
Stability of Dry and Liquid Metschnikowia pulcherrima Formulations for Biocontrol Applications against Apple Postharvest Diseases

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Cultivation
2.2. Dry Formulation of M. pulcherrima
2.3. Liquid Formulation of M. pulcherrima
2.4. CFU Determination
2.5. Competition Assay with Stored Cells
2.6. Growth Assays in YNB Supplemented with Xanthan
2.7. In Situ Competition Assays on Apple Fruit
2.8. Field Trial, Harvest, Storage and Evaluation
2.9. Statistical Analyses
3. Results
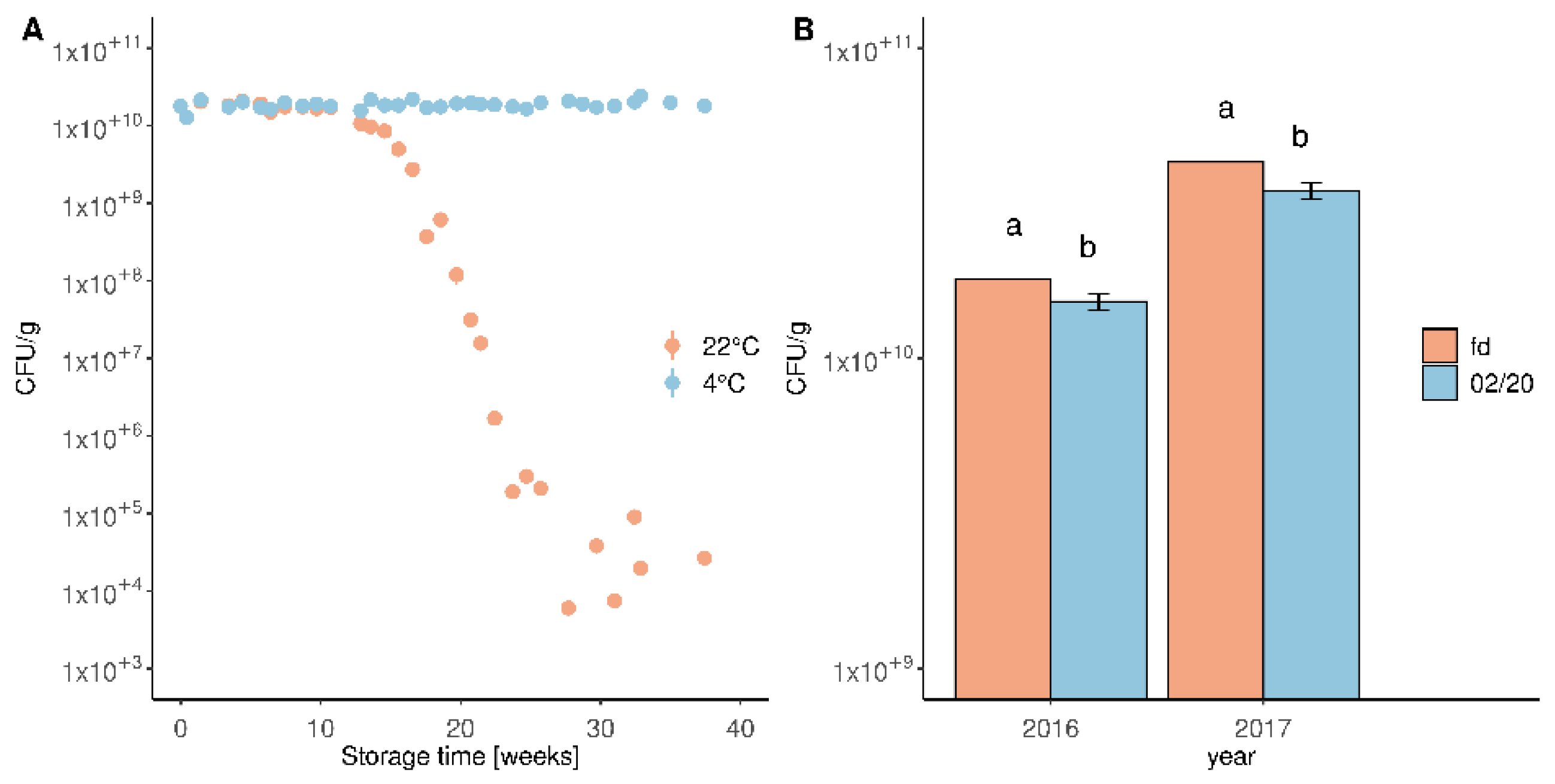
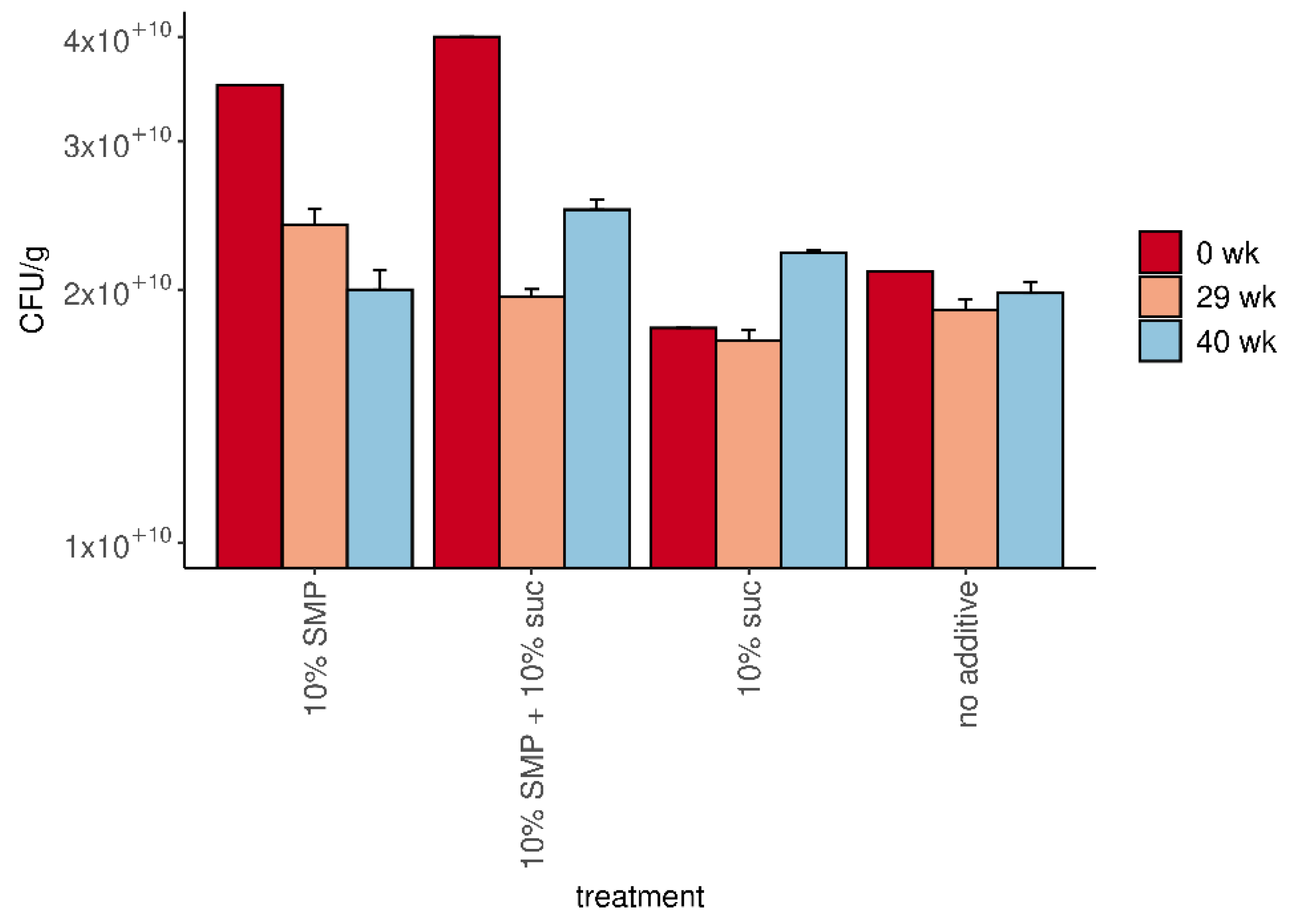
3.1. Freeze-Dried Formulation of M. pulcherrima Retains Cell Viability over Several Months
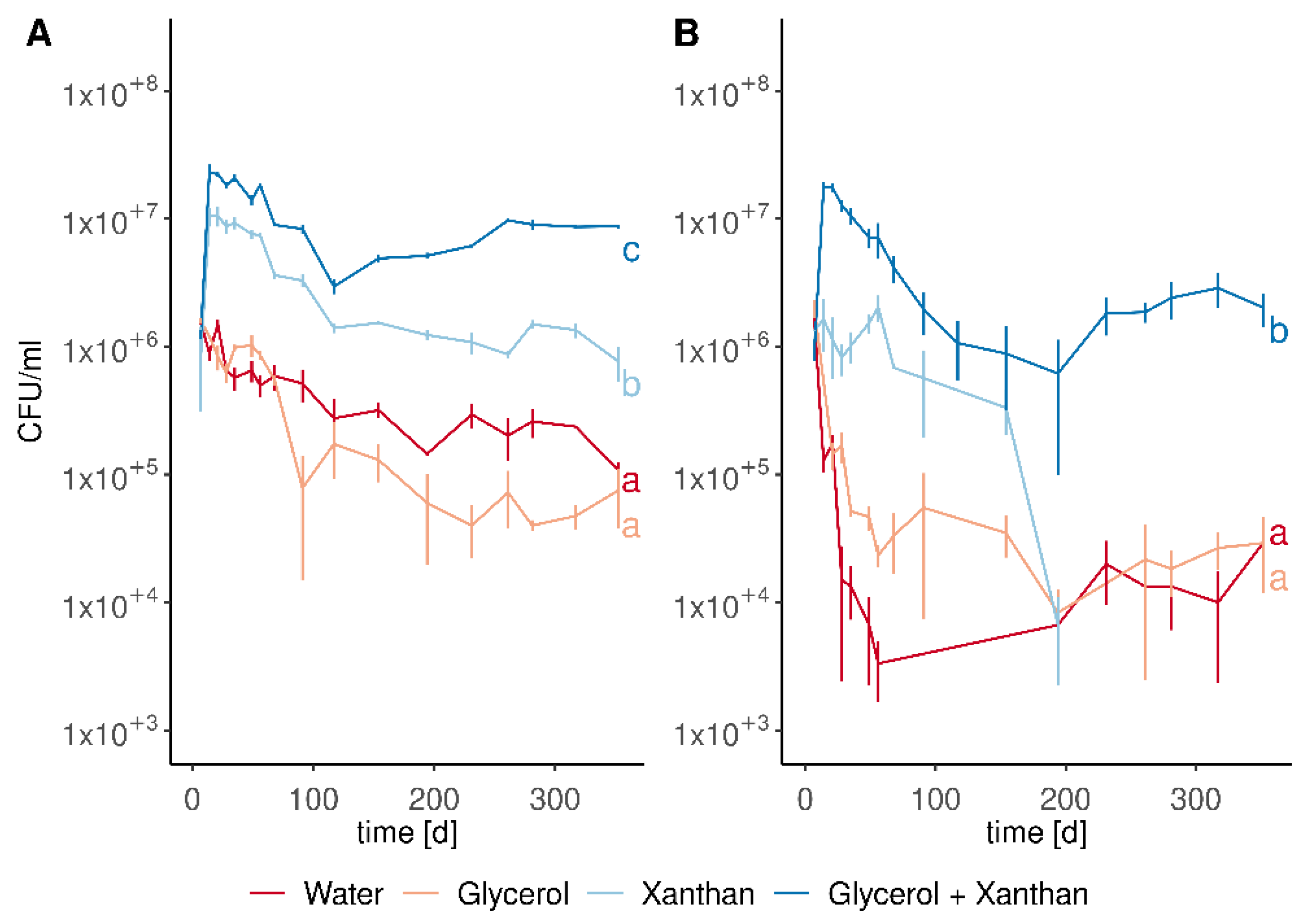
3.2. Liquid Formulations of M. pulcherrima Remain Viable for Several Months
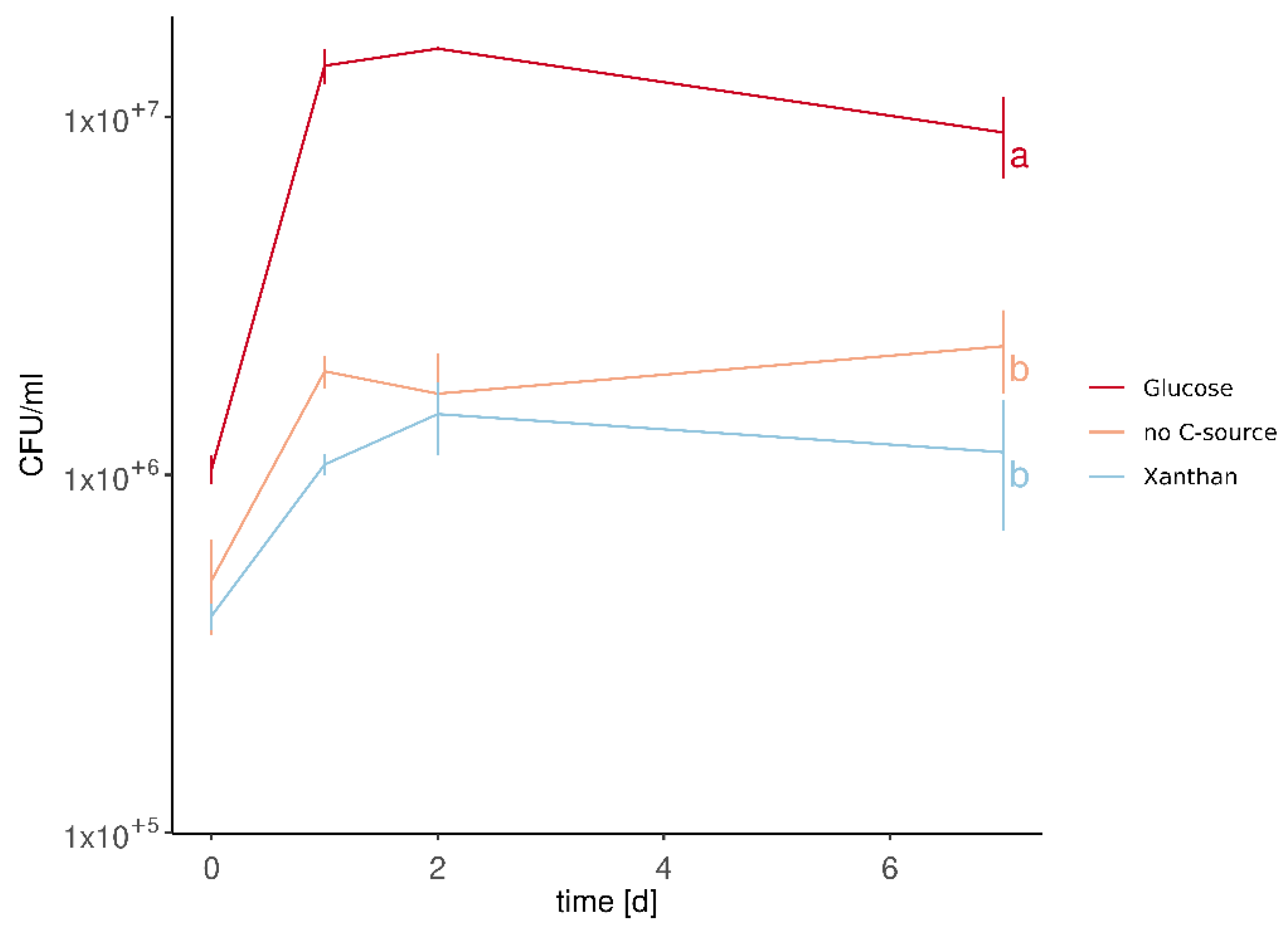
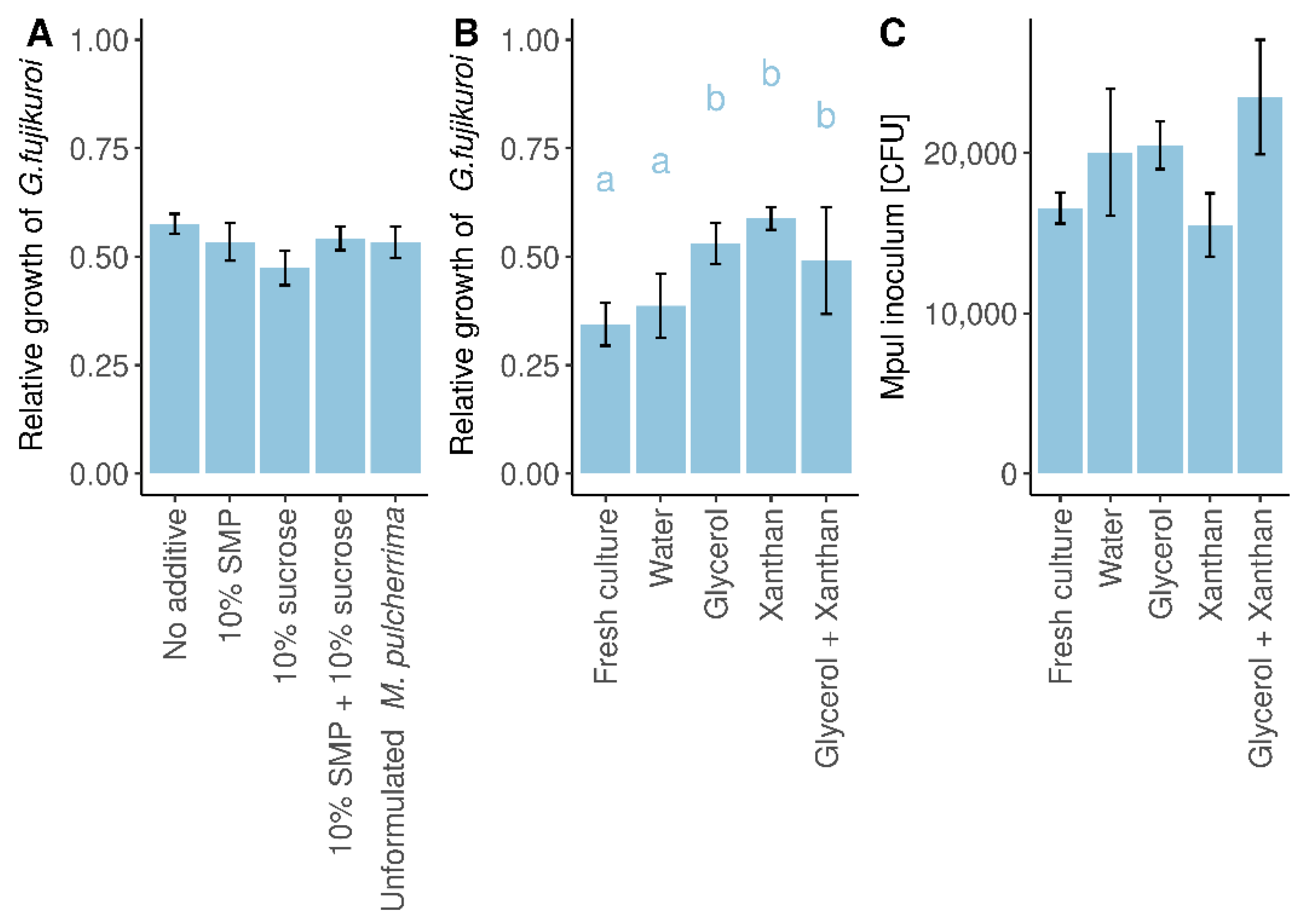
3.3. M. pulcherrima Is Not Able to Metabolise Xanthan
3.4. Formulated M. pulcherrima Cells Retain Their Antagonistic Activity against G. fujikuroi
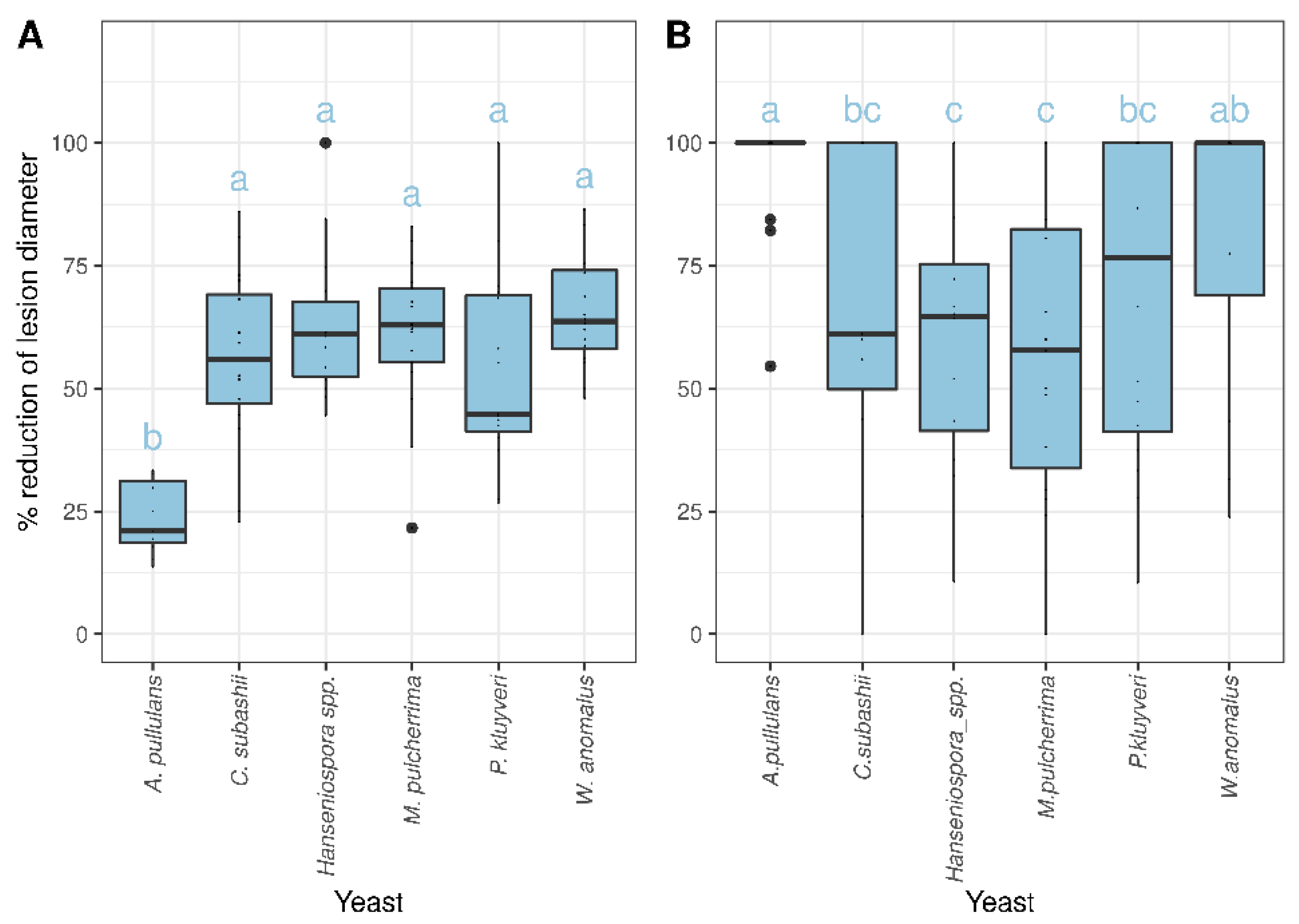
3.5. In Situ Competition Assays on Apple Fruit
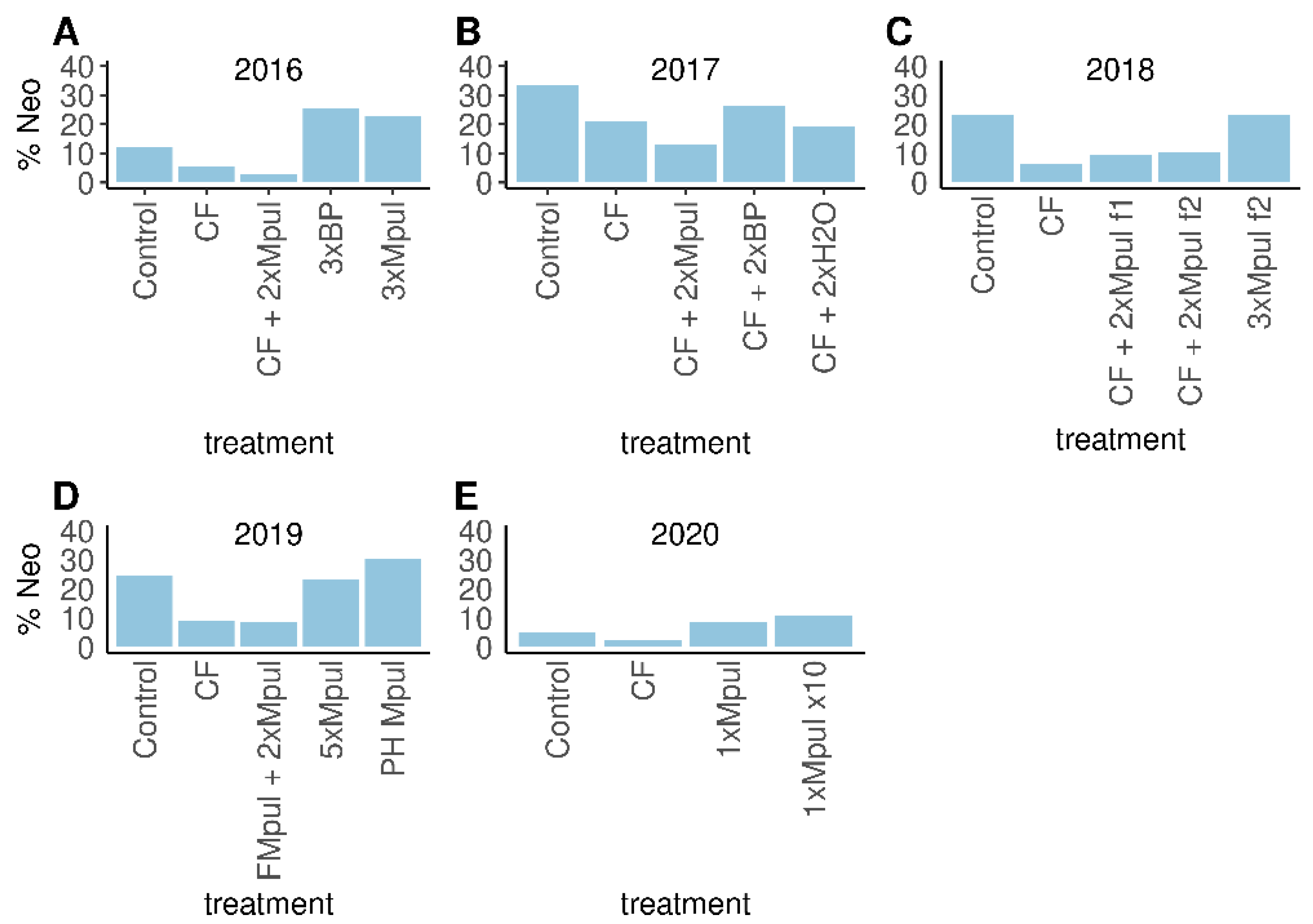
3.6. Field Trial Data
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Freimoser, F.M.; Rueda-Mejia, M.P.; Tilocca, B.; Migheli, Q. Biocontrol Yeasts: Mechanisms and Applications. World J. Microbiol. Biotechnol. 2019, 35, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gore-Lloyd, D.; Sumann, I.; Brachmann, A.O.; Schneeberger, K.; Ortiz-Merino, R.A.; Moreno-Beltrán, M.; Schläfli, M.; Kirner, P.; Santos Kron, A.; Rueda-Mejia, M.P.; et al. Snf2 Controls Pulcherriminic Acid Biosynthesis and Antifungal Activity of the Biocontrol Yeast Metschnikowia pulcherrima. Mol. Microbiol. 2019, 112, 317–332. [Google Scholar] [CrossRef] [Green Version]
- Vadkertiova, R.; Molnarova, J.; Vranova, D.; Slavikova, E. Yeasts and Yeast-like Organisms Associated with Fruits and Blossoms of Different Fruit Trees. Can. J. Microbiol. 2012, 58, 1344–1352. [Google Scholar] [CrossRef] [PubMed]
- Hilber-Bodmer, M.; Schmid, M.; Ahrens, C.H.; Freimoser, F.M. Competition Assays and Physiological Experiments of Soil and Phyllosphere Yeasts Identify Candida subhashii as a Novel Antagonist of Filamentous Fungi. BMC Microbiol. 2017, 17, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Droby, S.; El-Gerberia, B. Yeast Metschnikowia Fructicola NRRL Y-30752 for Inhibiting Deleterious Microorganisms on Plants. U.S. Patent No. 6,994,849, 2 February 2006. [Google Scholar]
