
The Combined Effect of Pseudomonas stutzeri and Biochar on the Growth Dynamics and Tolerance of Lettuce Plants (Lactuca sativa) to Cadmium Stress



Abstract
:1. Introduction
2. Materials and Methods
2.1. Pseudomonas Strains and Growth Conditions
2.2. Assessment of Different Pseudomonas Strains for Cd Tolerance
2.3. Biosorption of Cd by Different Pseudomonas Strains
2.4. Biochar Characterization
2.5. Pot Trial
2.6. Trait Measurements
2.6.1. Growth Parameters
2.6.2. Physiological Characteristics
2.6.3. Photosynthetic Pigments
2.6.4. Total Soluble Sugars (TSSs)
2.6.5. Proline
2.6.6. Activity of Antioxidant Enzymes
2.7. Soil Microbiological Activity
2.7.1. Microbial Biomass Carbon
2.7.2. CO2 Evolution
2.7.3. Dehydrogenase Activity (DHA)
2.8. Determination of Cd in Plant Organs
2.9. Bioconcentration and Translocation Factors
2.10. Statistical Analysis
3. Results
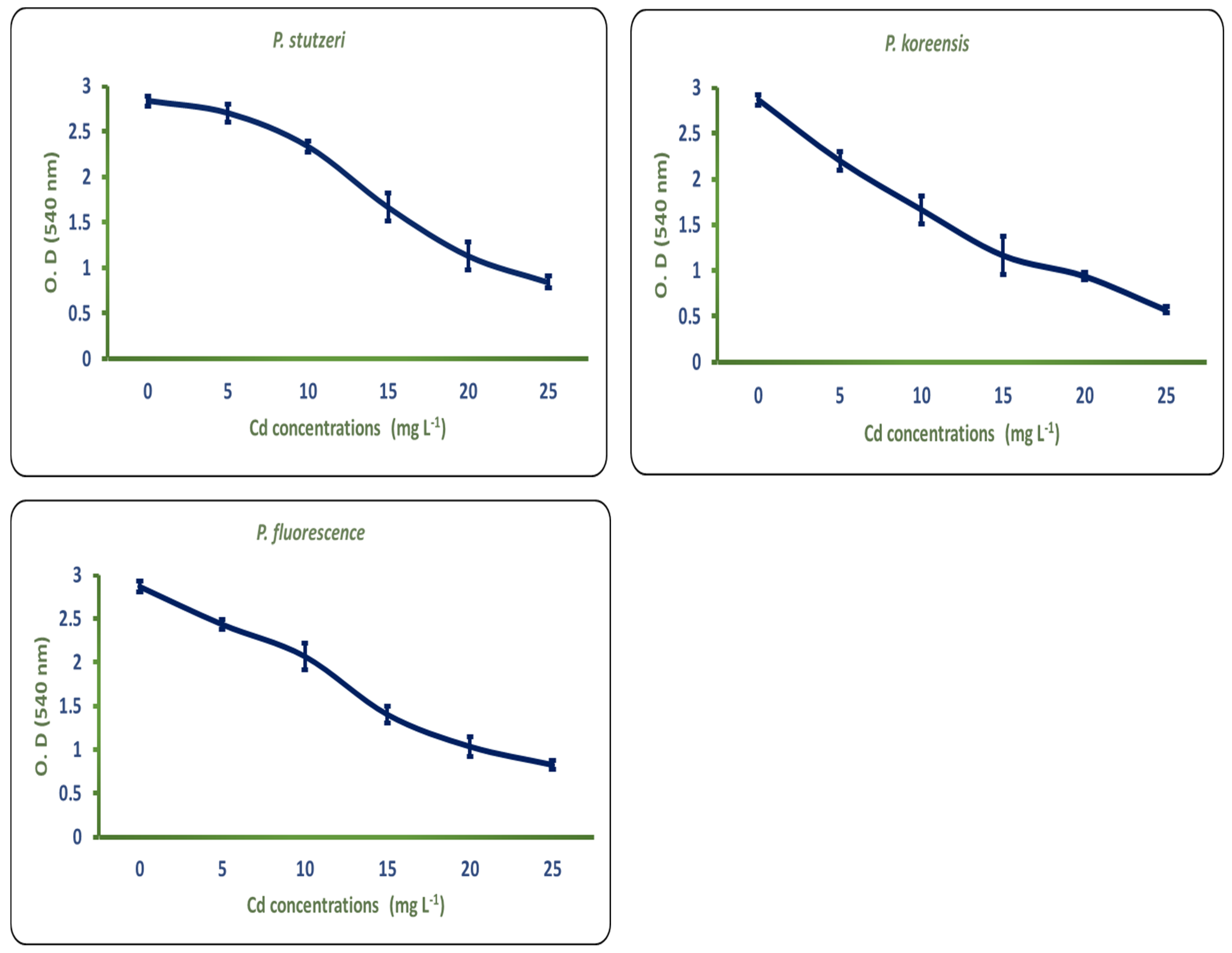
3.1. Assessment of Different Pseudomonas Strains for Cd Tolerance
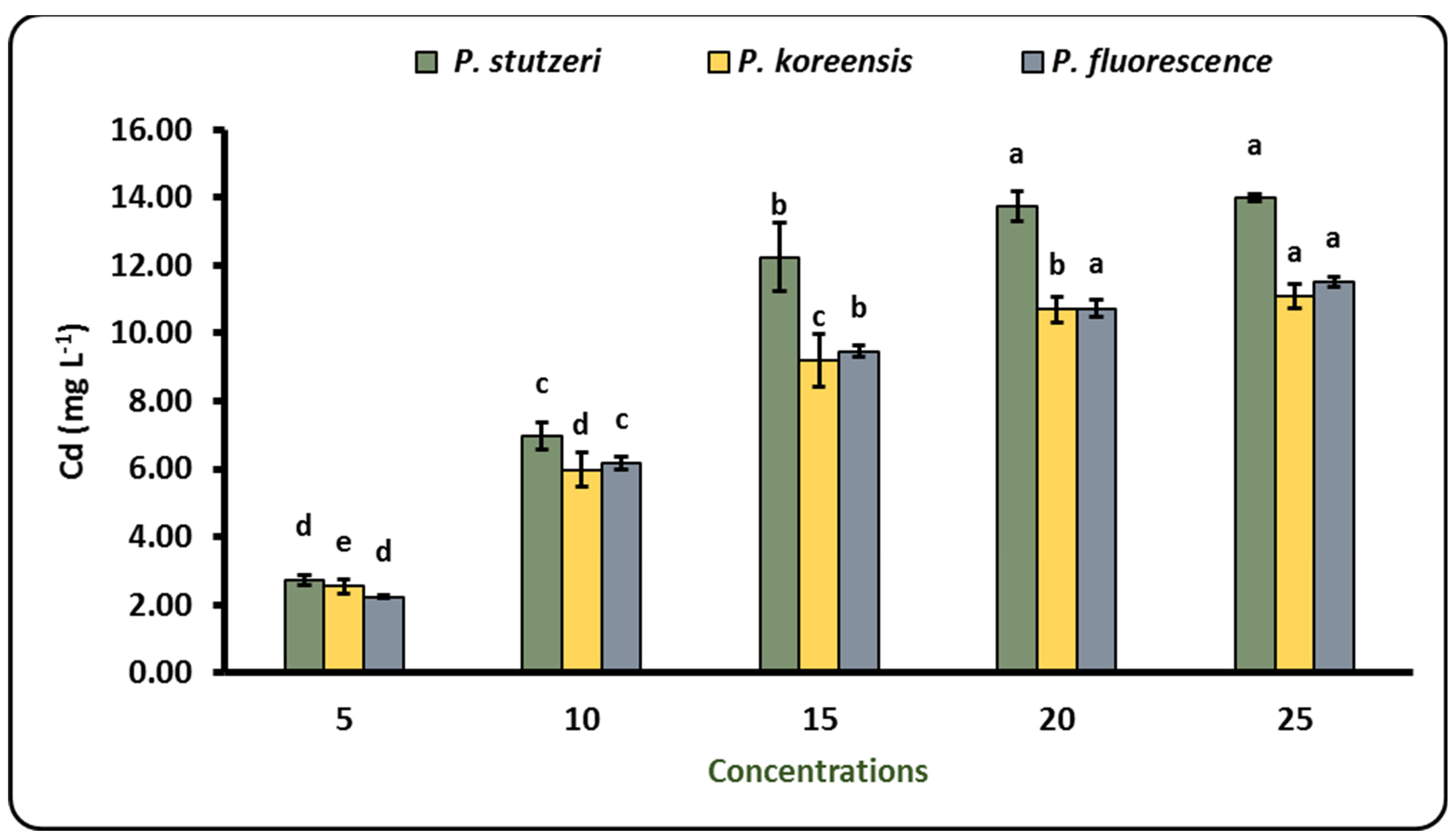
3.2. Biosorption of Cd by Different Pseudomonas Strains
3.3. Pot Trial
3.3.1. Growth Parameters
3.3.2. Physiological Characteristics
3.3.3. Activity of Antioxidant Enzymes
3.3.4. Soil Microbiological Activity
3.3.5. Cadmium Content
4. Discussion
4.1. Differential Tolerance Levels of Tested Pseudomonas Strains to Gradients’ Cadmium Concentrations
4.2. Cadmium Biosorption by Different Pseudomonas Strains
4.3. Pot Experiment
4.3.1. Plant Growth Analysis
4.3.2. Physiological Traits
4.3.3. Antioxidant Enzymes Activities
4.3.4. Microbiological Activity in the Soil
4.3.5. Cadmium Content of Lettuce Plants
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arshad, M.; Khan, A.H.; Hussain, I.; Anees, M.; Iqbal, M.; Soja, G.; Linde, C.; Yousaf, S. The reduction of chromium (VI) phytotoxicity and phytoavailability to wheat (Triticum aestivum L.) using biochar and bacteria. Appl. Soil Ecol. 2017, 114, 90–98. [Google Scholar] [CrossRef]
