
Effect of Cold Stress on Growth, Physiological Characteristics, and Calvin-Cycle-Related Gene Expression of Grafted Watermelon Seedlings of Different Gourd Rootstocks

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Chlorophyll Content and Plant Growth Measurements
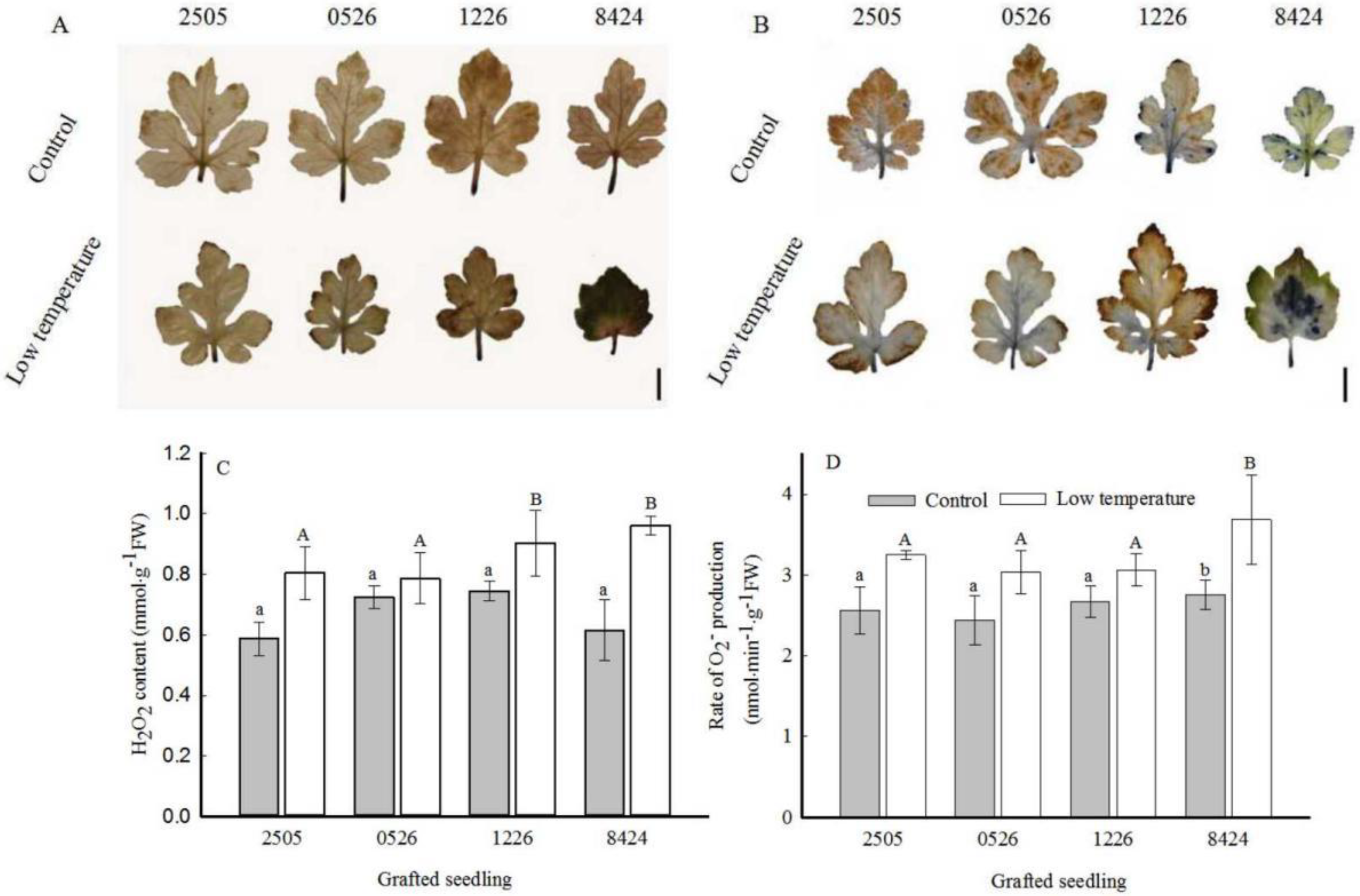
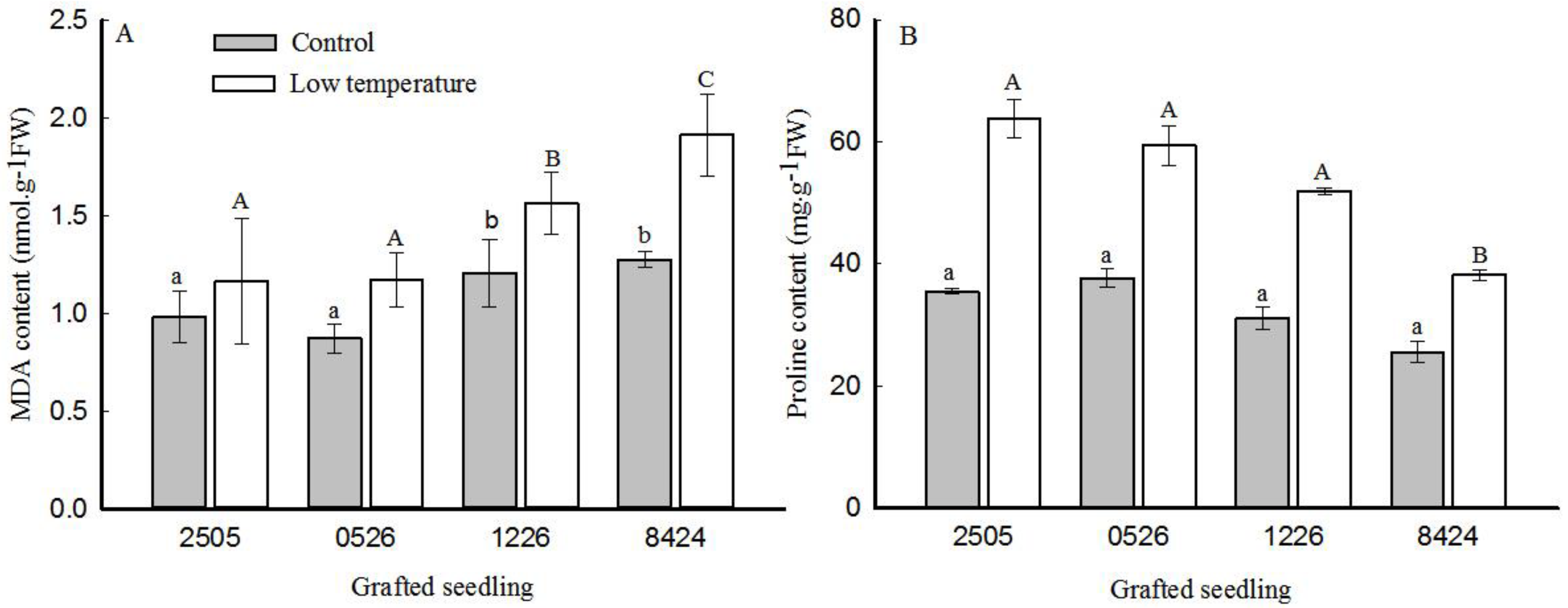
2.3. Measurements of ROS, MDA, and Proline Content
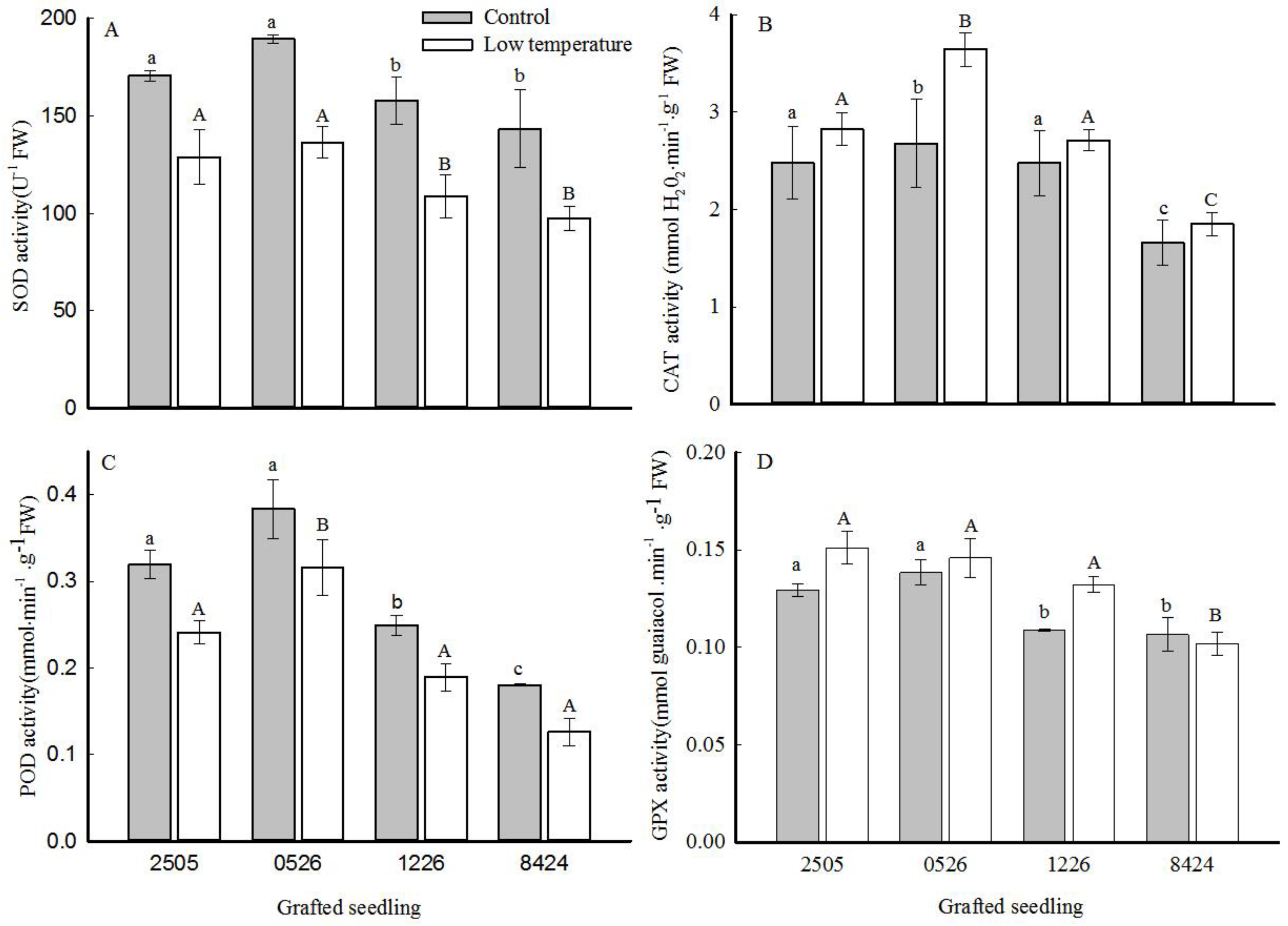
2.4. Antioxidant Enzyme Analysis
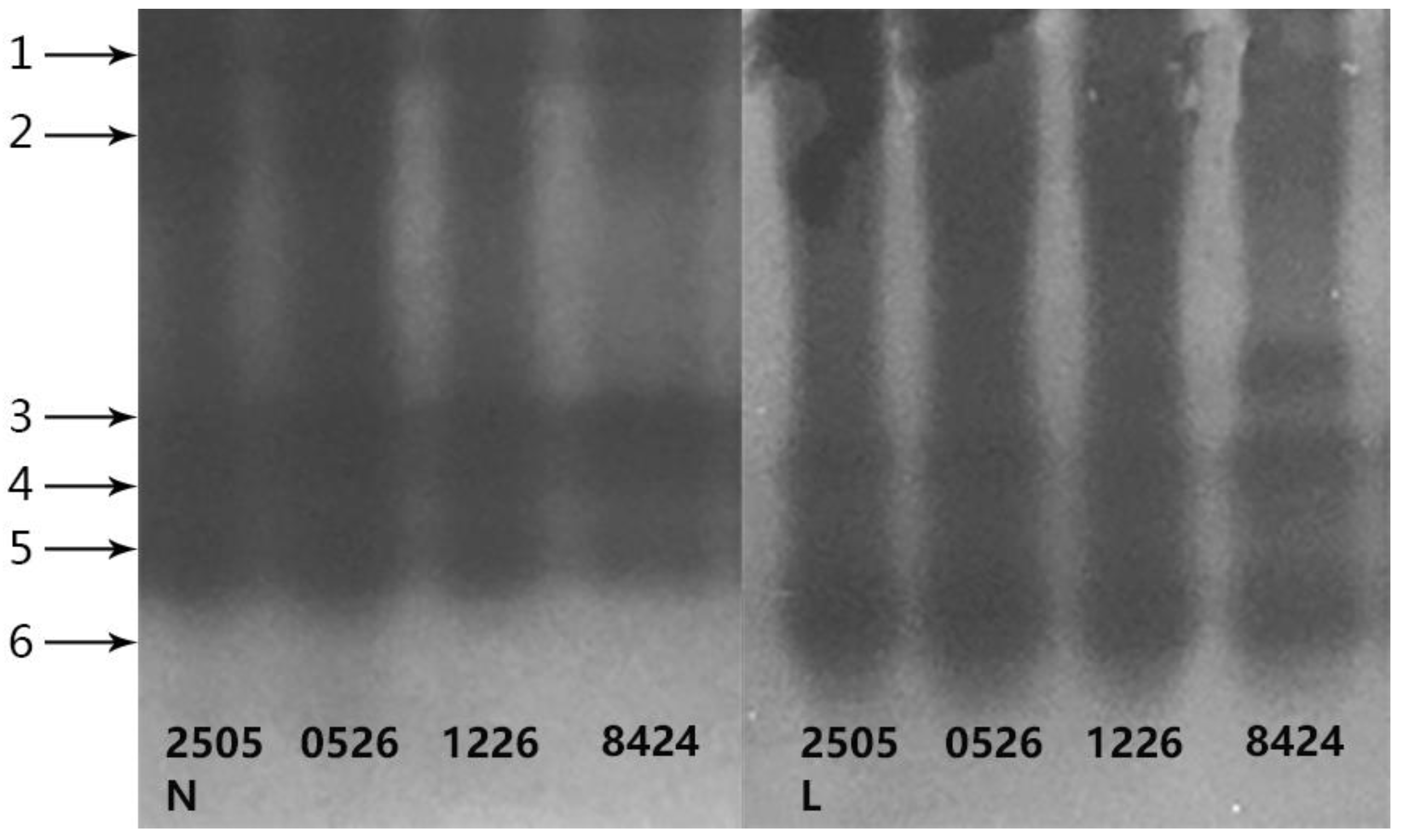
2.5. Gene Expression Analysis
2.6. Statistical Methods
3. Results
3.1. Growth Parameters
3.2. Chlorophyll Content
3.3. Oxidative Stress Evaluation
3.4. Lipid Peroxidation and Proline Accumulation
3.5. Antioxidant Enzyme Activity
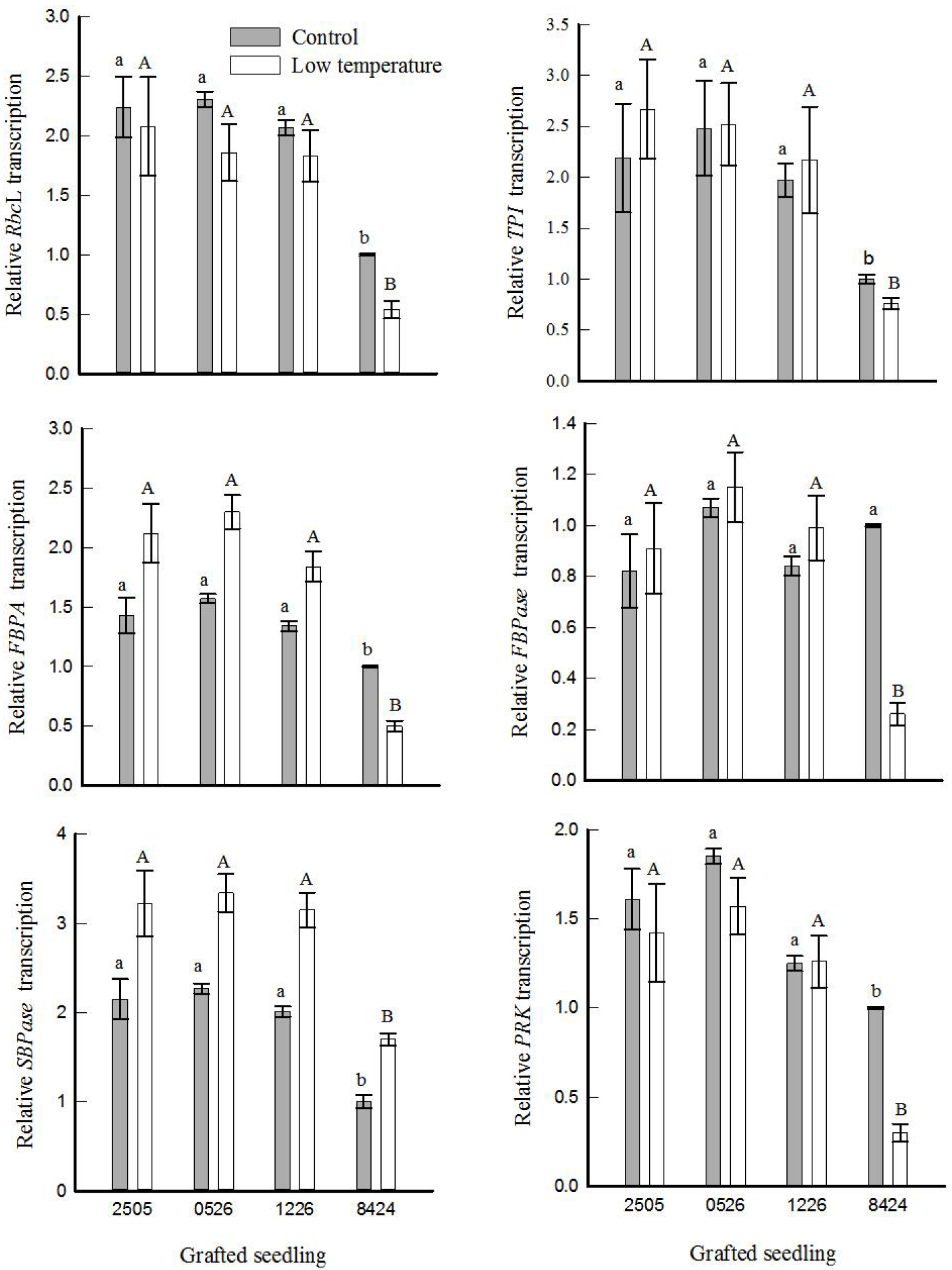
3.6. Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noh, J.; Kim, J.M.; Sheikh, S.; Lee, S.G.; Lim, J.H.; Seong, M.H.; Jung, G.T. Effect of heat treatment around the fruit set region on growth and yield of watermelon [Citrullus lanatus (Thunb.) Matsum. and Nakai]. Physiol. Mol. Biol. Plants 2013, 19, 509–514. [Google Scholar] [CrossRef] [Green Version]
- Hao, J.; Gu, F.; Zhu, J.; Lu, S.; Liu, Y.; Li, Y.; Chen, W.; Wang, L.; Fan, S.; Xian, C.J. Low night temperature affects the phloem ultrastructure of lateral branches and Raffinose Family Oligosaccharide (RFO) accumulation in RFO-Transporting plant Melon (Cucumismelo, L.) during fruit expansion. PLoS ONE 2016, 11, e0160909. [Google Scholar] [CrossRef] [Green Version]