- Pawlikowska, E.; James, S.A.; Breierova, E.; Antolak, H.; Kregiel, D. Biocontrol Capability of Local Metschnikowia sp. Isolates. Antonie Van Leeuwenhoek 2019, 112, 1425–1445. [Google Scholar] [CrossRef] [Green Version]
- Turkel, S.; Korukluoglu, M.; Yavuz, M. Biocontrol Activity of the Local Strain of Metschnikowia pulcherrima on Different Postharvest Pathogens. Biotechnol. Res. Int. 2014, 2014, 397167. [Google Scholar] [CrossRef] [Green Version]
- Turkel, S.; Ener, B. Isolation and Characterization of New Metschnikowia pulcherrima Strains as Producers of the Antimicrobial Pigment Pulcherrimin. Z. Naturforsch. C 2009, 64, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Sipiczki, M. Metschnikowia Strains Isolated from Botrytized Grapes Antagonize Fungal and Bacterial Growth by Iron Depletion. Appl. Environ. Microbiol. 2006, 72, 6716–6724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras, A.; Curtin, C.; Varela, C. Yeast Population Dynamics Reveal a Potential “collaboration” between Metschnikowia pulcherrima and Saccharomyces uvarum for the Production of Reduced Alcohol Wines during Shiraz Fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 1885–1895. [Google Scholar] [CrossRef] [PubMed]
- Contreras, A.; Hidalgo, C.; Henschke, P.A.; Chambers, P.J.; Curtin, C.; Varela, C. Evaluation of Non-Saccharomyces Yeasts for the Reduction of Alcohol Content in Wine. Appl. Environ. Microbiol. 2014, 80, 1670–1678. [Google Scholar] [CrossRef] [Green Version]
- Chantasuban, T.; Santomauro, F.; Gore-Lloyd, D.; Parsons, S.; Henk, D.; Scott Roderick, J.; Chuck, C. Elevated Production of the Aromatic Fragrance Molecule, 2-phenylethanol, Using Metschnikowia pulcherrima through Both de Novo and Ex Novo Conversion in Batch and Continuous Modes. J. Chem. Technol. Biotechnol. 2018, 93, 2118–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santamauro, F.; Whiffin, F.M.; Scott, R.J.; Chuck, C.J. Low-Cost Lipid Production by an Oleaginous Yeast Cultured in Non-Sterile Conditions Using Model Waste Resources. Biotechnol. Biofuels 2014, 7, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Cruz Quiroz, R.; Cruz Maldonado, J.J.; Rostro Alanis, M.D.J.; Torres, J.A.; Parra Saldívar, R. Fungi-Based Biopesticides: Shelf-Life Preservation Technologies Used in Commercial Products. J. Pest Sci. 2019, 92, 1003–1015. [Google Scholar] [CrossRef]