- Iqbal, A.; Mushtaq, M.; Khan, A.; Nawaz, I.; Yousaf, S.; Iqbal, M. Influence of Pseudomonas japonica and organic amendments on the growth and metal tolerance of Celosia argentea L. Environ. Sci. Pollut. Res. 2020, 27, 24671–24685. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Yang, L. A review of heavy metal contaminations in urban soils, urban road dusts and agricultural soils from China. Microchem. J. 2010, 94, 99–107. [Google Scholar] [CrossRef]
- Mazhar, R.; Ilyas, N.; Arshad, M.; Khalid, A.; Hussain, M. Isolation of heavy metal-tolerant PGPR strains and amelioration of chromium effect in wheat in combination with biochar. Iran. J. Sci. Technol. Trans. A Sci. 2020, 44, 1–12. [Google Scholar] [CrossRef]
- Boparai, H.K.; Joseph, M.; O’Carroll, D.M. Kinetics and thermodynamics of cadmium ions removal by adsorption onto nano zerovalent iron particles. J. Hazard Mater. 2011, 186, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Segura, E.; Solache-Rio, M.; Colin-Cruz, A.; Fall, C. Adsorption of cadmium by Na and Fe modified zeolitic tuffs and carbonaceous material from pyrolyzed sewage sludge. J. Env. Manag. 2012, 97, 6–13. [Google Scholar] [CrossRef]
- Khan, M.A.; Khan, S.; Khan, A.; Alam, M. Soil contamination with cadmium, consequences and remediation using organic amendments. Sci. Total Env. 2017, 601, 1591–1605. 8. [Google Scholar] [CrossRef] [PubMed]
- Balkaya, N.; Cesur, H. Adsorption of cadmium from aqueous solution by phosphogypsum. Chem. Eng. J. 2008, 140, 247–254. [Google Scholar] [CrossRef]
- ASTDR. Toxicological Profile for Cadmium; Prepared for US Department of Health and Human Services, Public Health Service, Agency for Toxic Substances and Disease Registry (ASTDR); ASTDR: Atlanta, GA, USA, 1999.
