
Development of Molecular Markers Associated with Resistance to Gray Mold Disease in Onion (Allium cepa L.) through RAPD-PCR and Transcriptome Analysis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Genomic DNA Extraction
2.2. RAPD and Phylogenetic Analysis
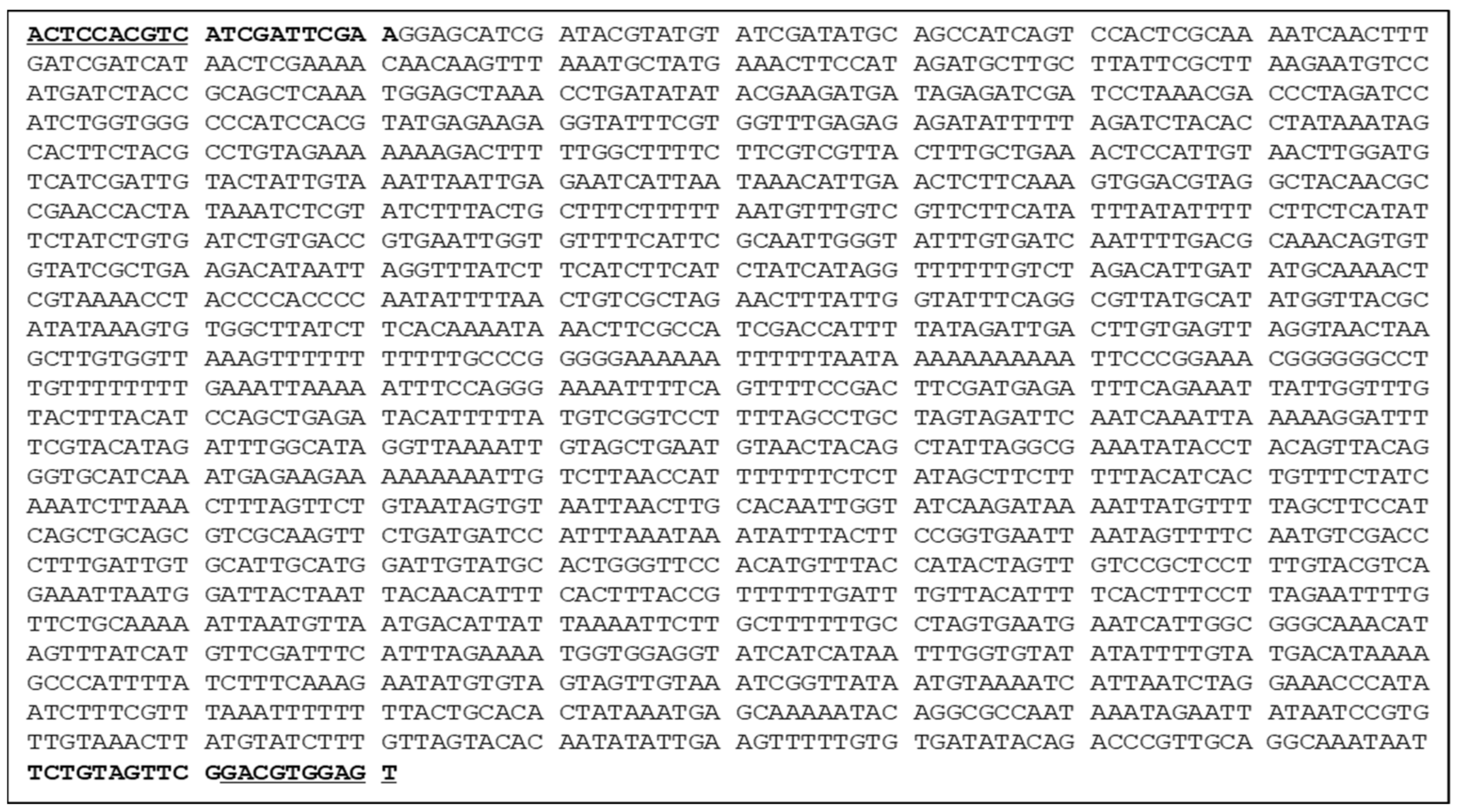
2.3. Development of the SCAR Marker
2.4. RNA Sequencing and Variant Analysis
2.5. Differentially Expressed Gene (DEG) and Gene Ontology (GO) Analysis for Selection of Transcript Related to Disease Resistance
2.6. HRM Primer Designs from Selected Transcripts and HRM Analysis
2.7. Quantitative Real-Time PCR (qPCR) to Identify Expression Level of Transcripts
3. Results
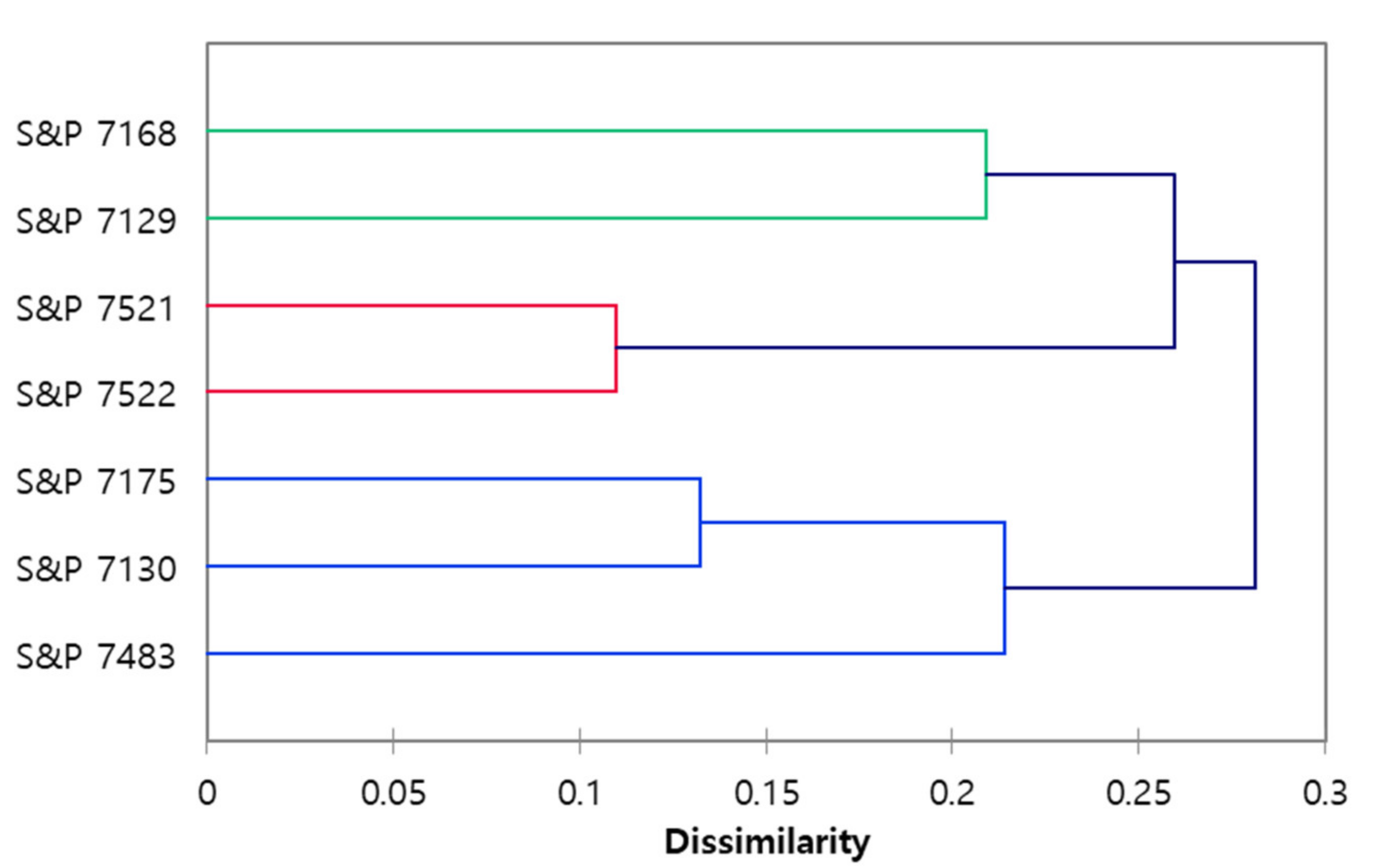
3.1. RAPD and Phylogenetic Analysis
3.2. Development of a SCAR Marker for the Selection of Resistant Lines
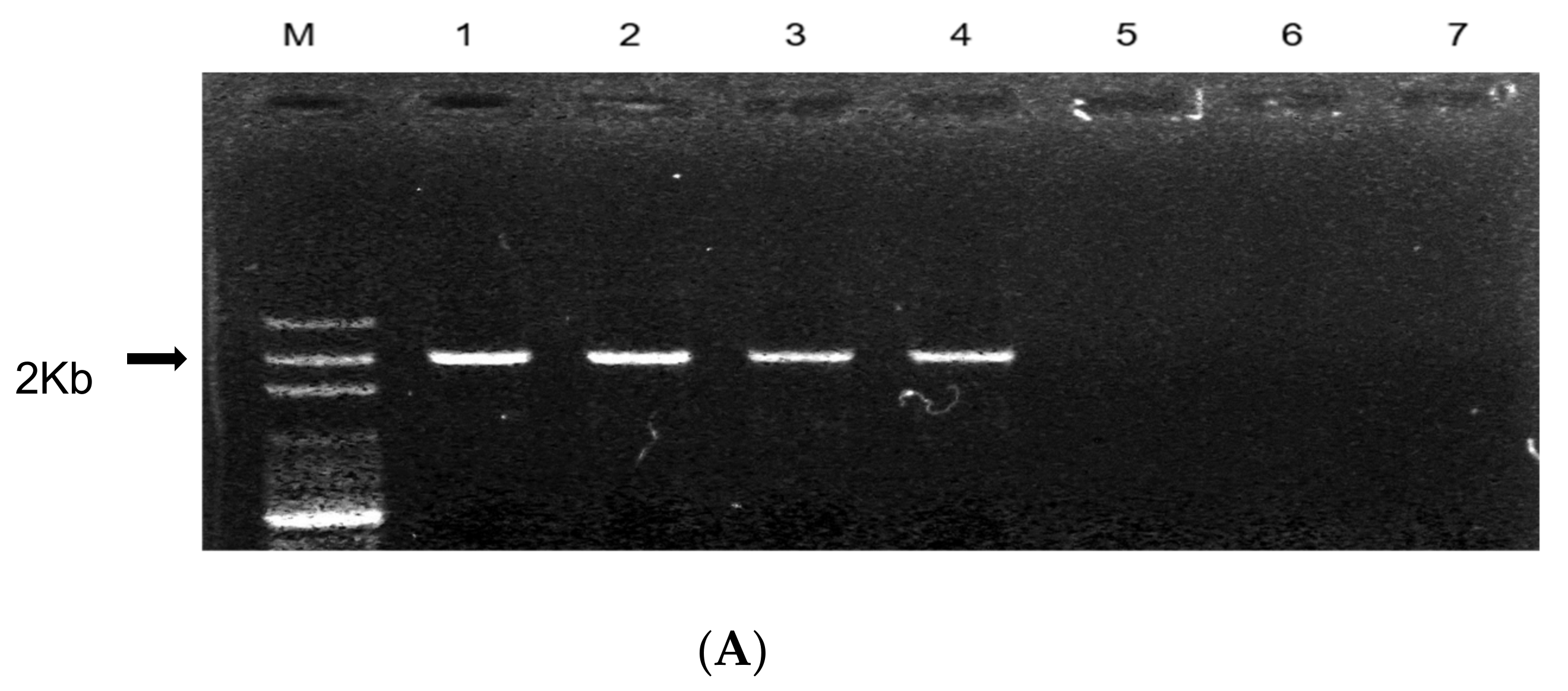
3.3. Validation of the Developed SCAR Marker Using the Resistant and Susceptible Onion Lines
3.4. Preprocessing of Raw Data of the RNA Sequence
3.5. Variant Analysis and Filtering for Selection of Transcripts
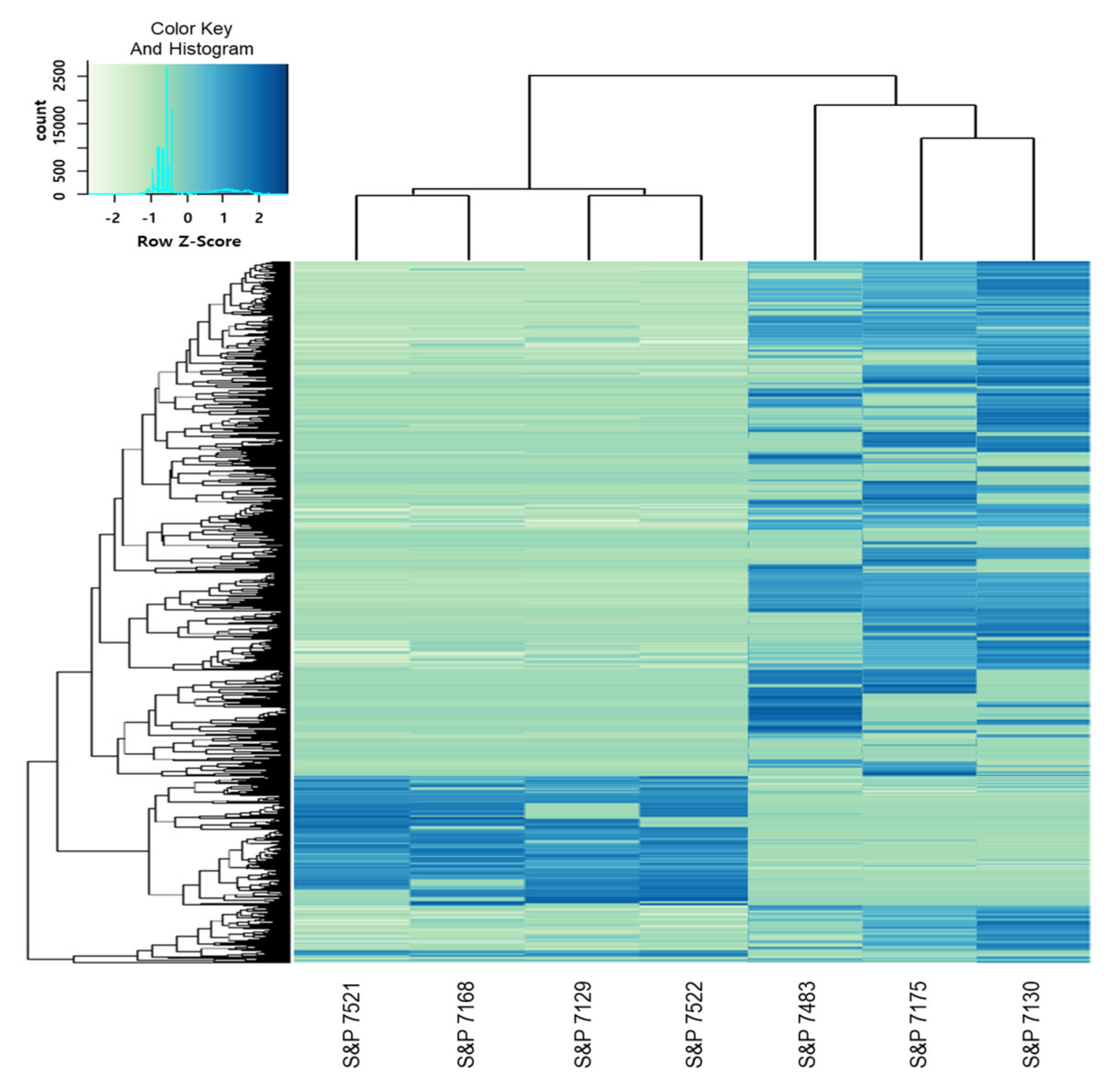
3.6. Selection of Transcripts Related to Disease Resistance through DEG Analysis and GO Annotation
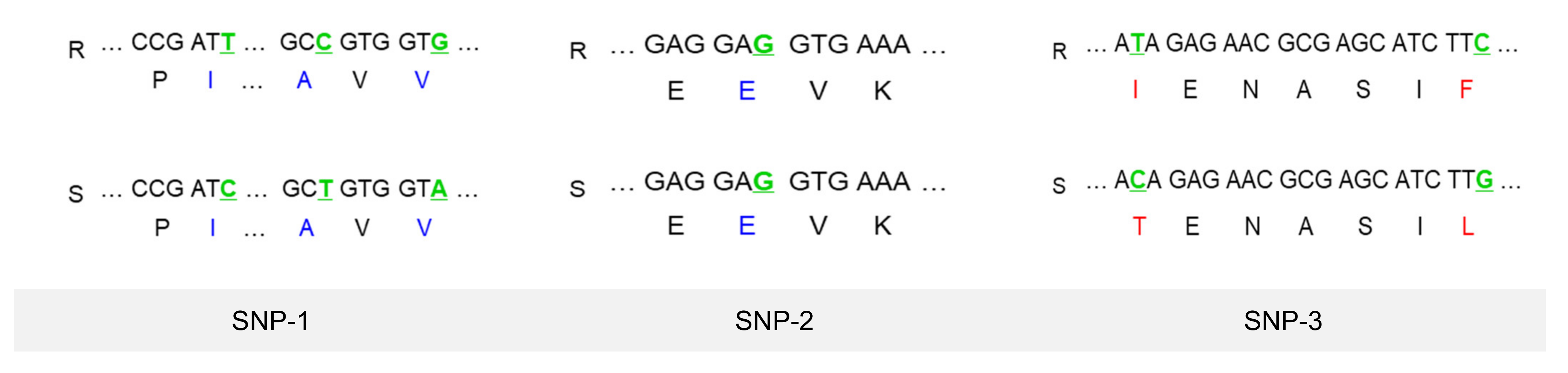
3.7. Transcripts Selection to Develop SNP Markers and a HRM Analysis
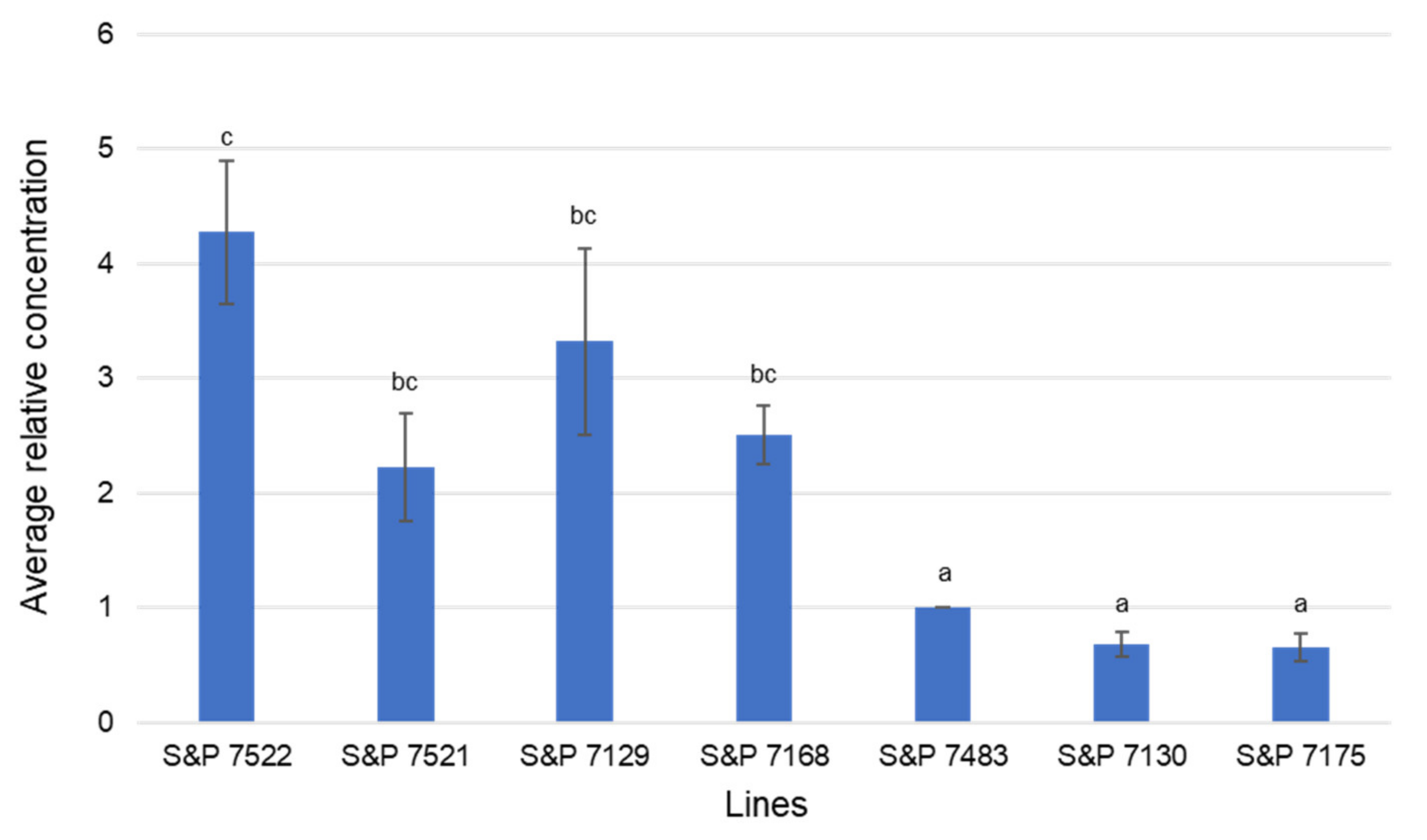
3.8. Confirmation of AKR Gene Expression Level in the Onion Lines through qPCR Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nam, K.H.; Baik, H.W.; Choi, T.Y.; Yoon, S.G.; Park, S.W.; Joung, H.J. Effects of ethanol extract of onion on the lipid profiles in patients with hypercholesterolemia. J. Nutr. Health 2007, 40, 242–248. [Google Scholar]
- Kim, J.Y.; Seo, Y.J.; Noh, S.K.; Cha, Y.J. Concentrated onion extract lowers serum lipid levels in rats fed a high-fat diet. Korean J. Food Preserv. 2010, 17, 398–404. [Google Scholar]
- Khosa, J.S.; McCallum, J.; Dhatt, A.S.; Macknight, R.C. Enhancing onion breeding using molecular tools. Plant. Breed. 2016, 135, 9–20. [Google Scholar] [CrossRef]