- Chaudhary, T.; Dixit, M.; Gera, R.; Shukla, A.K.; Prakash, A.; Gupta, G.; Shukla, P. Techniques for Improving Formulations of Bioinoculants. 3 Biotech 2020, 10, 199. [Google Scholar] [CrossRef] [PubMed]
- Marian, M.; Shimizu, M. Improving Performance of Microbial Biocontrol Agents against Plant Diseases. J. Gen. Plant Pathol. 2019, 85, 329–336. [Google Scholar] [CrossRef]
- Rao, M.S.; Umamaheswari, R.; Chakravarthy, A.K.; Grace, G.N.; Kamalnath, M.; Prabu, P. A Frontier Area of Research on Liquid Biopesticides: The Way Forward for Sustainable Agriculture in India. Curr. Sci. 2015, 108, 3. [Google Scholar]
- Gasic, S.; Tanovic, B. Biopesticide Formulations, Possibility of Application and Future Trends. Pestic. Fitomedicina 2013, 28, 97–102. [Google Scholar] [CrossRef]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.-P. Advances in Plant Growth-Promoting Bacterial Inoculant Technology: Formulations and Practical Perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Bashir, O.; Claverie, J.P.; Lemoyne, P.; Vincent, C. Controlled-Release of Bacillus thurigiensis Formulations Encapsulated in Light-Resistant Colloidosomal Microcapsules for the Management of Lepidopteran Pests of Brassica Crops. PeerJ 2016, 4, e2524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, H.; Xi, N.; Zhang, Y. Microemulsion Formulation of a New Biopesticide to Control the Diamondback Moth (Lepidoptera: Plutellidae). Sci. Rep. 2018, 8, 10565. [Google Scholar] [CrossRef] [Green Version]
- Yaakov, N.; Ananth Mani, K.; Felfbaum, R.; Lahat, M.; Da Costa, N.; Belausov, E.; Ment, D.; Mechrez, G. Single Cell Encapsulation via Pickering Emulsion for Biopesticide Applications. ACS Omega 2018, 3, 14294–14301. [Google Scholar] [CrossRef]
- Yang, Y.; Fang, Z.; Chen, X.; Zhang, W.; Xie, Y.; Chen, Y.; Liu, Z.; Yuan, W. An Overview of Pickering Emulsions: Solid-Particle Materials, Classification, Morphology, and Applications. Front. Pharmacol. 2017, 8, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berninger, T.; López, Ó.G.; Bejarano, A.; Preininger, C.; Sessitsch, A. Maintenance and Assessment of Cell Viability in Formulation of Non-Sporulating Bacterial Inoculants. Microb. Biotechnol. 2018, 11, 277–301. [Google Scholar] [CrossRef] [Green Version]
