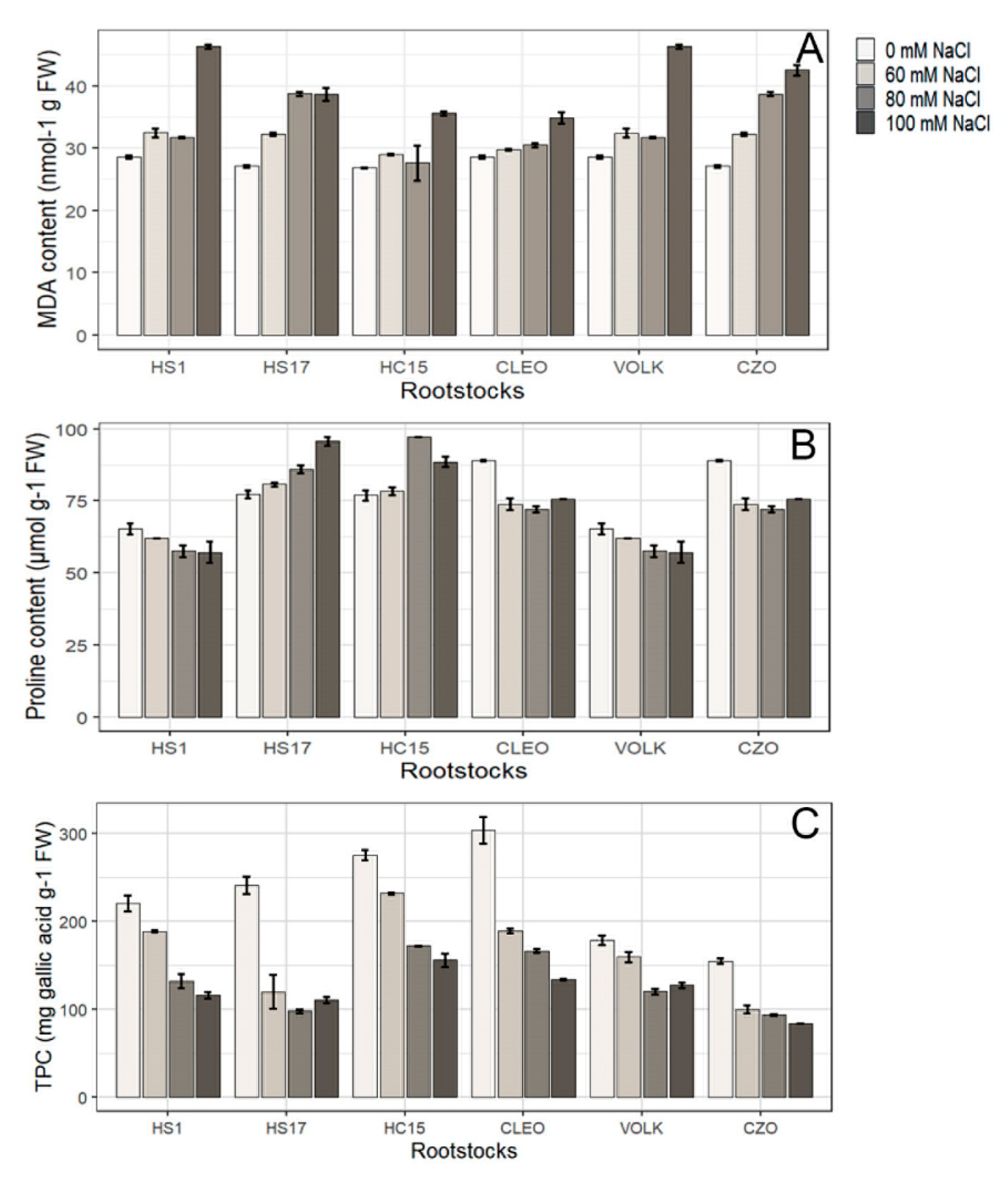
3.2.2. Lipid Peroxidation
3.2.3. Proline Content
3.2.4. Total Phenolic Compounds
3.3. Sodium and Chloride Ions Analysis
3.4. Correlation Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vincent, C.; Morillon, R.; Arbona, V.; Gómez-Cadenas, A. Citrus in changing environments. In The Genus Citrus; Elsevier: Amsterdam, The Netherlands, 2020; pp. 271–289. [Google Scholar]
- Syvertsen, J.P.; Garcia-Sanchez, F. Multiple abiotic stresses occurring with salinity stress in citrus. Environ. Exp. Bot. 2014, 103, 128–137. [Google Scholar] [CrossRef]
- Maas, E. Salinity and citriculture. Tree Physiol. 1993, 12, 195–216. [Google Scholar] [CrossRef]
- Storey, R.; Walker, R. Citrus and salinity. Sci. Hortic. 1998, 78, 39–81. [Google Scholar] [CrossRef]
- Mahmoud, L.M.; Dutt, M.; Shalan, A.M.; El-Kady, M.E.; El-Boray, M.S.; Shabana, Y.M.; Grosser, J.W. Silicon nanoparticles mitigate oxidative stress of in vitro-derived banana (Musa acuminata ‘Grand Nain’) under simulated water deficit or salinity stress. S. Afr. J. Bot. 2020, 132, 155–163. [Google Scholar] [CrossRef]
- Cunningham, M.A.; Snyder, E.; Yonkin, D.; Ross, M.; Elsen, T. Accumulation of deicing salts in soils in an urban environment. Urban Ecosyst. 2008, 11, 17–31. [Google Scholar] [CrossRef]
- Mugai, E.N. Salinity characterization of the Kenyan saline soils. Soil Sci. Plant Nutr. 2004, 50, 181–188. [Google Scholar] [CrossRef]
- Vincent, C.; Rowland, D.; Schaffer, B.; Bassil, E.; Racette, K.; Zurweller, B. Primed acclimation: A physiological process offers a strategy for more resilient and irrigation-efficient crop production. Plant Sci. 2020, 295, 110240. [Google Scholar] [CrossRef]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef]
- Abel, G. Inheritance of the Capacity for Chloride Inclusion and Chloride Exclusion by Soybeans 1. Crop Sci. 1969, 9, 697–698. [Google Scholar] [CrossRef]
- Sykes, S. The inheritance of salt exclusion in woody perennial fruit species. Plant Soil 1992, 146, 123–129. [Google Scholar] [CrossRef]
- Glenn, E.P.; Brown, J.J.; Blumwald, E. Salt tolerance and crop potential of halophytes. Crit. Rev. Plant Sci. 1999, 18, 227–255. [Google Scholar] [CrossRef]
- Ratner, A.; Jacoby, B. Non-specificity of salt effects on Mg2+ dependent ATPase from grass roots. J. Exp. Bot. 1973, 24, 231–238. [Google Scholar] [CrossRef]