- Machado, D.F.S.P.; Ribeiro, R.V.; da Silveira, J.A.G.; Filho, J.R.M.; Machado, E.C. Rootstocks induce contrasting photosyn-thetic responses of orange plants to low night temperature without affecting the antioxidant metabolism. Theor. Exp. Plant Phys. 2013, 25, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Niu, M.; Xie, J.; Sun, J.; Huang, Y.; Kong, Q.; Nawaz, M.A.; Bie, Z. A shoot based Na+ tolerance mechanism observed in pumpkin—An important consideration for screening salt tolerant rootstocks. Sci. Hortic. 2017, 218, 38–47. [Google Scholar] [CrossRef]
- Ntatsi, G.; Savvas, D.; Kläring, H.-P.; Schwarz, D. Growth, Yield, and metabolic responses of temperature-stressed tomato to grafting onto rootstocks differing in cold tolerance. J. Am. Soc. Hortic. Sci. 2014, 139, 230–243. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.M.; Chai, F.M.; Zhu, Z.F.; Elias, G.K.; Xin, H.P.; Liang, Z.C.; Li, S.H. The inheritance of cold tolerance in seven in-terspecifific grape populations. Sci. Hortic. 2020, 266, 109260. [Google Scholar] [CrossRef]
- Xu, J.H.; Zhang, M.; Liu, G.; Yang, X.P.; Hou, X.L. Comparative transcriptome profifiling of chilling stress responsiveness in grafted watermelon seedlings. Plant Physiol. Biochem. 2016, 109, 561e570. [Google Scholar] [CrossRef]
- Kawaide, T. Utilization of rootstocks in cucurbits production in Japan. Jpn. Agric. Res. Q. 1985, 18, 284–289. [Google Scholar]
- Lee, J.-M.; Kubota, C.; Tsao, S.J.; Bie, Z.; Echevarria, P.H.; Morra, L.; Oda, M. Current status of vegetable grafting: Diffusion, grafting techniques, automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Yarşi, G.; Altuntas, O.; Sivaci, A.; Dasgan, H.Y. Effects of salinity stress on plant growth and mineral composition of grafted and un-grafted Galia C8 melon cultivar. Pak. J. Bot. 2017, 49, 819–822. [Google Scholar]