- Abadias, M.; Teixido, N.; Usall, J.; Benabarre, A.; Vinas, I. Viability, Efficacy, and Storage Stability of Freeze-Dried Biocontrol Agent Candida sake Using Different Protective and Rehydration Media. J. Food Prot. 2001, 64, 856–861. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Güitrón, L.; Calderón-Santoyo, M.; Ortiz-Basurto, R.I.; Bautista-Rosales, P.U.; Ragazzo-Sánchez, J.A. Optimisation of the Spray Drying Process of Formulating the Post-Harvest Biocontrol Agent Meyerozyma caribbica. Biocontrol Sci. Technol. 2018, 28, 574–590. [Google Scholar] [CrossRef]
- Kunz, S. Development of “Blossom-Protect”—A Yeast Preparation for the Reduction of Blossom Infections by Fire Blight. In Proceedings of the 11th International Conference on Cultivation Technique and Phytopathological Problems in Organic Fruit-Growing, Weinsberg, Germany, 3–5 February 2004; Boos, M., Ed.; Fördergemeinschaft Ökologischer Obstbau—FÖKO: Weinsberg, Germany, 2004. [Google Scholar]
- Droby, S.; Cohen, L.; Daus, A.; Weiss, B.; Horev, B.; Chalutz, E.; Katz, H.; Keren-Tzur, M.; Shachnai, A. Commercial Testing of Aspire: A Yeast Preparation for the Biological Control of Postharvest Decay of Citrus. Biol. Control 1998, 12, 97–101. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glockner, F.O.; Tedersoo, L.; et al. The UNITE Database for Molecular Identification of Fungi: Handling Dark Taxa and Parallel Taxonomic Classifications. Nucleic Acids Res. 2018, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- Rueda-Mejia, M.P.; Nägeli, L.; Lutz, S.; Hayes, R.D.; Varadarajan, A.R.; Grigoriev, I.V.; Ahrens, C.H.; Freimoser, F.M. Genome, Transcriptome and Secretome Analyses of the Antagonistic, Yeast-like Fungus Aureobasidium pullulans to Identify Potential Biocontrol Genes. Microb Cell 2021, 8, 184–202. [Google Scholar] [CrossRef] [PubMed]
- Magoye, E.; Hilber-Bodmer, M.; Pfister, M.; Freimoser, F.M. Unconventional Yeasts Are Tolerant to Common Antifungals, and Aureobasidium pullulans Has Low Baseline Sensitivity to Captan, Cyprodinil, and Difenoconazole. Antibiotics 2020, 9, 602. [Google Scholar] [CrossRef]