- Jo, J.; Purushotham, P.M.; Han, K.; Lee, H.R.; Nah, G.; Kang, B.C. Development of a genetic map for onion (Allium cepa L.) using reference-free genotyping-by-sequencing and SNP assays. Front. Plant. Sci. 2017, 8, 1606. [Google Scholar]
- Havey, M.J. Onion Breeding. In Plant Breeding Review; Wiley: New Jersey, NJ, USA, 2018; Volume 42, pp. 39–85. [Google Scholar]
- Chilvers, M.I.; du Toit, L.J. Detection and identification of Botrytis species associated with neck rot, scape blight, and umbel blight of onion. Plant. Health Prog. 2006, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; van Kan, J.A.L. Botrytis cinerea: The cause of grey mould disease. Mol. Plant. Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Choquer, M.; Fournier, E.; Kunz, C.; Levis, C.; Pradier, J.M.; Simon, A.; Viaud, M. Botrytis cinerea virulence factors: New insights into a necrotrophic and polyphageous pathogen. FEMS Microbiol. Lett. 2007, 277, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Carisse, O.; Tremblay, D.M.; McDonald, M.R.; Brodeur, L.; McRoberts, N. Management of botrytis leaf blight of onion: The Québec experience of 20 years of continual improvement. Plant. Dis. 2001, 95, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Abbey, J.A.; Percival, D.; Abbey, L.; Asiedu, S.K.; Prithiviraj, B.; Schilder, A. Biofungicides as alternative to synthetic fungicide control of grey mould (Botrytis cinerea)—prospects and challenges. Biocontrol. Sci. Technol. 2018, 29, 207–228. [Google Scholar] [CrossRef]
- Howell, G.S. Sustainable grape productivity and the growth-yield relationship: A review. Am. J. Enol. Vitic. 2001, 52, 165–174. [Google Scholar]
- Tripathi, P.; Dubey, N.K.; Shukla, A.K. Use of some essential oils as post-harvest botanical fungicides in the management of grey mould of grapes caused by Botrytis cinerea. World J. Microbiol. Biotechnol. 2008, 24, 39–46. [Google Scholar] [CrossRef]
- Fan, F.; Hamada, M.S.; Li, N.; Li, G.Q.; Luo, C.X. Multiple fungicide resistance in Botrytis cinerea from greenhouse strawberries in Hubei province, China. Plant. Dis. 2017, 101, 601–606. [Google Scholar] [CrossRef] [Green Version]