- Xu, Y.; Guo, S.-R.; Li, H.; Sun, H.-Z.; Na, L.; Shu, S.; Sun, J. Resistance of cucumber grafting rootstock pumpkin cultivars to chilling and salinity stresses. Hort. Sci. Technol. 2017, 35, 220–231. [Google Scholar] [CrossRef]
- Lo’ay, A.A.; Abo EL-Ezz, S.F. Performance of ‘Flame seedless’ grapevines grown on different rootstocks in response to soil salinity stress. Sci. Hortic. 2021, 275, 109704. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, S.; Wei, M.; Gong, B.; Shi, Q. Effect of different rootstocks on the salt stress tolerance in watermelon seedlings. Hortic. Plant J. 2018, 4, 239–249. [Google Scholar] [CrossRef]
- Li, H.; Liu, S.S.; Yi, C.Y.; Wang, F.; Zhou, J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Hydrogen peroxide mediates abscisic acid-induced HSP 70 accumulation and heat tolerance in grafted cucumber plants. Plant Cell Environ. 2014, 37, 2768–2780. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, E.; Romero, L.; Ruiz, J. Accumulation of free polyamines enhances the antioxidant response in fruits of grafted tomato plants under water stress. J. Plant Physiol. 2016, 190, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.Q.; Chen, C.; Nawaz, M.A.; Jiao, Y.Y.; Zheng, Z.H.; Shi, X.F.; Xie, W.T.; Yu, Y.G.; Guo, J.; Zhu, S.H.; et al. Using rootstock to increase watermelon fruit yield and quality at low potassium supply: A comprehensive analysis from agronomic, physiological and transcriptional perspective. Sci. Hortic. 2018, 241, 144–151. [Google Scholar] [CrossRef]