- Takahashi, R.; Ishimaru, Y.; Nakanishi, H.; Nishizawa, N.K. Role of the iron transporter OsNRAMP1 in cadmium uptake and accumulation in rice. Plant Signal. Behav. 2011, 6, 1813–1816. [Google Scholar] [CrossRef] [Green Version]
- Zorrig, W.; Abdelly, C.; Berthomieu, P. The phylogenetic tree gathering the plant Zn/Cd/Pb/Co P1B-ATPases appears to be structured according to the botanical families. Comptes Rendus Biol. 2011, 334, 863–871. [Google Scholar] [CrossRef]
- Zorrig, W.; El Khouni, A.; Ghnaya, T.; Davidian, J.C.; Abdelly, C.; Berthomieu, P. Lettuce (Lactuca sativa): A species with a high capacity for cadmium (Cd) accumulation and growth stimulation in the presence of low Cd concentrations. J. Hortic. Sci. Biotechnol. 2013, 88, 783–789. [Google Scholar] [CrossRef]
- Benavides, M.P.; Gallego, S.M.; Tomaro, M.L. Cadmium toxicity in plants. Braz J. Plant Physiol. 2005, 17, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Akoto, O.; Addo, D.; Baidoo, E.; Agyapong, E.A.; Apau, J.; Fei-Baffoe, B. Heavy metal accumulation in untreated wastewater-irrigated soil and lettuce (Lactuca sativa). Environ. Earth Sci. 2015, 74, 6193–6198. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Cervantes, C.; Silver, S. Chromium reduction in Pseudomonas putida. Appl. Environ. Microbiol. 1990, 56, 2268–2270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itusha, A.W.; Jabez, O.; Mohanasrinivasan, V. Enhanced uptake of Cd by biofilm forming Cd resistant plant growth promoting bacteria bioaugmented to the rhizosphere of Vetiveria zizanioides. Int. J. Phytoremediat. 2019, 21, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Zand, A.D.; Alireza, M.T.; Azar, V.H. Application of titanium dioxide nanoparticles to promote phytoremediation of Cd-polluted soil: Contribution of PGPR inoculation. Bioremediat. J. 2020, 24, 171–189. [Google Scholar] [CrossRef]
- Novak, J.M.; Busscher, W.J.; Laird, D.L.; Ahmedna, M.; Watts, D.W.; Niandou, M.A.S. Impact of biochar amendment on fertility of a Southeastern Coastal Plain soil. Soil Sci. 2009, 174, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, L.; Lesueur, D. Challenges of formulation and quality of biofertilizers for successful inoculation. Appl. Microbiol. Biotechnol. 2013, 97, 8859–8873. [Google Scholar] [CrossRef] [PubMed]
- Beesley, L.; Moreno-Jimenez, E.; Gomez-Eyles, J.L. Effect of biochar and greenwaste compost amendments on mobility, bioavailability and toxicity of inorganic and organic contaminants in a multielement polluted soil. Env. Pollut. 2010, 158, 2282–2287. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota–a review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Ventura, M.; Sorrenti, G.; Panzacchi, P.; George, E.; Tonon, G. Biochar reduces short-term nitrate leaching from a horizon in an apple orchard. J. Env. Qual. 2013, 42, 76–82. [Google Scholar] [CrossRef]
- Major, J.; Lehmann, J.; Rondon, M.; Goodale, C. Fate of soil-applied black carbon: Downward migration, leaching and soil respiration. Glob. Chang. Biol. 2010, 16, 1366–1379. [Google Scholar] [CrossRef]
- Lehmann, J.; da Silva, J.P.; Steiner, C.; Nehls, T.; Zech, W.; Glaser, B. Nutrient availability and leaching in an archaeological Anthrosol and a Ferralsol of the Central Amazon basin: Fertilizer, manure and charcoal amendments. Plant Soil 2003, 249, 343–357. [Google Scholar] [CrossRef]
- Lehmann, J.; Joseph, S. Biochar for environmental management: An introduction. In Biochar for Environmental Management: Science, Technology and Implementation, 2nd ed.; Lehmann, J., Joseph, S., Eds.; Earthscan from Routledge: London, UK, 2015; pp. 1–1214. [Google Scholar]
- Mondal, S.C.; Sarma, B.; Farooq, M.; Nath, D.J.; Gogoi, N. Cadmium bioavailability in acidic soils under bean cultivation: Role of soil additives. Int. J. Environ. Sci. Technol. 2020, 17, 153–160. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Imran, M.; Naveed, M.; Khan, M.Y.; Ahmad, M.; Zahir, Z.A.; Crowley, D.E. Synergistic use of biochar, compost and plant growth-promoting rhizobacteria for enhancing cucumber growth under water deficit conditions. J. Sci Food Agric. 2017, 97, 5139–5145. [Google Scholar] [CrossRef]
- Singleton, P.; Sainsbury, D. Dictionary of Microbiology and Molecular Biology; Wiley: New York, NY, USA, 1987; Volume 721. [Google Scholar]