- Köhl, J.; Wenneker, M.; Haas, B.H.G.; Anbergen, R.; de Geijn, H.M.G.; der Plas, C.H.L.; Pinto, F.A.M.F.; Kastelein, P. Dynamics of Post-Harvest Pathogens Neofabraea spp. and Cadophora spp. in Plant Residues in Dutch Apple and Pear Orchards. Plant Pathol. 2018, 67, 1264–1277. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, V.; Radecka, D.; Aerts, G.; Verstrepen, K.J.; Lievens, B.; Thevelein, J.M. Phenotypic Landscape of Non-Conventional Yeast Species for Different Stress Tolerance Traits Desirable in Bioethanol Fermentation. Biotechnol. Biofuels 2017, 10, 216. [Google Scholar] [CrossRef]
- Miyamoto-Shinohara, Y.; Sukenobe, J.; Imaizumi, T.; Nakahara, T. Survival Curves for Microbial Species Stored by Freeze-Drying. Cryobiology 2006, 52, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Bond, C. Freeze-Drying of Yeast Cultures. Methods Mol. Biol. 2007, 368, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Melin, P.; Hakansson, S.; Eberhard, T.H.; Schnurer, J. Survival of the Biocontrol Yeast Pichia anomala after Long-Term Storage in Liquid Formulations at Different Temperatures, Assessed by Flow Cytometry. J. Appl. Microbiol. 2006, 100, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Mokhtarnejad, L.; Etebarian, H.R.; Fazeli, M.R.; Jamalifar, H. Evaluation of Different Formulations of Potential Biocontrol Yeast Isolates Efficacy on Apple Blue Mold at Storage Condition. Arch. Phytopathol. Plant Prot. 2011, 40, 970–980. [Google Scholar] [CrossRef]
- Patino-Vera, M.; Jimenez, B.; Balderas, K.; Ortiz, M.; Allende, R.; Carrillo, A.; Galindo, E. Pilot-Scale Production and Liquid Formulation of Rhodotorula minuta, a Potential Biocontrol Agent of Mango Anthracnose. J. Appl. Microbiol. 2005, 99, 540–550. [Google Scholar] [CrossRef]
- Spadaro, D.; Ciavorella, A.A.; Lopez-Reyes, J.G.; Garibaldi, A.; Gullino, M.L. Effect of Culture Age, Protectants, and Initial Cell Concentration on Viability of Freeze-Dried Cells of Metschnikowia pulcherrima. Can. J. Microbiol. 2010, 56, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto-Shinohara, Y.; Imaizumi, T.; Sukenobe, J.; Murakami, Y.; Kawamura, S.; Komatsu, Y. Survival Rate of Microbes after Freeze-Drying and Long-Term Storage. Cryobiology 2000, 41, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Polomska, X.; Wojtatowicz, M.; Zarowska, B.; Szołtysik, M.; Chrzanowska, J. Freeze-Drying Preservation of Yeast Adjunct Cultures for Cheese Production. Pol. J. Food Nutr. Sci. 2012, 62, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Melin, P.; Hakansson, S.; Schnurer, J. Optimisation and Comparison of Liquid and Dry Formulations of the Biocontrol Yeast Pichia Anomala J121. Appl. Microbiol. Biotechnol. 2007, 73, 1008–1016. [Google Scholar] [CrossRef]