- Rupp, S.; Weber, R.W.S.; Rieger, D.; Detzel, P.; Hahn, M. Spread of Botrytis cinerea strains with multiple fungicide resistance in German horticulture. Front. Microbiol. 2017, 7, 2075. [Google Scholar] [CrossRef] [Green Version]
- Maniruzzaman, M.; Haque, M.E.; Haque, M.M.; Sayem, M.A.; Al-Amin, M. Molecular characterization of onion (Allium cepa) using RAPD markers. Bangladesh J. Agric. Res. 2010, 35, 313–322. [Google Scholar]
- Sudha, G.S.; Ramesh, P.; Sekhar, A.C.; Krishna, T.S.; Bramhachari, P.V.; Riazunnisa, K. Genetic diversity analysis of selected onion (Allium cepa L.) germplasm using specific RAPD and ISSR polymorphism markers. Biocatal. Agric. Biotechnol. 2019, 17, 110–118. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Thamllarasan, S.K.; Natarajan, S.; Park, J.I.; Chung, M.Y.; Nou, I.S. De novo assembly and transcriptome analysis of bulb onion (Allium cepa L.) during cold acclimation using contrasting genotypes. PLoS ONE 2016, 11, e0161987. [Google Scholar] [CrossRef]
- Graham, R.; Liew, M.; Meadows, C.; Lyon, E.; Wittwer, C.T. Distinguishing different DNA Heterozygotes by high-resolution melting. Clin. Chem. 2005, 51, 1295–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, G.H.; Kent, J.O.; Wittwer, C.T. High-resolution DNA melting analysis for simple and efficient molecular diagnostics. Pharmacogenomics 2007, 8, 597–608. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Sokal, R.R.; Michener, C.D. A statistical method for evaluation systematic relationships. Univ. Kans. Sci. Bull. 1958, 28, 1409–1438. [Google Scholar]
- Jaccard, P. Étude comparative de la distribution florale dans une portion des Alpes et des Jura. Bull. Soc. Vaud. Sci. Nat. 1901, 37, 547–579. [Google Scholar]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Welsh, J.; McClelland, M. Fingerprinting genomes using PCR with arbitrary primers. Nucleic. Acids Res. 1990, 18, 7213–7218. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.G.K.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic. Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [Green Version]