- Cho, J.S.; Hur, J.S.; Kang, B.H.; Kim, P.J.; Sohn, B.K.; Lee, H.J.; Jung, Y.K.; Heo, J.S. Biosorption of copper by immobilized biomass of Pseudomonas stutzer. J. Microbiol. Biotechnol. 2001, 11, 964–972. [Google Scholar]
- Singh, P.B.; Hatton, J.B.; Singh, B.; Cowie, L.A.; Kathuria, A. Influence of biochars on nitrous oxide emission and nitrogen leaching from two contrasting soils. J. Env. Qual. 2010, 39, 1224–1235. [Google Scholar] [CrossRef]
- Deb, S.; Ahmed, S.F.; Basu, M. Metal accumulation in cell wall: A possible mechanism of cadmium resistance by Pseudomonas stutzeri. Bull. Environ. Contamin. Toxicol. 2013, 90, 323–328. [Google Scholar] [CrossRef]
- King, E.; Ward, W.; Ramy, D. Two simple media for the demonstration of pyocyanin and fluorescence. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar]
- Massadeh, A.M.; Al-Momani, F.A.; Haddad, H.I. Removal of lead and cadmium by halophilic bacteria isolated from the Dead Sea shore, Jordan. Biol. Trace Elem. Res. 2005, 108, 259–269. [Google Scholar] [CrossRef]
- Omara, A.A. Study on Relationship of N2- Fixing and Phosphate–Dissolving Microorganisms with Soil Heavy Metals Pollution. Master’s Thesis, Faculty of Agriculture Mansoura University of Egypt, Mansoura, Egypt, 2009. [Google Scholar]
- Skradleta, V.; Gaudinova, A.; Necova, M.; Hydrakova, A. Behaviour of nodulated Pisum sativum L. under short term nitrate stress conditions. Biol. Plant 1984, 26, 364. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Academic Press: San Diego, CA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Hendrix, D.L. Rapid extraction and analysis of nonstructural carbohydrates in plant tissues. Crop. Sci. 1993, 33, 1306–1311. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 5, 867–880. [Google Scholar]
- Rao, M.V.; Paliyath, C.; Ormrod, D.P.; Murr, D.P.; Watkins, C.B. Influence of salicylic acid on H2O2 production, oxidative stress and H2O2-metabolizing enzymes: Salicylic acidmediated oxidative damage requires H2O2. Plant Physiol. 1997, 115, 137–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Joergensen, R.G.; Pommerening, B. Measurement of soil microbial biomass C by fumigation-extraction an automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Hu, X.; Cao, C.; Zhiping, A. Size and activity of the soil microbial biomass and chemical and biological properties. Commun. Soil Sci. Plant Anal. 2007, 40, 2072–2086. [Google Scholar]
- Coleman, D.C.; Anderson, R.V.; Cole, C.V. Tropic interactions in soils as they affect energy and nutrient dynamics. IV. Flows of metabolic and biomass carbon. Microb. Ecol. 1978, 4, 373–380. [Google Scholar] [CrossRef]
- Chander, K.; Brookes, C. Is the dehydrogenase assay invalid to estimate microbial activity in copper-contaminated soils? Soil Biol. Biochem. 1991, 23, 909–915. [Google Scholar] [CrossRef]
- Humphries, E.C. Mineral components and ash analysis. Mod. Methods. Plant. Anal. 1956, 1, 468–502. [Google Scholar]
- Baker, A.J. Accumulators and excluders-strategies in the response of plants to heavy metals. J. Plant Nutr. 1981, 3, 643–654. [Google Scholar] [CrossRef]
- Embrandiri, A.; Rupani, P.; Shahadat, M.; Singh, R.; Ismail, S.; Ibrahim, M.; Kadir, A. The phytoextraction potential of selected vegetable plants from soil amended with oil palm decanter cake. Int. J. Recycl. Org. Waste Agric. 2017, 6, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Env. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Xie, Y.; Fan, J.; Zhu, W.; Amombo, E.; Lou, Y.; Chen, L.; Fu, J. Effect of heavy metals pollution on soil microbial diversity and bermudagrass genetic variation. Front. Plant Sci. 2016, 7, 755. [Google Scholar] [CrossRef] [PubMed]
- Haritha, A.; Sagar, K.P.; Tiwari, A.; Kiranmayi, P.; Rodrigue, A.; Mohan, P.M.; Singh, S.S. MrdH, a novel metal resistance determinant of Pseudomonas putida KT2440, is flanked by metalinducible mobile genetic elements. J. Bacteriol. 2009, 191, 5976–5987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelatey, L.M.; Khalil, W.K.B.; Ali, T.H.; Mahrous, K.F. Heavy metal resistance and gene expression analysis of metal resistance genes in gram positive and gram negative bacteria present in Egyptian soil. J. Appl. Sci. Env. Sanit. 2011, 6, 201–211. [Google Scholar]
- Trevos, J.T.; Oddie, K.M.; Belliveau, B.H. Metal resistance in bacteria FEMS Microbiol. Rev. 1985, 32, 39–54. [Google Scholar]
- Zhang, C.G.; Xu, H.X.; Jiang, J.N.; Zhang, C.S.C.; Li, L.; Liu, Q.S. Microbial ecology in water area polluted by high concentrations of Cd2+ and Pb2+. Chin. J. Appl. Ecol. 1993, 4, 423–429. [Google Scholar]