- Costa, E.; Usall, J.; Teixidó, N.; Garcia, N.; Viñas, I. Effect of Protective Agents, Rehydration Media and Initial Cell Concentration on Viability of Pantoea Agglomerans Strain CPA-2 Subjected to Freeze-Drying. J. Appl. Microbiol. 2000, 89, 793–800. [Google Scholar] [CrossRef]
- Chen, H.-C.; Lin, C.-W.; Chen, M.-J. The Effects of Freeze Drying and Rehydration on Survival of Microorganisms in Kefir. Asian-Australas. J. Anim. Sci. 2006, 19, 126–130. [Google Scholar] [CrossRef]
- Li, B.Q.; Tian, S.P. Effects of Trehalose on Stress Tolerance and Biocontrol Efficacy of Cryptococcus laurentii. J. Appl. Microbiol. 2006, 100, 854–861. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Lu, Z.; Yuan, Y.; Lü, F.; Bie, X. Optimization of a Protective Medium for Enhancing the Viability of Freeze-Dried Lactobacillus delbrueckii Subsp. Bulgaricus Based on Response Surface Methodology. J. Ind. Microbiol. Biotechnol. 2006, 33, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.; Weindling, E.; Rabinovich, E.; Nachman, I.; Fuchs, S.; Chuartzman, S.; Gal, L.; Schuldiner, M.; Bar-Nun, S. Water-Transfer Slows Aging in Saccharomyces Cerevisiae. PLoS ONE 2016, 11, e0148650. [Google Scholar] [CrossRef]
- Granot, D.; Snyder, M. Carbon Source Induces Growth of Stationary Phase Yeast Cells, Independent of Carbon Source Metabolism. Yeast 1993, 9, 465–479. [Google Scholar] [CrossRef]
- Longo, V.D.; Mattson, M.P. Fasting: Molecular Mechanisms and Clinical Applications. Cell Metab. 2014, 19, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Wei, M.; Fabrizio, P.; Hu, J.; Ge, H.; Cheng, C.; Li, L.; Longo, V.D. Life Span Extension by Calorie Restriction Depends on Rim15 and Transcription Factors Downstream of Ras/PKA, Tor, and Sch9. PLoS Genet. 2008, 4, e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrizio, P.; Hoon, S.; Shamalnasab, M.; Galbani, A.; Wei, M.; Giaever, G.; Nislow, C.; Longo, V.D. Genome-Wide Screen in Saccharomyces cerevisiae Identifies Vacuolar Protein Sorting, Autophagy, Biosynthetic, and TRNA Methylation Genes Involved in Life Span Regulation. PLoS Genet. 