- Ramón, G.-M.; Yaiza, G.P.; Mary-Rus, M.-C.; Salvador, L.-G.; Ángeles, C. Suitable rootstocks can alleviate the effects of heat stress on pepper plants. Sci. Hortic. 2021, 290, 110529. [Google Scholar]
- Chen, S.; Yi, L.; Korpelainen, H.; Yu, F.; Liu, M. Roots play a key role in drought-tolerance of poplars as suggested by reciprocal grafting between male and female clones. Plant Physiol. Biochem. 2020, 153, 81–91. [Google Scholar] [CrossRef]
- Kumar, P.; Rouphael, Y.; Cardarelli, M.; Colla, G. Vegetable grafting as a tool to improve drought resistance and water use efficiency. Front. Plant Sci. 2017, 8, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.J.; Liang, G.Y.; Liu, X.J.; Liu, D.C.; Fang, C. Proteomic study on grafted and non-grafted cucumber (Cucumis sativus L.). Acta. Hortic. Sin. 2009, 36, 1147–1152. [Google Scholar]
- Yang, Y.; Wang, L.; Tian, J.; Li, J.; Sun, J.; He, L.; Guo, S.; Tezuka, T. Proteomic study participating the enhancement of growth and salt tolerance of bottle gourd rootstock-grafted watermelon seedlings. Plant Physiol. Biochem. 2012, 58, 54–65. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, L.; Wang, L.; Guo, S. Bottle gourd rootstock-grafting promotes photosynthesis by regulating the stomata and non-stomata performances in leaves of watermelon seedlings under NaCl stress. J. Plant Physiol. 2015, 186, 50–58. [Google Scholar] [CrossRef]
- Han, Q.; Guo, Q.; Korpelainen, H.; Niinemets, Ü.; Li, C. Rootstock determines the drought resistance of poplar grafting com-binations. Tree Physiol. 2019, 39, 1855–1866. [Google Scholar] [CrossRef]