- Sidiq, Y.; Subiastuti, A.S.; Wibowo, W.A.; Daryono, B.S. Development of SCAR marker linked to begomovirus resistance in melon (Cucumis melo L.). Jordan J. Biol. Sci. 2020, 13, 145–151. [Google Scholar]
- Baite, M.S.; Upadhyay, B.K.; Dube, S.C. Development of a sequence-characterized amplified region marker for detection of Ascochyta rabiei causing Ascochyta blight in chickpea. Folia Microbiol. 2020, 65, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Mei, Z.; Khan, M.A. Genetic authentication of Eclipta prostrate (Asteraceae) from Penthorum chinense (Penthoraceae) by sequence characterized amplified region (SCAR) markers. Rev. Biol. Trop. 2020, 68, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.Y.; Huo, Y.M.; Miao, J.; Liu, B.J.; Kong, S.P.; Gao, L.M.; Liu, C.; Wang, Z.B.; Tahara, Y.; Kitano, H.; et al. Identification of two SCAR markers co-segregated with the dominant Ms and recessive ms alleles in onion (Allium cepa L.). Euphytica 2013, 190, 267–277. [Google Scholar] [CrossRef]
- Kim, G.H.; Yun, H.K.; Choi, C.S.; Park, J.H.; Jung, Y.J.; Park, K.S.; Dane, F.; Kang, K.K. Identification of AFLP and RAPD markers linked to anthracnose resistance in grapes and their conversion to SCAR markers. Plant. Breed. 2008, 127, 418–423. [Google Scholar] [CrossRef]
- Chowdhury, M.A.; Andrahennadi, C.P.; Slinkard, A.E.; Vandenberg, A. RAPD and SCAR markers for resistance to acochyta blight in lentil. Euphytica 2001, 118, 331–337. [Google Scholar] [CrossRef]
- Hunt, M.D.; Ryals, J.A.; Reinhardt, D. Systemic acquired resistance signal transduction. Crit. Rev. Plant. Sci. 1996, 15, 583–606. [Google Scholar] [CrossRef]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Ward, E.R.; Uknes, S.J.; Williams, S.C.; Dincher, S.S.; Wiederhold, D.L.; Alexander, D.C.; Aphl-Goy, P.; Metraux, J.P.; Ryals, J.A. Coordinate gene activity in response to agents that induce systemic acquired resistance. Plant. Cell 1991, 3, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Sticher, L.; Mauch-Mani, B.; Métraux, A.J. Systemic acquired resistance. Annu. Rev. Phytopathol. 1997, 35, 235–270. [Google Scholar] [CrossRef] [PubMed]