2010, 6, e1001024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aris, J.P.; Alvers, A.L.; Ferraiuolo, R.A.; Fishwick, L.K.; Hanvivatpong, A.; Hu, D.; Kirlew, C.; Leonard, M.T.; Losin, K.J.; Marraffini, M.; et al. Autophagy and Leucine Promote Chronological Longevity and Respiration Proficiency during Calorie Restriction in Yeast. Exp. Gerontol. 2013, 48, 1107–1119. [Google Scholar] [CrossRef] [Green Version]
- Burtner, C.R.; Murakami, C.J.; Kennedy, B.K.; Kaeberlein, M. A Molecular Mechanism of Chronological Aging in Yeast. Cell Cycle 2009, 8, 1256–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granot, D.; Snyder, M. Glucose Induces CAMP-Independent Growth-Related Changes in Stationary-Phase Cells of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1991, 88, 5724–5728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, S.; Kaplan, D.L. Biosynthesized Materials: Properties and Processing. In Encyclopedia of Materials: Science and Technology; Buschow, K.H.J., Cahn, R.W., Flemings, M.C., Ilschner, B., Kramer, E.J., Mahajan, S., Veyssière, P., Eds.; Elsevier: Oxford, UK, 2001; pp. 609–615. ISBN 978-0-08-043152-9. [Google Scholar]
- Williams, P.A.; Phillips, G.O. GUMS—Properties of Individual Gums. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 2992–3001. ISBN 978-0-12-227055-0. [Google Scholar]
- Kulkarni, V.S.; Shaw, C. Chapter 5—Use of Polymers and Thickeners in Semisolid and Liquid Formulations. In Essential Chemistry for Formulators of Semisolid and Liquid Dosages; Kulkarni, V.S., Shaw, C., Eds.; Academic Press: Boston, MA, USA, 2016; pp. 43–69. ISBN 978-0-12-801024-2. [Google Scholar]
- Morris, E.R. Mixed Polymer Gels. In Food Gels; Harris, P., Ed.; Elsevier Applied Food Science Series; Springer: Dordrecht, The Netherlands, 1990; pp. 291–359. ISBN 978-94-009-0755-3. [Google Scholar]
- Iijima, M.; Shinozaki, M.; Hatakeyama, T.; Takahashi, M.; Hatakeyama, H. AFM Studies on Gelation Mechanism of Xanthan Gum Hydrogels. Carbohydr. Polym. 2007, 68, 701–707. [Google Scholar] [CrossRef]
- Wang, C.-S.; Virgilio, N.; Wood-Adams, P.; Heuzey, M.-C. A Mechanism for the Synergistic Gelation Properties of Gelatin B and Xanthan Gum Aqueous Mixtures. Carbohydr. Polym. 2017, 175, 484–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, P.A.; Clegg, S.M.; Day, D.H.; Phillips, G.O.; Nishinari, K. Mixed Gels Formed with Konjac Mannan and Xanthan Gum. In Food Polymers, Gels and Colloids; Dickinson, E., Ed.; Woodhead Publishing: Sawston, UK, 1991; pp. 339–348. ISBN 978-1-85573-787-7. [Google Scholar]