- Usanmaz, S.; Abak, K. Plant growth and yield of cucumber plants grafted on different commercial and local rootstocks grown under salinity stress. Saudi J. Biol. Sci. 2019, 26, 1134–1139. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-M. Cultivation of grafted vegetables, I. current status, grafting methods, and benefits. HortScience 1994, 29, 235–239. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Airaki, M.; Leterrier, M.; Mateos, R.M.; Valderrama, R.; Chaki, M.; Barroso, J.B.; Delrío, L.A.; Palma, J.M.; Corpas, F.J. Metabolism of reactive oxygen species and reactive nitrogen species in pepper (Capsicum annuum L.) plants under low temperature stress. Plant Cell Environ. 2012, 35, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Ding, W.; Zhu, S.; Jiang, D. Salt-induced difference between Glycine cyrtoloba and G. max in anti-oxidative ability and K+ vs. Na+ selective accumulation. Crop. J. 2016, 4, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Huang, Z.A.; Jiang, D.A.; Yang, Y.; Sun, J.W.; Jin, S.H. Effects of nitrogen deficiency on gas exchange, chlorophyll fluorescence, and antioxidant enzymes in leaves of rice plants. Photosynthetica 2004, 42, 357–364. [Google Scholar] [CrossRef]
- Woodbury, W.; Spencer, A.; Stahmann, M. An improved procedure using ferricyanide for detecting catalase isozymes. Anal. Biochem. 1971, 44, 301–305. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Tao, M.-Q.; Jahan, M.S.; Hou, K.; Shu, S.; Wang, Y.; Sun, J.; Guo, S.-R. Bitter melon (Momordica charantia L.) rootstock improves the heat tolerance of cucumber by regulating photosynthetic and antioxidant defense pathways. Plants 2020, 9, 692. [Google Scholar] [CrossRef]