- Rains, D.W. Sodium and potassium absorption by bean stem tissue. Plant Physiol. 1969, 44, 547–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, E.; Preston, J. Salt tolerance of Trifolium alexandrinum L. IV. Ion measurements by X-ray microanalysis in unfixed, frozen hydrated leaf cells at various stages of salt treatment. Funct. Plant Biol. 1982, 9, 251–259. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Env. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Shabala, S.; Bose, J.; Hedrich, R. Salt bladders: Do they matter? Trends Plant Sci. 2014, 19, 687–691. [Google Scholar] [CrossRef]
- Greenway, H.; Munns, R. Mechanisms of salt tolerance in nonhalophytes. Annu. Rev. Plant Physiol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Cooper, W.C.; Gorton, B.; Olson, E. Ionic accumulation in citrus as influenced by rootstock and scion and concentration of salts and boron in the substrate. Plant Physiol. 1952, 27, 191. [Google Scholar] [CrossRef] [Green Version]
- Grosser, J.; Gmitter, F.; Tusa, N.; Chandler, J. Somatic hybrid plants from sexually incompatible woody species: Citrus reticulata and Citropsis gilletiana. Plant Cell Rep. 1990, 8, 656–659. [Google Scholar] [CrossRef]
- Brlansky, R.; Lee, R.; Agostini, J.P. The occurrence of citrus blight on sweet orange and cleopatra mandarin rootstocks in northwestern Argentina. Microbiol. Plant Pathol. 1988, 10, 381–383. [Google Scholar]
- Chellemi, D.; Sonoda, R.; Pelosi, R.; Cohen, M. Temporal and spatial comparisons between epidemics of citrus blight and citrus tristeza virus. Microbiol. Plant Pathol. 1991, 11, 289–296. [Google Scholar]
- Alam, A.; Ullah, H.; Attia, A.; Datta, A. Effects of salinity stress on growth, mineral nutrient accumulation and biochemical parameters of seedlings of three citrus rootstocks. Int. J. Fruit Sci. 2020, 20, 786–804. [Google Scholar] [CrossRef]
- Awang, Y.; Shafieizar, A.; Ajamgard, F.; Juraimi, A.S.; Othman, R.; Ahmadi, A.K. Assessing five citrus rootstocks for NaCl salinity tolerance using mineral concentrations, proline and relative water contents as indicators. Asian J. Plant Sci. 2015, 14, 20–26. [Google Scholar] [CrossRef]
- Etehadpour, M.; Fatahi, R.; Zamani, Z.; Golein, B.; Naghavi, M.R.; Gmitter, F. Evaluation of the salinity tolerance of Iranian citrus rootstocks using morph-physiological and molecular methods. Sci. Hortic. 2020, 261, 109012. [Google Scholar] [CrossRef]
- Khalid, M.F.; Hussain, S.; Anjum, M.A.; Ahmad, S.; Ali, M.A.; Ejaz, S.; Morillon, R. Better salinity tolerance in tetraploid vs diploid volkamer lemon seedlings is associated with robust antioxidant and osmotic adjustment mechanisms. J. Plant Physiol. 2020, 244, 153071. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, H.; Yoshikawa, H.; Usuki, T. Extraction of Nobiletin and Tangeretin From Peels of Shekwasha and Ponkan Using [C2mim][(MeO)(H) PO2] and Centrifugation. Nat. Prod. Commun. 2019, 14. [Google Scholar] [CrossRef] [Green Version]