- Williams, P.A.; Annable, P.; Phillips, G.O.; Nishinari, K. Mixed Polysaccharide Gels Formed between Xanthan Gum and Glucomannan. In Food Hydrocolloids: Structures, Properties, and Functions; Nishinari, K., Doi, E., Eds.; Springer: Boston, MA, USA, 1993; pp. 435–449. ISBN 978-1-4615-2486-1. [Google Scholar]
- Xue, D.; Sethi, R. Viscoelastic Gels of Guar and Xanthan Gum Mixtures Provide Long-Term Stabilization of Iron Micro- and Nanoparticles. J. Nanopart. Res. 2012, 14, 1239. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulation | CFU/g | Yield (g) | Survival (%) | Total Viable Cells |
|---|---|---|---|---|
| No additive | 2.1 × 1010 | 30.3 | 12.5 | 6.3 × 1011 |
| 10% sucrose | 1.8 × 1010 | 61.9 | 22.6 | 1.1 × 1012 |
| 10% SMP | 3.5 × 1010 | 60.8 | 43.1 | 2.1 × 1012 |
| 10% sucrose + 10% SMP | 4.0 × 1010 | 90.6 | 72.1 | 3.6 × 1012 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bühlmann, A.; Kammerecker, S.; Müller, L.; Hilber-Bodmer, M.; Perren, S.; Freimoser, F.M. Stability of Dry and Liquid Metschnikowia pulcherrima Formulations for Biocontrol Applications against Apple Postharvest Diseases. Horticulturae 2021, 7, 459. https://doi.org/10.3390/horticulturae7110459
Bühlmann A, Kammerecker S, Müller L, Hilber-Bodmer M, Perren S, Freimoser FM. Stability of Dry and Liquid Metschnikowia pulcherrima Formulations for Biocontrol Applications against Apple Postharvest Diseases. Horticulturae. 2021; 7(11):459. https://doi.org/10.3390/horticulturae7110459
Chicago/Turabian StyleBühlmann, Andreas, Sandrine Kammerecker, Laurin Müller, Maja Hilber-Bodmer, Sarah Perren, and Florian M. Freimoser. 2021. "Stability of Dry and Liquid Metschnikowia pulcherrima Formulations for Biocontrol Applications against Apple Postharvest Diseases" Horticulturae 7, no. 11: 459. https://doi.org/10.3390/horticulturae7110459
APA StyleBühlmann, A., Kammerecker, S., Müller, L., Hilber-Bodmer, M., Perren, S., & Freimoser, F. M. (2021). Stability of Dry and Liquid Metschnikowia pulcherrima Formulations for Biocontrol Applications against Apple Postharvest Diseases. Horticulturae, 7(11), 459. https://doi.org/10.3390/horticulturae7110459