- Mauch-Mani, B.; Métraux, J.P. Salicylic acid and systemic acquired resistance to pathogen attack. Ann. Bot. 1998, 82, 535–540. [Google Scholar] [CrossRef] [Green Version]
- Frías, M.; Brito, N.; González, C. The Botrytis cinerea cerato-platanin BcSpl1 is a potent inducer of systemic acquired resistance (SAR) in tobacco and generates a wave of salicylic acid expanding from the site of application. Mol. Plant. Pathol. 2013, 14, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Windram, O.; Madhou, P.; McHattie, S.; Hill, C.; Hickman, R.; Cooke, E.; Jenkins, D.J.; Penfold, C.A.; Baxter, L.; Breeze, E.; et al. Arabidopsis defense against Botrytis cinerea: Chronology and regulation deciphered by high-resolution temporal transcriptomic analysis. Plant. Cell 2012, 24, 3530–3557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poland, C.A.; Duffin, R.; Kinloch, I.; Maynard, A.; Wallace, W.A.; Seaton, A.; Stone, V.; Brown, S.; MacNee, W.; Donaldson, K. Carbon nanotubes introduced into the abdominal cavity of mice show asbestos-like pathogenicity in a pilot study. Nat. Nanotechnol. 2008, 3, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Turóczy, Z.; Kis, P.; Török, K.; Cserháti, M.; Lendvai, Á.; Dudits, D.; Horváth, G.V. Overproduction of a rice aldo-keto reductase increases oxidative and heat stress tolerance by malondialdehyde and methylglyoxal detoxification. Plant. Mol. Biol. 2011, 75, 399–412. [Google Scholar] [PubMed]
- Gullner, G.; Komives, T.; Király, L.; Schröder, P. Glutathione S-transferase enzymes in plant-pathogen interactions. Front. Plant Sci. 2018, 9, 1836. [Google Scholar] [CrossRef] [Green Version]