- Soares, C.; Carvalho, M.; Azevedo, R.A.; Fidalgo, F. Plants facing oxidative challenges—A little help from the antioxidant networks. Environ. Exp. Bot. 2019, 161, 4–25. [Google Scholar] [CrossRef]
- Lo’ay, A.A.; Doaa, M.H. The potential of vine rootstocks impacts on ‘Flame Seedless’ bunches behavior under cold storage and antioxidant enzyme activity performance. Sci. Hortic. 2020, 260, 108844. [Google Scholar] [CrossRef]
- Wakeel, A.; Xu, M.; Gan, Y. Chromium-induced reactive oxygen species accumulation by altering the enzymatic antioxidant system and associated cytotoxic, genotoxic, ultrastructural, and photosynthetic changes in plants. Int. J. Mol. Sci. 2020, 21, 728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- Petrić, M.; Jevremović, S.; Trifunović, M.; Tadić, V.; Milošević, S.; Dragićević, M.; Subotić, A. The effect of low temperature and GA3 treatments on dormancy breaking and activity of antioxidant enzymes in Fritillaria meleagris bulblets cultured in vitro. Acta Physiol. Plant. 2013, 35, 3223–3236. [Google Scholar] [CrossRef]
- Xu, S.-C.; Li, Y.-P.; Hu, J.; Guan, Y.-J.; Ma, W.-G.; Zheng, Y.-Y.; Zhu, S.-J. Responses of Antioxidant enzymes to chilling stress in tobacco seedlings. Agric. Sci. China 2010, 9, 1594–1601. [Google Scholar] [CrossRef]
- Song, G.; Hou, W.; Wang, Q.; Wang, J.; Jin, X. Effect of low temperature on eutrophicated waterbody restoration by Spirodela polyrhiza. Bioresour. Technol. 2006, 97, 1865–1869. [Google Scholar] [CrossRef] [PubMed]