- Swingle, W.R.; Reece, P. The Botany Of Citrus and Its Wild Relatives. In The Citrus Industry; Reuther, W.W., Batchelor, L.D., Webber, H.J., Eds.; University of California Press: Berkeley, CA, USA, 1967; Volume I, pp. 190–340. [Google Scholar]
- Sun, Y.; Niu, G.; Perez, C. Relative salt tolerance of seven Texas Superstar® perennials. HortScience 2015, 50, 1562–1566. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of Total Carotenoids and Chlorophylls A and B of Leaf Extracts in Different Solvents; Portland Press Limited: London, UK, 1983. [Google Scholar]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Anderson, D.; Henderson, L. Comparing sealed chamber digestion with other digestion methods used for plant-tissue analysis. Agron. J. 1988, 80, 549–552. [Google Scholar] [CrossRef]
- Munter, R.; Halverson, T.; Anderson, R. Quality assurance for plant tissue analysis by ICP-AES. Commun. Soil Sci. Plant Anal. 1984, 15, 1285–1322. [Google Scholar] [CrossRef]
- Anjum, M.A. Effect of NaCl concentrations in irrigation water on growth and polyamine metabolism in two citrus rootstocks with different levels of salinity tolerance. Acta Physiol. Plant. 2008, 30, 43. [Google Scholar] [CrossRef]
- López-Climent, M.F.; Arbona, V.; Pérez-Clemente, R.M.; Gómez-Cadenas, A. Relationship between salt tolerance and photosynthetic machinery performance in citrus. Environ. Exp. Bot. 2008, 62, 176–184. [Google Scholar] [CrossRef]
- Hussain, S.; Luro, F.; Costantino, G.; Ollitrault, P.; Morillon, R. Physiological analysis of salt stress behaviour of citrus species and genera: Low chloride accumulation as an indicator of salt tolerance. S. Afr. J. Bot. 2012, 81, 103–112. [Google Scholar] [CrossRef] [Green Version]
- García-Sánchez, F.; Jifon, J.L.; Carvajal, M.; Syvertsen, J.P. Gas exchange, chlorophyll and nutrient contents in relation to Na+ and Cl− accumulation in ‘Sunburst’mandarin grafted on different rootstocks. Plant Sci. 2002, 162, 705–712. [Google Scholar] [CrossRef]
- Ruiz, M.; Quiñones, A.; Martínez-Cuenca, M.R.; Aleza, P.; Morillon, R.; Navarro, L.; Primo-Millo, E.; Martínez-Alcántara, B. Tetraploidy enhances the ability to exclude chloride from leaves in Carrizo citrange seedlings. J. Plant Physiol. 2016, 205, 1–10. [Google Scholar] [CrossRef]
- Khoshbakht, D.; Asghari, M.; Haghighi, M. Effects of foliar applications of nitric oxide and spermidine on chlorophyll fluorescence, photosynthesis and antioxidant enzyme activities of citrus seedlings under salinity stress. Photosynthetica 2018, 56, 1313–1325. [Google Scholar] [CrossRef]