- Dean, J.D.; Goodwin, P.H.; Hsiang, T. Induction of glutathione S-transferase genes of Nicotiana benthamiana following infection by Colletotrichum destructivum and C. orbiculare and involvement of one in resistance. J. Exp. Bot. 2005, 56, 1525–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, P.J.; Tantitadapitak, C.; Reed, A.M.; Mather, O.C.; Bunce, C.M.; White, S.A.; Ride, J.P. Characterization of two novel aldo—keto reductases from Arabidopsis: Expression patterns, broad substrate specificity, and an open active-site structure suggest a role in toxicant metabolism following stress. J. Mol. Biol. 2009, 392, 465–480. [Google Scholar] [CrossRef]
- Kanayama, Y.; Mizutani, R.; Yaguchi, S.; Hojo, A.; Ikeda, H.; Nishiyama, M.; Kanahama, K. Characterization of an uncharacterized aldo-keto reductase gene from peach and its role in abiotic stress tolerance. Phytochemistry 2014, 104, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, D.; Naik, D.; Reddy, A.R. Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plant metabolic processes and stress defense: A structure-function update. J. Plant. Physiol. 2015, 179, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Vemanna, R.S.; Vennapusa, A.R.; Easwaran, M.; Chandrashekar, B.K.; Rao, H.; Ghanti, K.; Sudhakar, C.; Mysore, K.S.; Makarla, U. Aldo-keto reductase enzymes detoxify glyphosate and improve herbicide resistance in plants. Plant. Biotechnol. J. 2017, 15, 794–804. [Google Scholar] [CrossRef]
- Wang, H.; He, Z.; Luo, L.; Zhao, X.; Lu, Z.; Luo, T.; Li, M.; Zhang, Y. An aldo-keto reductase, Bbakr1, is involved in stress response and detoxification of heavy metal chromium but not required for virulence in the insect fungal pathogen, Beauveria bassiana. Fungal Genet. Biol. 2018, 111, 7–15. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | SNP Name | Primer Name | Sequence (5′→3′) | Tm | PCR Product Size |
|---|---|---|---|---|---|
| 1 | SNP-1 | SNP 1 F | CTTTGAACTTCGGGCAATACCCG | 60.5 | 199 bp |
| SNP 1 R | CCTCATCAGGCGAGTGAGTGGAC | 59.6 | |||
| 2 | SNP-2 | SNP 2 F | AACGTCCGCCGAAGAAGCTGA | 60.7 | 204 bp |
| SNP 2 R | TTTGCTGGAGGAGGTGGTGGTG | 60.1 | |||
| 3 | SNP-3 | SNP 3 F | CGTTAGCTCAAGTGGGTTTGAGGTG | 59.9 | 134 bp |
| SNP 3 R | TTCTTCCAGCTCTTCTTCGCT | 59.2 | |||
| 4 | SNP-4 | SNP 4 F | AGGGTTCAGAACCAAAACAGCATCA | 59.8 | 163 bp |
| SNP 4 R | CGATGCTTTTTTGGTAACTGGGAAG | 59.1 | |||