- Lu, W.B.; Shi, J.J.; Wang, C.H.; Chang, J.S. Biosorption of lead, copper and cadmium by an indigenous isolate Enterobacter sp. J1 possessing high heavy-metal resistance. J. Hazard. Mater. 2006, 134, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Gabr, R.M.; Hassan, S.H.A.; Shoreit, A.A.M. Biosorption of lead and nickel by living and non-living cells of Pseudomonas aeruginosa ASU 6a. Int Biodeterior. Biodegrad. 2008, 62, 195–203. [Google Scholar] [CrossRef]
- Oh, S.E.; Hassan, S.H.; Joo, J.H. Biosorption of heavy metals by lyophilized cells of Pseudomonas stutzeri. World J. Microbiol. Biotechnol. 2009, 25, 1771–1778. [Google Scholar] [CrossRef]
- Suksabye, P.; Pimthong, A.; Dhurakit, P.; Mekvichitsaeng, P.; Thiravetyan, P. Effect of biochars and microorganisms on cadmium accumulation in rice grains grown in Cd-contaminated soil. Environ. Sci. Pollut. Res. 2016, 23, 962–973. [Google Scholar] [CrossRef]
- Salim, B.B.M.; Hikal, M.S.; Osman, H.S. Ameliorating the deleterious effects of saline water on the antioxidants defense system and yield of eggplant using foliar application of zinc sulphate. Ann. Agric. Sci. 2019, 64, 244–251. [Google Scholar] [CrossRef]
- Santos, D.; Duarte, B.; Caçador, I. Biochemical and photochemical feedbacks of acute Cd toxicity in Juncus acutus seedlings: The role of non-functional Cd-chlorophylls. Estuar. Coast Shelf Sci. 2015, 167, 228–239. [Google Scholar] [CrossRef]
- Akhtar, S.S.; Andersen, M.N.; Liu, F. Biochar mitigates salinity stress in potato. J. Agron. Crop Sci. 2015, 201. [Google Scholar] [CrossRef]
- Hafez, E.M.; Alsohim, A.S.; Farig, M.; Omara, A.E.-D.; Rashwan, E.; Kamara, M.M. Synergistic effect of biochar and plant growth promoting rhizobacteria on alleviation of water deficit in rice plants under salt-affected soil. Agronomy 2019, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Swarnalakshmi, K.; Yadav, V.; Tyagi, D.; Dhar, D.W.; Kannepalli, A.; Kumar, S. Significance of plant growth promoting rhizobacteria in grain legumes: Growth promotion and crop production. Plants 2020, 9, 1596. [Google Scholar] [CrossRef] [PubMed]
- Hafez, E.M.; Osman, H.S.; Gowayed, S.M.; Okasha, S.A.; Omara, A.E.-D.; Sami, R.; Abd El-Monem, A.M.; Abd El-Razek, U.A. Minimizing the adversely impacts of water deficit and soil salinity on maize growth and productivity in response to the application of plant growth-promoting Rhizobacteria and Silica nanoparticles. Agronomy 2021, 11, 676. [Google Scholar] [CrossRef]
- Hafez, E.M.; Omara, A.E.D.; Alhumaydhi, F.A.; El-Esawi, M.A. Minimizing hazard impacts of soil salinity and water stress on wheat plants by soil application of vermicompost and biochar. Phys. Plant. 2021, 172, 587–602. [Google Scholar] [CrossRef]
- Kolahi, M.; Kazemi, E.M.; Yazdi, M.; Goldson-Barnaby, A. Oxidative stress induced by cadmium in lettuce (Lactuca sativa Linn.): Oxidative stress indicators and prediction of their genes. Plant Physiol. Biochem. 2020, 146, 71–89. [Google Scholar] [CrossRef] [PubMed]
- Iannelli, M.; Pietrini, F.; Fiore, L.; Petrilli, L.; Massacci, A. Antioxidant response to cadmium in Phragmites australis plants. Plant Physiol. Biochem. 2002, 40, 977–982. [Google Scholar] [CrossRef]
- Xiao, L.; Guo, H.; Wang, S.; Li, J.; Wang, Y.; Xing, B. Carbon dots alleviate the toxicity of cadmium ions (Cd2+) toward wheat seedlings. Env. Sci. Nan. 2019, 6, 1493–1506. [Google Scholar] [CrossRef]
- Unadkat, K.; Parikh, P. Localization of Cadmium metal ion in Lemna polyrhiza L. using SEM morphology and EDX analysis. Environ. Conserv. J. 2019, 20, 81–86. [Google Scholar] [CrossRef]
- Vig, K.; Megharaj, M.; Sethunathan, N.; Naidu, R. Bioavailability and toxicity of cadmium to microorganisms and their activities in soil: A review. Adv. Env. Res. 2003, 8, 121–135. [Google Scholar] [CrossRef]
- Gomes, N.C.M.; Landi, L.; Smalla, K.; Nannipieri, P.; Brookes, P.C.; Renella, G. Effects of Cd- and Zn-enriched sewage sludge on soil bacterial and fungal communities. Ecotoxicol. Env. Saf. 2010, 73, 1255–1263. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud Univ. Sci. 2014, 26, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Sarma, B.; Buragohain, S.; Nath, D.J.; Gogoi, N. Temporal responses of soil biological characteristics to organic inputs and mineral fertilizers under wheat cultivation in inceptisol. Arch. Agron. Soil Sci. 2016, 63. [Google Scholar] [CrossRef]