- Hu, Y.; Huang, Y.; Zhou, S.; Zhang, Y.; Cheng, R.; Guo, J.; Ling, Y. Traditional rice landraces in Lei-Qiong area of South China tolerate salt stress with strong antioxidant activity. Plant Signal. Behav. 2020, 15, 1740466. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Torabian, S. Antioxidant enzyme and osmotic adjustment changes in bean seedlings as affected by biochar under salt stress. Ecotoxicol. Environ. Saf. 2017, 137, 64–70. [Google Scholar] [CrossRef]
- Rivero, R.M.; Ruiz, J.M.; Garcıa, P.C.; Lopez-Lefebre, L.R.; Sánchez, E.; Romero, L. Resistance to cold and heat stress: Accumulation of phenolic compounds in tomato and watermelon plants. Plant Sci. 2001, 160, 315–321. [Google Scholar] [CrossRef]
- Hussain, S.; Khalid, M.F.; Saqib, M.; Ahmad, S.; Zafar, W.; Rao, M.J.; Morillon, R.; Anjum, M.A. Drought tolerance in citrus rootstocks is associated with better antioxidant defense mechanism. Acta Physiol. Plant. 2018, 40, 135. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Moya, J.L.; Gómez-Cadenas, A.; Primo-Millo, E.; Talon, M. Chloride absorption in salt-sensitive Carrizo citrange and salt-tolerant Cleopatra mandarin citrus rootstocks is linked to water use. J. Exp. Bot. 2003, 54, 825–833. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, J.; Kriedemann, P.; Aspinall, D. Contrasts between Citrus species in response to salinisation: An analysis of photosynthesis and water relations for different rootstock-scion combinations. Physiol. Plant. 1990, 78, 236–246. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rootstocks | NaCl Treatments | |||
|---|---|---|---|---|
| 0 mm NaCl | 60 mm NaCl | 80 mm NaCl | 100 mm NaCl | |
| HS1 | 5.00 ± 0.00 a* | 5.00 ± 0.00 a | 4.75 ± 0.19 a | 3.50 ± 0.28 b,c |
| HS17 | 5.00 ± 0.00 a | 5.00 ± 0.00 a | 4.75 ± 0.19 a | 4.25 ± 0.21 a,b |
| HC15 | 5.00 ± 0.00 a | 5.00 ± 0.00 a | 4.75 + 0.19 a | 4.75 ± 0.20 a |
| Cleo | 5.00 ± 0.00 a | 5.00 ± 0.00 a | 5.00 ± 0.00 a | 4.75 ± 0.19 a |
| Volk | 5.00 ± 0.00 a | 5.00 ± 0.00 a | 5.00 ± 0.00 a | 3.00 ± 0.00 c |
| Czo | 5.00 ± 0.00 a | 1.75 ± 0.30 c | 0.75 ± 0.40 d,e | 0.00 ± 0.00 e |
| Rootstocks | NaCl Treatments | Combination | |
|---|---|---|---|
| Chl a | 24.2 ***a | 140.5 *** | 3.9 *** |
| Chl b | 11.8 *** | 11.6 *** | 3.3 *** |
| TChl | 26.3 *** | 117.2 *** | 2.1 * |
| Fv/Fo | 15.8 *** | 7.5 *** | 1.6 ns |
| Fv/Fm | 20.2 *** | 7.8 *** | 1.5 ns |
| E | 3.7 ** | 67.8 *** | 1.6 ns |
| A | 18.7 *** | 275.3 *** | 7.7 *** |
| Gsw | 6.3 *** | 185.6 *** | 5.1 *** |