| 5 | SNP-5 | SNP 5 F | TCGATGGCATTAAGGATGCTAAGGA | 59.8 | 163 bp |
| SNP 5 R | ATTGCCTTTGCTAGGGAGCCATAA | 59.1 | |||
| 6 | SNP-6 | SNP 6 F | TGGTGACAAGAAATTCTTCAACGGC | 60.2 | 158 bp |
| SNP 6 R | TCTCCATGCATCTCTTTCCCCACT | 59.8 | |||
| 7 | SNP-7 | SNP 7 F | TGAGCTCCTTTCAGACTCCTTTCCC | 60.1 | 170 bp |
| SNP 7 R | CGACCACCTTAACAGCTTGATCGTC | 59.9 | |||
| 8 | SNP-8 | SNP 8 F | CTTTCTCAGGGTTAATAGAGGCGGG | 60.1 | 171 bp |
| SNP 8 R | GCCAAACTGGCTGAAAACCTTTTCT | 59.9 | |||
| 9 | SNP-9 | SNP 9 F | TTCATGGTCACAGAAACGCCAAGA | 60.1 | 181 bp |
| SNP 9 R | GGCAGAACTTCTTTGTTCATCCGCT | 59.9 | |||
| 10 | SNP-10 | SNP 10 F | AATCTCACAATCGAACCTCACTGCC | 59.4 | 175 bp |
| SNP 10 R | TGCGAGGTGAATTCCAGTCAAAGAG | 60.4 | |||
| 11 | SNP-11 | SNP 11 F | GCAACAAGGGCTGCAAATTACAGTT | 60.1 | 169 bp |
| SNP 11 R | GTTTGTGTGCATGAATCTGTGCAGG | 59.9 | |||
| 12 | SNP-12 | SNP 12 F | CGACTATGGCTGGGACACTGCA | 59.6 | 164 bp |
| SNP 12 R | TCCCCGAACTTGACCCCGTTAC | 60.7 | |||
| 13 | SNP-13 | SNP 13 F | AGCAATGTTGTCCGGTACTCCAAAG | 59.4 | 185 bp |
| SNP 13 R | CGCTCAAAAACCCAGCTCGTACA | 60.1 | |||
| 14 | SNP-14 | SNP 14 F | TATCGTACCTTCCTACCCTGAGCGA | 60.1 | 182 bp |
| SNP 14 R | TCCGAACATGGGCAGCTTCC | 59.9 |
| No | Transcripts No | TAIR ID z | Gene Function |
|---|---|---|---|
| 1 | SNP-1 transcript | AT1G17420 | Lipoxygenase 3 |
| 2 | SNP-2 transcript | AT1G42970 | Glyceraldehyde-3-phosphate dehydrogenase B subunit |
| 3 | SNP-3 transcript | AT1G59960 | Aldo/keto reductase |
| 4 | SNP-4 transcript | AT1G67090 | Ribulose bisphosphate carboxylase small chain 1A |
| 5 | SNP-5 transcript | AT1G68090 | Annexin Arabidopsis thaliana 5 |
| 6 | SNP-6 transcript | AT1G78380 | Glutathione S-transferase |
| 7 | SNP-7 transcript | AT3G16640 | Translationally controlled tumor protein |
| 8 | SNP-8 transcript | AT4G39260 | Glycine-rich protein |
| 9 | SNP-9 transcript | AT5G13750 | Zinc induced facilitator-like 1 |
| 10 | SNP-10 transcript | AT5G52810 | Systemic acquired resistance |
| 11 | SNP-11 transcript | AT1G14290 | Sphingoid base hydroxylase2 |
| 12 | SNP-12 transcript | AT1G29930 | Chlorophyll A/B protein |