- Liu, X.; Hu, X.; Zhang, X.; Chen, X.; Chen, J.; Yuan, X. Effect of Bacillus subtilis and NTA-APG on pyrene dissipation in phytoremediation of nickel co-contaminated wetlands by Scirpus triqueter. Ecotoxicol. Env. Saf. 2018, 154, 69–74. [Google Scholar] [CrossRef]
- El-Nahrawy, S.; Elhawat, N.; Alshaal, T. Biochemical traits of Bacillus subtilis MF497446: Its implications on the development of cowpea under cadmium stress and ensuring food safety. Ecotoxicol. Env. Saf. 2019, 180, 384–395. [Google Scholar] [CrossRef]
- Zhai, Y.; Wei, X.; Zeng, G.; Zhang, D.; Chu, K. Study of adsorbent derived from sewage sludge for the removal of Cd2+, Ni2+ in aqueous solution. Sep. Purif. Technol. 2004, 38, 191–196. [Google Scholar] [CrossRef]
- Tajar, A.F.; Kaghazchi, T.; Soleimani, M. Adsorption of cadmium from aqueous solutions on sulfurized activated carbon prepared from nut shells. J. Hazard. Mater. 2009, 165, 1159–1164. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatments | Fresh Weight (g Plant−1) | Dry Weight (g Plant−1) | Plant Height (cm Plant−1) | Leaf Area (cm2) | Number of Leaves (Plant−1) |
|---|---|---|---|---|---|
| First season (2020) | |||||
| 0 T1 | 387.00 ± 5.29 h | 33.88 ± 0.48 h | 34.30 ± 0.51 d | 10.74 ± 0.16 d | 36.80 ± 0.54 d |
| 0 T2 | 481.33 ± 5.13 c | 42.59 ± 0.56 c | 36.17 ± 0.59 c | 11.31 ± 0.18 c | 38.78 ± 0.62 c |
| 0 T3 | 427.33 ± 4.51 e | 37.75 ± 0.41 e | 33.79 ± 0.61 d | 10.58 ± 0.19 d | 36.26 ± 0.65 d |
| 0 T4 | 512.00 ± 6.24 a | 45.56 ± 0.57 a | 43.03 ± 0.97 a | 13.41 ± 0.30 a | 46.04 ± 1.02 a |
| 40 T1 | 354.33 ± 4.51 j | 30.91 ± 0.41 j | 28.12 ± 0.43 f | 8.85 ± 0.13 f | 30.26 ± 0.45 f |
| 40 T2 | 459.00 ± 4.58 d | 40.63 ± 0.42 d | 31.86 ± 0.87 e | 9.99 ± 0.27 e | 34.22 ± 0.92 e |
| 40 T3 | 401.67 ± 4.16 fg | 35.45 ± 0.32 fg | 32.09 ± 0.74 e | 10.06 ± 0.23 e | 34.46 ± 0.78 e |
| 40 T4 | 491.67 ± 5.03 b | 43.71 ± 0.46 b | 38.66 ± 0.87 b | 12.07 ± 0.27 b | 41.42 ± 0.92 b |
| 80 T1 | 336.67 ± 4.04 k | 29.31 ± 0.37 k | 26.25 ± 1.09 g | 8.28 ± 0.33 g | 28.28 ± 1.16 g |
| 80 T2 | 453.67 ± 2.08 d | 40.14 ± 0.19 d | 29.26 ± 0.64 f | 9.20 ± 0.20 f | 31.46 ± 0.68 f |
| 80 T3 | 399.00 ± 4.36 g | 35.28 ± 0.40 g | 28.97 ± 0.52 f | 9.11 ± 0.16 f | 31.16 ± 0.55 f |
| 80 T4 | 479.33 ± 4.04 c | 42.59 ± 0.37 c | 33.79 ± 1.06 d | 10.58 ± 0.32 d | 36.26 ± 1.12 d |
| 120 T1 | 298.67 ± 6.81 l | 25.85 ± 0.62 l | 13.84 ± 0.71 j | 4.48 ± 0.22 j | 15.14 ± 0.75 j |
| 120 T2 | 408.67 ± 6.11 f | 36.05 ± 0.56 f | 16.73 ± 0.94 i | 5.37 ± 0.29 i | 18.20 ± 0.99 i |
| 120 T3 | 366.00 ± 3.61 i | 32.28 ± 0.33 i | 17.87 ± 0.43 hi | 5.71 ± 0.13 hi | 19.40 ± 0.45 hi |
| 120 T4 | 452.00 ± 4.36 d | 40.10 ± 0.40 d | 18.89 ± 0.64 h | 6.03 ± 0.20 h | 20.48 ± 0.68 h |
| Second season (2021) | |||||
| 0 T1 | 389.33 ± 4.93 h | 34.55 ± 0.86 h | 34.39 ± 0.50 d | 10.75 ± 0.16 d | 37.68 ± 0.54 d |
| 0 T2 | 484.67 ± 4.93 c | 43.26 ± 0.84 c | 36.22 ± 0.55 c | 11.33 ± 0.18 c | 39.66 ± 0.62 c |
| 0 T3 | 430.67 ± 5.69 e | 38.97 ± 0.41 f | 33.83 ± 0.61 d | 10.61 ± 0.18 d | 37.14 ± 0.65 d |
| 0 T4 | 514.00 ± 6.24 a | 46.78 ± 0.57 a | 43.07 ± 0.97 a | 13.42 ± 0.31 a | 46.92 ± 1.02 a |
| 40 T1 | 357.33 ± 2.08 j | 31.58 ± 0.95 i | 28.17 ± 0.39 f | 8.87 ± 0.13 f | 31.14 ± 0.45 f |
| 40 T2 | 461.33 ± 4.93 d | 42.63 ± 1.35 cd | 31.82 ± 0.79 e | 10.03 ± 0.25 e | 35.10 ± 0.92 e |
| 40 T3 | 404.00 ± 4.58 g | 36.67 ± 0.32 g | 32.13 ± 0.75 e | 10.08 ± 0.22 e | 35.34 ± 0.78 e |
| 40 T4 | 493.33 ± 5.51 b | 44.93 ± 0.46 b | 38.70 ± 0.87 b | 12.10 ± 0.28 b | 42.30 ± 0.92 b |
| 80 T1 | 339.33 ± 3.21 k | 29.97 ± 0.27 j | 26.28 ± 1.11 g | 8.29 ± 0.33 g | 29.16 ± 1.16 g |
| 80 T2 | 456.33 ± 2.08 d | 41.14 ± 1.18 de | 29.28 ± 0.64 f | 9.20 ± 0.20 f | 32.34 ± 0.68 f |
| 80 T3 | 403.67 ± 6.66 g | 36.50 ± 0.40 g | 29.01 ± 0.52 f | 9.13 ± 0.16 f | 32.04 ± 0.55 f |
| 80 T4 | 481.67 ± 4.51 c | 44.14 ± 0.90 bc | 33.84 ± 1.04 d | 10.59 ± 0.33 d | 37.14 ± 1.12 d |
| 120 T1 | 302.00 ± 7.81 l | 24.18 ± 2.70 k | 13.91 ± 0.67 j | 4.50 ± 0.21 j | 16.02 ± 0.75 j |
| 120 T2 | 412.67 ± 5.13 f | 35.94 ± 0.72 gh | 16.75 ± 0.95 i | 5.39 ± 0.29 i | 19.08 ± 0.99 i |
| 120 T3 | 369.33 ± 4.93 i | 32.17 ± 0.28 i | 17.91 ± 0.43 hi | 5.73 ± 0.13 hi | 20.28 ± 0.45 hi |
| 120 T4 | 454.33 ± 4.73 d | 40.65 ± 0.82 e | 18.92 ± 0.65 h | 6.05 ± 0.17 h | 21.36 ± 0.68 h |
| F-test | |||||
| Main | *** | *** | *** | *** | *** |