| MDA | 35.3 *** | 294.4 *** | 18.8 *** |
| TPC | 125.6 *** | 318.3 *** | 12.2 *** |
| Proline | 133.0 *** | 13.4 *** | 12.3 *** |
| Na (Leaves) | 13.4 *** | 633.5 *** | 10.6 *** |
| Na (Roots) | 13.5 *** | 409.0 *** | 5.0 *** |
| Cl (Leaves) | 75.4 *** | 1098.7 *** | 14.4 *** |
| Cl (Roots) | 77.4 *** | 4313.3 *** | 54.2 *** |
| Chl a | Chl b | T Chl | ||||
|---|---|---|---|---|---|---|
| Level | Difference | p-Value | Difference | p-Value | Difference | p-Value |
| HS1 | 0.5405 | 0.0316 * | 0.6725 | 0.0014 * | 1.2135 | <0.0001 * |
| HS17 | 1.3205 | <0.0001 * | 0.7856 | 0.0001 * | 2.1069 | <0.0001 * |
| HC15 | −0.0661 | 0.9971 | 1.1289 | <0.0001 * | 1.1307 | 0.0001 * |
| Volk | −0.3983 | 0.1687 | 0.7577 | 0.0003 * | 0.3595 | 0.4782 |
| Czo | −0.5177 | 0.0425 * | 0.1340 | 0.9081 | −0.3837 | 0.4157 |
| E | A | gsw | ||||
|---|---|---|---|---|---|---|
| Level | Difference | p-Value | Difference | p-Value | Difference | p-Value |
| HS1 | 0.0668 | 0.9932 | −0.1012 | 0.9951 | 6.6938 | 0.4283 |
| HS17 | 0.3162 | 0.2103 | −0.0425 | 0.9999 | 13.1075 | 0.0194 * |
| HC15 | −0.1068 | 0.9498 | 0.8187 | 0.0139 * | −0.9031 | 0.9998 |
| Volk | −0.1750 | 0.7337 | 0.2856 | 0.7302 | 9.2594 | 0.1541 |
| Czo | −0.3443 | 0.15 | −1.6375 | <0.0001 * | −8.8038 | 0.1890 |
| Fv/Fo | Fv/Fm | |||
|---|---|---|---|---|
| Level | Difference | p-Value | Difference | p-Value |
| HS1 | −0.1255 | 0.8287 | −0.0045 | 0.9162 |
| HS17 | −0.3946 | 0.0230 * | −0.0169 | 0.0317 * |
| HC15 | −0.3598 | 0.0445 * | −0.0178 | 0.0217 * |
| Volk | −0.4769 | 0.0040 * | −0.0217 | 0.0033 * |
| Czo | −1.1125 | <0.0001 * | −0.0556 | <0.0001 * |
| MDA | TPC | Proline | ||||
|---|---|---|---|---|---|---|
| Level | Difference | p-Value | Difference | p-Value | Difference | p-Value |
| HS1 | 3.8623 | <0.0001 * | −34.086 | <0.0001 * | −17.1324 | <0.0001 * |
| HS17 | 3.2560 | <0.0001 * | −56.077 | <0.0001 * | 7.2909 | <0.0001 * |
| HC15 | −1.1645 | 0.1002 | 10.4936 | 0.1147 | 7.5692 | <0.0001 * |
| Volk | 3.8449 | <0.0001 * | −52.0436 | <0.0001 * | −17.1324 | <0.0001 * |
| Czo | 4.2042 | <0.0001 * | −90.3323 | <0.0001 * | −1.42 × 10−14 | 1 |
| Na (Leaves) | Na (Roots) | Cl (Leaves) | Cl (Roots) | |||||
|---|---|---|---|---|---|---|---|---|
| Level | Difference | p-Value | Difference | p-Value | Difference | p-Value | Difference | p-Value |
| HS1 | −0.0168 | 0.9986 | −0.1112 | <0.0001 * | 0.1606 | 0.1009 | 0.1343 | <0.0001 * |
| HS17 | −0.095 | 0.3548 | −0.0568 | 0.0315 * | −0.1112 | 0.3821 | 0.1562 | <0.0001 * |
| HC15 | −0.0318 | 0.9754 | 0.0325 | 0.3819 | 0.3512 | <0.0001 * | 0.0831 | <0.0001 * |