| 13 | SNP-13 transcript | AT5G25220 | KNAT3 |
| 14 | SNP-14 transcript | AT5G38430 | Rubisco small subunit 1B |
| (A) HRM analysis results of the ‘Seeds & People’ Co. onion lines. | ||||
| HRM Marker | Line | LS Means | Group * | |
| SNP-3 | S&P 7522 | 79.642 | A | |
| S&P 7521 | 79.768 | A | ||
| S&P 7129 | 79.782 | A | ||
| S&P 7168 | 79.838 | A | ||
| S&P 7483 | 80.388 | B | ||
| S&P 7130 | 80.353 | B | ||
| S&P 7175 | 80.472 | B | ||
| (B) HRM analysis results of the ‘Asia seed’ Co. onion lines. | ||||
| HRM Marker | Line | LS Means | Group * | |
| SNP-3 | Asia-12 | 82.183 | A | |
| Asia-53 | 82.217 | A | ||
| Asia-48 | 82.253 | A | ||
| Asia-50 | 82.473 | B | ||
| Asia-35 | 82.500 | B | ||
| Asia-45 | 82.527 | B | ||
| Asia-30 | 82.550 | B | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-J.; Park, J.-S.; Park, T.; Lee, H.-M.; Choi, J.-R.; Park, Y.-D. Development of Molecular Markers Associated with Resistance to Gray Mold Disease in Onion (Allium cepa L.) through RAPD-PCR and Transcriptome Analysis. Horticulturae 2021, 7, 436. https://doi.org/10.3390/horticulturae7110436
Kim S-J, Park J-S, Park T, Lee H-M, Choi J-R, Park Y-D. Development of Molecular Markers Associated with Resistance to Gray Mold Disease in Onion (Allium cepa L.) through RAPD-PCR and Transcriptome Analysis. Horticulturae. 2021; 7(11):436. https://doi.org/10.3390/horticulturae7110436
Chicago/Turabian StyleKim, So-Jeong, Jee-Soo Park, TaeHoon Park, Hyun-Min Lee, Ju-Ri Choi, and Young-Doo Park. 2021. "Development of Molecular Markers Associated with Resistance to Gray Mold Disease in Onion (Allium cepa L.) through RAPD-PCR and Transcriptome Analysis" Horticulturae 7, no. 11: 436. https://doi.org/10.3390/horticulturae7110436
APA StyleKim, S. -J., Park, J. -S., Park, T., Lee, H. -M., Choi, J. -R., & Park, Y. -D. (2021). Development of Molecular Markers Associated with Resistance to Gray Mold Disease in Onion (Allium cepa L.) through RAPD-PCR and Transcriptome Analysis. Horticulturae, 7(11), 436. https://doi.org/10.3390/horticulturae7110436