| Sub main | *** | *** | *** | *** | *** |
| Interaction | *** | *** | *** | *** | *** |
| Treatments | Total Chlorophyll (mg g−1 FW) | Carotenoids (μg g−1 FW) | TSS (μg g−1 FW) | Proline (µmol g−1 FW) |
|---|---|---|---|---|
| First season (2020) | ||||
| 0 T1 | 2.20 ± 0.03 d | 0.75 ± 0.04 d | 5.78 ± 0.07 d | 8.13 ± 0.10 ef |
| 0 T2 | 2.31 ± 0.03 c | 0.94 ± 0.02 a | 6.03 ± 0.08 c | 8.08 ± 0.05 f |
| 0 T3 | 2.17 ± 0.04 d | 0.91 ± 0.01 ab | 5.71 ± 0.08 d | 7.46 ± 0.05 hi |
| 0 T4 | 2.71 ± 0.06 a | 0.96 ± 0.02 a | 6.96 ± 0.13 a | 7.22 ± 0.25 j |
| 40 T1 | 1.84 ± 0.03 f | 0.64 ± 0.02 f | 4.94 ± 0.06 f | 8.60 ± 0.05 d |
| 40 T2 | 2.06 ± 0.05 e | 0.86 ± 0.05 b | 5.45 ± 0.12 e | 7.54 ± 0.25 gh |
| 40 T3 | 2.07 ± 0.04 e | 0.81 ± 0.02 c | 5.48 ± 0.10 e | 7.28 ± 0.05 ij |
| 40 T4 | 2.46 ± 0.05 b | 0.89 ± 0.05 b | 6.37 ± 0.12 b | 6.17 ± 0.08 k |
| 80 T1 | 1.73 ± 0.06 g | 0.53 ± 0.08 g | 4.69 ± 0.15 g | 8.77 ± 0.14 cd |
| 80 T2 | 1.90 ± 0.04 f | 0.71 ± 0.03 de | 5.10 ± 0.09 f | 8.30 ± 0.20 e |
| 80 T3 | 1.89 ± 0.03 f | 0.68 ± 0.04 ef | 5.06 ± 0.07 f | 7.68 ± 0.20 g |
| 80 T4 | 2.17 ± 0.06 d | 0.70 ± 0.05 e | 5.71 ± 0.14 d | 7.49 ± 0.20 gh |
| 120 T1 | 1.00 ± 0.04 j | 0.30 ± 0.02 i | 3.01 ± 0.10 j | 9.94 ± 0.15 a |
| 120 T2 | 1.17 ± 0.06 i | 0.37 ± 0.02 h | 3.40 ± 0.13 i | 9.60 ± 0.20 b |
| 120 T3 | 1.23 ± 0.03 hi | 0.33 ± 0.03 hi | 3.56 ± 0.06 hi | 8.97 ± 0.20 c |
| 120 T4 | 1.29 ± 0.04 h | 0.35 ± 0.02 h | 3.69 ± 0.09 h | 8.79 ± 0.20 cd |
| Second season (2021) | ||||
| 0 T1 | 2.38 ± 0.03 d | 0.84 ± 0.04 d | 5.89 ± 0.07 d | 8.19 ± 0.10 ef |
| 0 T2 | 2.48 ± 0.02 c | 1.02 ± 0.01 a | 6.17 ± 0.08 c | 8.16 ± 0.05 f |
| 0 T3 | 2.34 ± 0.02 d | 0.99 ± 0.01 ab | 5.85 ± 0.08 d | 7.55 ± 0.05 gh |
| 0 T4 | 2.90 ± 0.06 a | 1.03 ± 0.02 a | 7.09 ± 0.13 a | 7.27 ± 0.25 i |
| 40 T1 | 2.02 ± 0.03 f | 0.73 ± 0.02 f | 5.05 ± 0.06 g | 8.66 ± 0.05 d |
| 40 T2 | 2.22 ± 0.05 e | 0.94 ± 0.05 b | 5.59 ± 0.12 e | 7.63 ± 0.25 g |
| 40 T3 | 2.26 ± 0.04 e | 0.89 ± 0.02 c | 5.61 ± 0.10 e | 7.37 ± 0.05 hi |
| 40 T4 | 2.65 ± 0.05 b | 0.96 ± 0.05 b | 6.50 ± 0.12 b | 6.22 ± 0.08 j |
| 80 T1 | 1.91 ± 0.06 g | 0.62 ± 0.08 g | 4.80 ± 0.15 h | 8.83 ± 0.14 cd |
| 80 T2 | 2.06 ± 0.04 f | 0.79 ± 0.03 e | 5.24 ± 0.09 f | 8.39 ± 0.20 e |
| 80 T3 | 2.08 ± 0.03 f | 0.76 ± 0.04 ef | 5.19 ± 0.07 fg | 7.73 ± 0.20 g |
| 80 T4 | 2.36 ± 0.06 d | 0.77 ± 0.05 ef | 5.84 ± 0.14 d | 7.54 ± 0.20 gh |
| 120 T1 | 1.18 ± 0.04 j | 0.39 ± 0.02 i | 3.12 ± 0.10 k | 10.00 ± 0.15 a |
| 120 T2 | 1.33 ± 0.06 i | 0.45 ± 0.02 h | 3.54 ± 0.13 j | 9.69 ± 0.20 b |
| 120 T3 | 1.42 ± 0.03 h | 0.41 ± 0.03 hi | 3.69 ± 0.06 ij | 9.02 ± 0.20 c |
| 120 T4 | 1.48 ± 0.04 h | 0.43 ± 0.02 hi | 3.82 ± 0.09 i | 8.84 ± 0.20 cd |
| F-test | ||||
| Main | *** | *** | *** | *** |
| Sub main | *** | *** | *** | *** |
| Interaction | *** | *** | *** | *** |
| Treatments | Microbial Biomass Carbon (mg g−1 Soil) | CO2 Evaluation (mg CO2/100 g Soil) | Dehydrogenase (DHA) (mg TPF g−1 Soil d−1) |
|---|---|---|---|
| First season (2020) | |||
| 0 T1 | 4.40 ± 0.03 d | 110.00 ± 5.29 h | 143.00 ± 5.29 h |
| 0 T2 | 4.37 ± 0.04 d | 150.33 ± 4.51 e | 183.33 ± 4.51 e |
| 0 T3 | 4.51 ± 0.03 c | 204.33 ± 5.13 c | 237.33 ± 5.13 c |
| 0 T4 | 4.91 ± 0.06 a | 235.00 ± 6.24 a | 268.00 ± 6.24 a |
| 40 T1 | 4.04 ± 0.03 f | 77.33 ± 4.51 j | 110.33 ± 4.51 j |
| 40 T2 | 4.27 ± 0.04 e | 124.67 ± 4.16 fg | 157.67 ± 4.16 fg |
| 40 T3 | 4.26 ± 0.05 e | 182.00 ± 4.58 d | 215.00 ± 4.58 d |
| 40 T4 | 4.66 ± 0.05 b | 214.67 ± 5.03 b | 247.67 ± 5.03 b |
| 80 T1 | 3.93 ± 0.06 g | 59.67 ± 4.04 k | 92.67 ± 4.04 k |
| 80 T2 | 4.09 ± 0.03 f | 122.00 ± 4.36 g | 155.00 ± 4.36 g |
| 80 T3 | 4.10 ± 0.04 f | 176.67 ± 2.08 d | 209.67 ± 2.08 d |
| 80 T4 | 4.37 ± 0.06 d | 202.33 ± 4.04 c | 235.33 ± 4.04 c |
| 120 T1 | 3.20 ± 0.04 j | 21.67 ± 6.81 l | 54.67 ± 6.81 l |
| 120 T2 | 3.43 ± 0.03 hi | 89.00 ± 3.61 i | 122.00 ± 3.61 i |
| 120 T3 | 3.37 ± 0.06 i | 131.67 ± 6.11 f | 164.67 ± 6.11 f |
| 120 T4 | 3.49 ± 0.04 gh | 175.00 ± 4.36 d | 208.00 ± 4.36 d |
| Second season (2021) | |||
| 0 T1 | 4.62 ± 0.03 cd | 116.00 ± 5.29 h | 147.70 ± 5.29 h |
| 0 T2 | 4.56 ± 0.06 d | 156.17 ± 4.25 e | 188.63 ± 5.46 e |
| 0 T3 | 4.69 ± 0.03 c | 209.83 ± 5.13 c | 243.83 ± 5.13 c |
| 0 T4 | 5.11 ± 0.06 a | 242.80 ± 6.24 a | 275.00 ± 6.24 a |
| 40 T1 | 4.26 ± 0.03 f | 83.33 ± 4.51 j | 115.03 ± 4.51 j |
| 40 T2 | 4.45 ± 0.04 e | 130.17 ± 4.16 g | 164.17 ± 4.16 g |