- Whitney, S.M.; Houtz, R.L.; Alonso, H. Advancing our understanding and capacity to engineer nature’s CO2-sequestering enzyme, Rubisco. Plant Physiol. 2011, 155, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| rbcL | TCTTGGCAGCATTCCGAGTAA | TCGCAAATCCTCCAGACGTAG |
| TPI | GAAATTCTTCGTCGGTGGC | GAACCCAACAATTCCGTGCTG |
| FBPA | GTTGGTCCCTATTGTGGAGCC | CCTTGTAAACCGCAGC |
| FBPase | TCACAGCCCTCGAATTTA | CTTCGGAAACAAGGATACAAG |
| SBPase | TCGAGGCCTTGAGATACTCAC | GCCATCGCTGCTGTAACC |
| PRK | GGGCTGAGAAGATTACC | GAAGGATCTACAATCTCATGG |
| Treatment | Seedlings | Plant Height (cm) | Stem Diameter (mm) | Dry Weight (g) | Number of Leaves |
|---|---|---|---|---|---|
| Normal | 2505 | 16.69 ± 0.87 c | 3.32 ± 0.35 c | 1.04 ± 0.11 c | 9.11 ± 0.51 c |
| 0526 | 17.08 ± 0.28 c | 2.90 ± 0.27 ab | 1.15 ± 0.08 c | 8.53 ± 0.60 c | |
| 1226 | 15.20 ± 1.03 b | 2.67 ± 0.16 b | 0.92 ± 0.07 b | 7.18 ± 0.57 b | |
| 8424 | 8.47 ± 0.64 a | 2.13 ± 0.12 a | 0.51 ± 0.05 a | 4.07 ± 0.65 a | |
| Cold | 2505 | 15.06 ± 0.99 c | 3.04 ± 0.27 b | 0.92 ± 0.06 c | 7.62 ± 0.43 c |
| 0526 | 15.82 ± 0.43 d | 2.47 ± 0.19 b | 0.81 ± 0.05 d | 7.02 ± 0.68 b | |
| 1226 | 13.03 ± 0.82 b | 2.18 ± 0.14 b | 1.06 ± 0.10 b | 5.57 ± 0.48 b | |
| 8424 | 7.08 ± 0.38 a | 1.81 ± 0.08 a | 0.48 ± 0.04 a | 3.82 ± 0.31 a | |
| Two way ANOVA | |||||
| Fs | 10.05 *** | 5.33 ** | 6.27 ** | 3.97 ** | |
| Fc | 6.85 *** | 17.19 * | 6.43 ** | 10.58 ** | |
| Fs×c | 3.27 *** | 6.84 * | 9.51 * | 2.20 ** |
| Treatment | Seedlings | Chlorophyll (mg g−1 FW) | Chl a/b |
|---|---|---|---|
| Normal | 2505 | 1.84 ± 0.04 b | 3.46 ± 0.10 a |
| 0526 | 1.86 ± 0.02 b | 3.41 ± 0.03 a | |