| Volk | −0.4025 | <0.0001 * | −0.0712 | 0.0043 * | −0.0568 | 0.8876 | 0.0768 | <0.0001 * |
| Czo | −0.1612 | 0.0318 * | −0.0031 | 0.9999 | −0.9125 | <0.0001 * | 0.2918 | <0.0001 * |
| Chl a | Chl b | T Chl | E | A | gsw | Fv/Fm | Fv/Fo | MDA | TPC | Proline | Na (leaves) | Na (Roots) | Cl (Leaves) | Cl (Roots) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chl a | 1 a | ||||||||||||||
| Chl b | 0.1893 | 1 | |||||||||||||
| T Chl | 0.9074 | 0.5799 | 1 | ||||||||||||
| E | 0.6051 | 0.4026 | 0.67 | 1 | |||||||||||
| A | 0.6772 | 0.3545 | 0.7082 | 0.6747 | 1 | ||||||||||
| gsw | 0.7034 | 0.2802 | 0.7008 | 0.7111 | 0.8427 | 1 | |||||||||
| Fv/Fm | 0.3127 | 0.1774 | 0.335 | 0.3778 | 0.4251 | 0.312 | 1 | ||||||||
| Fv/Fo | 0.2996 | 0.2104 | 0.3375 | 0.377 | 0.4168 | 0.2924 | 0.9638 | 1 | |||||||
| MDA | −0.621 | −0.313 | −0.648 | −0.534 | −0.65 | −0.54 | −0.28 | −0.276 | 1 | ||||||
| TPC | 0.5341 | 0.3256 | 0.5813 | 0.6103 | 0.7566 | 0.6171 | 0.492 | 0.4999 | −0.645 | 1 | |||||
| Proline | 0.1761 | −0.033 | 0.138 | 0.0318 | 0.122 | 0.0891 | −0.074 | −0.053 | −0.239 | 0.151 | 1 | ||||
| Na (leaves) | −0.66 | −0.291 | −0.669 | −0.626 | −0.756 | −0.825 | −0.173 | −0.169 | 0.6135 | −0.605 | −0.042 | 1 | |||
| Na (Roots) | −0.734 | −0.194 | −0.69 | −0.53 | −0.743 | −0.834 | −0.2 | −0.195 | 0.5245 | −0.588 | −0.035 | 0.8667 | 1 | ||
| Cl (Leaves) | −0.66 | −0.129 | −0.599 | −0.581 | −0.635 | −0.728 | −0.001 | −0.006 | 0.5708 | −0.486 | −0.076 | 0.8874 | 0.7721 | 1 | |
| Cl (Roots) | −0.733 | −0.266 | −0.717 | −0.621 | −0.841 | −0.841 | −0.287 | −0.29 | 0.725 | −0.76 | −0.048 | 0.8865 | 0.8596 | 0.8356 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmoud, L.M.; Dutt, M.; Vincent, C.I.; Grosser, J.W. Salinity-Induced Physiological Responses of Three Putative Salt Tolerant Citrus Rootstocks. Horticulturae 2020, 6, 90. https://doi.org/10.3390/horticulturae6040090
Mahmoud LM, Dutt M, Vincent CI, Grosser JW. Salinity-Induced Physiological Responses of Three Putative Salt Tolerant Citrus Rootstocks. Horticulturae. 2020; 6(4):90. https://doi.org/10.3390/horticulturae6040090
Chicago/Turabian StyleMahmoud, Lamiaa M., Manjul Dutt, Christopher I. Vincent, and Jude W. Grosser. 2020. "Salinity-Induced Physiological Responses of Three Putative Salt Tolerant Citrus Rootstocks" Horticulturae 6, no. 4: 90. https://doi.org/10.3390/horticulturae6040090
APA StyleMahmoud, L. M., Dutt, M., Vincent, C. I., & Grosser, J. W. (2020). Salinity-Induced Physiological Responses of Three Putative Salt Tolerant Citrus Rootstocks. Horticulturae, 6(4), 90. https://doi.org/10.3390/horticulturae6040090