| 40 T3 | 4.44 ± 0.05 e | 187.50 ± 4.58 d | 221.50 ± 4.58 d |
| 40 T4 | 4.86 ± 0.05 b | 222.47 ± 5.03 b | 254.67 ± 5.03 b |
| 80 T1 | 4.15 ± 0.06 g | 65.67 ± 4.04 k | 97.37 ± 4.04 k |
| 80 T2 | 4.27 ± 0.03 f | 127.50 ± 4.36 g | 161.50 ± 4.36 g |
| 80 T3 | 4.30 ± 0.04 f | 183.70 ± 0.85 d | 216.50 ± 1.80 d |
| 80 T4 | 4.57 ± 0.06 d | 210.13 ± 4.04 c | 242.33 ± 4.04 c |
| 120 T1 | 3.42 ± 0.04 j | 27.67 ± 6.81 l | 59.37 ± 6.81 l |
| 120 T2 | 3.61 ± 0.03 i | 94.50 ± 3.61 i | 128.50 ± 3.61 i |
| 120 T3 | 3.57 ± 0.06 i | 139.47 ± 6.11 f | 171.67 ± 6.11 f |
| 120 T4 | 3.69 ± 0.04 h | 182.80 ± 4.36 d | 215.00 ± 4.36 d |
| F-test | |||
| Main | *** | *** | *** |
| Sub main | *** | *** | *** |
| Interaction | *** | *** | *** |
| Treatments | Cd Content in Root (µg g−1) | Cd Content in Shoots (µg g−1) | Bioconcentration Factor (BCF) | Translocation Factor (TF) |
|---|---|---|---|---|
| First season (2020) | ||||
| 0 T1 | 0.00 | 0.00 | 0.00 | 0.00 |
| 0 T2 | 0.00 | 0.00 | 0.00 | 0.00 |
| 0 T3 | 0.00 | 0.00 | 0.00 | 0.00 |
| 0 T4 | 0.00 | 0.00 | 0.00 | 0.00 |
| 40 T1 | 23.53 ± 1.00 f | 15.19 ± 1.00 f | 0.58 ± 0.02 c | 0.64 ± 0.02 d |
| 40 T2 | 12.08 ± 0.75 g | 3.74 ± 0.75 g | 0.30 ± 0.02 ef | 0.30 ± 0.04 f |
| 40 T3 | 13.61 ± 0.33 g | 5.27 ± 0.33 g | 0.34 ± 0.01 d | 0.38 ± 0.02 e |
| 40 T4 | 7.84 ± 0.33 h | 5.54 ± 0.98 g | 0.19 ± 0.01 h | 0.70 ± 0.10 c |
| 80 T1 | 65.17 ± 7.66 b | 56.83 ± 7.66 b | 0.81 ± 0.10 a | 0.87 ± 0.02 a |
| 80 T2 | 22.03 ± 1.21 f | 13.69 ± 1.21 f | 0.27 ± 0.02 ef | 0.62 ± 0.02 d |
| 80 T3 | 27.49 ± 2.55 e | 19.15 ± 2.55 e | 0.34 ± 0.03 d | 0.69 ± 0.03 c |
| 80 T4 | 21.72 ± 2.55 f | 13.38 ± 2.55 f | 0.27 ± 0.03 ef | 0.61 ± 0.05 d |
| 120 T1 | 92.71 ± 3.50 a | 84.37 ± 3.50 a | 0.77 ± 0.03 b | 0.91 ± 0.00 a |
| 120 T2 | 27.96 ± 1.16 de | 19.62 ± 1.16 de | 0.23 ± 0.01 gh | 0.70 ± 0.01 c |
| 120 T3 | 36.67 ± 1.17 c | 28.33 ± 1.17 c | 0.30 ± 0.01 de | 0.77 ± 0.01 b |
| 120 T4 | 30.90 ± 1.17 d | 22.56 ± 1.17 d | 0.25 ± 0.01 fg | 0.73 ± 0.01 bc |
| Second season (2021) | ||||
| 0 T1 | 0.00 | 0.00 | 0.00 | 0.00 |
| 0 T2 | 0.00 | 0.00 | 0.00 | 0.00 |
| 0 T3 | 0.00 | 0.00 | 0.00 | 0.00 |
| 0 T4 | 0.00 | 0.00 | 0.00 | 0.00 |
| 40 T1 | 24.61 ± 1.26 f | 15.24 ± 0.96 f | 0.61 ± 0.02 c | 0.61 ± 0.03 ef |
| 40 T2 | 12.10 ± 0.74 h | 3.81 ± 0.74 g | 0.30 ± 0.02 ef | 0.31 ± 0.04 h |
| 40 T3 | 13.95 ± 0.84 h | 5.32 ± 0.35 g | 0.34 ± 0.02 d | 0.38 ± 0.01 g |
| 40 T4 | 8.34 ± 0.82 i | 5.60 ± 0.98 g | 0.20 ± 0.02 h | 0.67 ± 0.09 de |
| 80 T1 | 65.22 ± 6.99 b | 56.89 ± 7.66 b | 0.81 ± 0.10 a | 0.87 ± 0.03 a |
| 80 T2 | 23.05 ± 1.03 fg | 13.78 ± 1.18 f | 0.28 ± 0.02 ef | 0.59 ± 0.04 f |
| 80 T3 | 27.83 ± 3.01 e | 19.52 ± 2.96 e | 0.34 ± 0.04 d | 0.70 ± 0.03 cd |
| 80 T4 | 21.91 ± 2.31 g | 13.75 ± 3.01 f | 0.27 ± 0.03 ef | 0.62 ± 0.07 ef |
| 120 T1 | 91.10 ± 3.64 a | 84.43 ± 3.55 a | 0.77 ± 0.03 b | 0.92 ± 0.01 a |
| 120 T2 | 28.29 ± 1.31 e | 19.77 ± 1.23 e | 0.23 ± 0.01 gh | 0.69 ± 0.02 cd |
| 120 T3 | 37.34 ± 1.79 c | 28.92 ± 1.63 c | 0.31 ± 0.01 de | 0.77 ± 0.02 b |
| 120 T4 | 31.28 ± 1.39 d | 23.56 ± 2.16 d | 0.26 ± 0.01 fg | 0.75 ± 0.04 bc |
| F-test | ||||
| Main | *** | *** | *** | *** |
| Sub main | *** | *** | *** | *** |
| Interaction | *** | *** | *** | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elbagory, M.; Farrag, D.K.; Hashim, A.M.; Omara, A.E.-D. The Combined Effect of Pseudomonas stutzeri and Biochar on the Growth Dynamics and Tolerance of Lettuce Plants (Lactuca sativa) to Cadmium Stress. Horticulturae 2021, 7, 430. https://doi.org/10.3390/horticulturae7110430
Elbagory M, Farrag DK, Hashim AM, Omara AE-D. The Combined Effect of Pseudomonas stutzeri and Biochar on the Growth Dynamics and Tolerance of Lettuce Plants (Lactuca sativa) to Cadmium Stress. Horticulturae. 2021; 7(11):430. https://doi.org/10.3390/horticulturae7110430
Chicago/Turabian StyleElbagory, Mohssen, Diaa K. Farrag, Ahmed M. Hashim, and Alaa El-Dein Omara. 2021. "The Combined Effect of Pseudomonas stutzeri and Biochar on the Growth Dynamics and Tolerance of Lettuce Plants (Lactuca sativa) to Cadmium Stress" Horticulturae 7, no. 11: 430. https://doi.org/10.3390/horticulturae7110430
APA StyleElbagory, M., Farrag, D. K., Hashim, A. M., & Omara, A. E. -D. (2021). The Combined Effect of Pseudomonas stutzeri and Biochar on the Growth Dynamics and Tolerance of Lettuce Plants (Lactuca sativa) to Cadmium Stress. Horticulturae, 7(11), 430. https://doi.org/10.3390/horticulturae7110430