| 1226 | 1.71 ± 0.07 b | 3.39 ± 0.08 a | |
| 8424 | 1.46 ± 0.13 a | 3.29 ± 0.10 a | |
| Cold | 2505 | 1.36 ± 0.03 c | 2.44 ± 0.03 c |
| 0526 | 1.39 ± 0.03 b | 2.35 ± 0.02 c | |
| 1226 | 1.13 ± 0.07 ab | 2.28 ± 0.04 b | |
| 8424 | 0.92 ± 0.09 a | 2.03 ± 0.02 a | |
| Two-way ANOVA | |||
| Fs | 10.15 ** | 8.14 ** | |
| Fc | 13.24 ** | 6.82 ** | |
| Fs×c | 12.49 * | 9.37 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, K.; Sun, J.; Li, Q.; Li, X.; Jin, S. Effect of Cold Stress on Growth, Physiological Characteristics, and Calvin-Cycle-Related Gene Expression of Grafted Watermelon Seedlings of Different Gourd Rootstocks. Horticulturae 2021, 7, 391. https://doi.org/10.3390/horticulturae7100391
Lu K, Sun J, Li Q, Li X, Jin S. Effect of Cold Stress on Growth, Physiological Characteristics, and Calvin-Cycle-Related Gene Expression of Grafted Watermelon Seedlings of Different Gourd Rootstocks. Horticulturae. 2021; 7(10):391. https://doi.org/10.3390/horticulturae7100391
Chicago/Turabian StyleLu, Kaixing, Jiutong Sun, Qiuping Li, Xueqin Li, and Songheng Jin. 2021. "Effect of Cold Stress on Growth, Physiological Characteristics, and Calvin-Cycle-Related Gene Expression of Grafted Watermelon Seedlings of Different Gourd Rootstocks" Horticulturae 7, no. 10: 391. https://doi.org/10.3390/horticulturae7100391
APA StyleLu, K., Sun, J., Li, Q., Li, X., & Jin, S. (2021). Effect of Cold Stress on Growth, Physiological Characteristics, and Calvin-Cycle-Related Gene Expression of Grafted Watermelon Seedlings of Different Gourd Rootstocks. Horticulturae, 7(10), 391. https://doi.org/10.3390/horticulturae7100391