Drought and Darkness during Long-Term Simulated Shipping Delay Post-Shipping Flowering of Phalaenopsis Sogo Yukidian ‘V3’


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
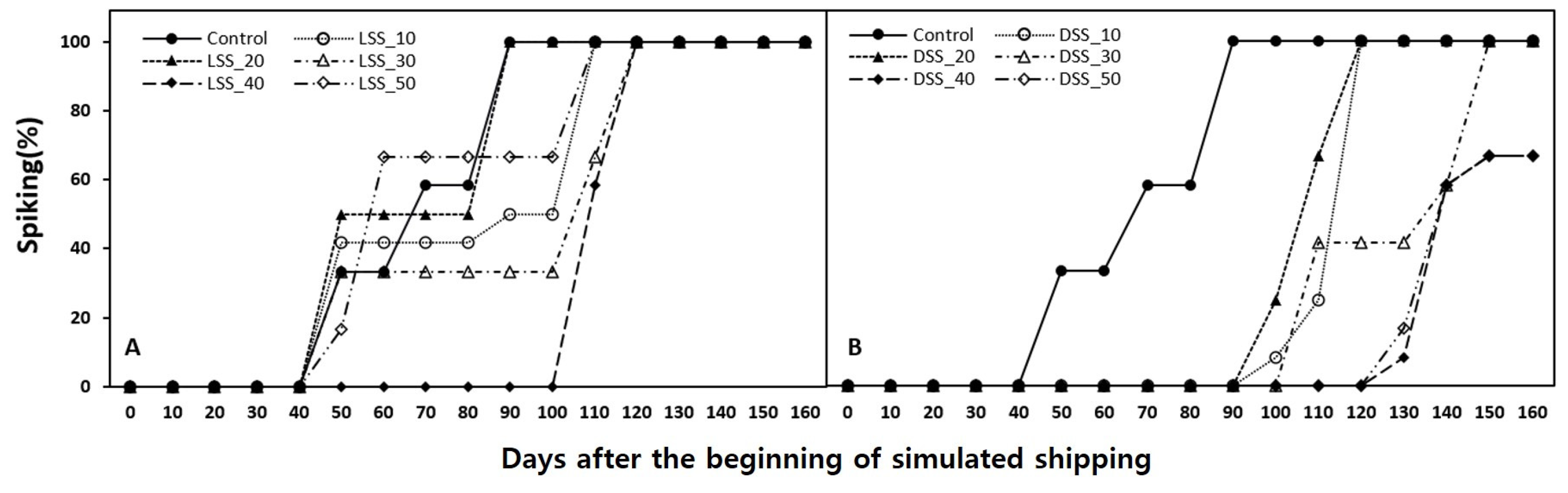
2.2. Simulated Shipping Conditions
2.3. Measurements of Photochemical Reflectance Index and Chlorophyll Fluorescence
2.4. Data Collection and Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lopez, R.; Runkle, E.; Wang, Y.T.; Blanchard, M.; Hsu, T. Growing the best Phalaenopsis. Part 3: Temperature and light requirements, height, insect and disease control. Orchids 2007, 76, 182–187. [Google Scholar]
- Min, S.Y.; Oh, W. Effects of nutrient solution application methods and rhizospheric ventilation on vegetative growth of young moth orchids without a potting medium in a closed-type plant factory. J. People Plants Environ. 2020, 23, 545–554. [Google Scholar] [CrossRef]
- Hew, C.S.; Yong, J.W.H. Physiology of Tropical Orchids in Relation to the Industry, 2nd ed.; World Scientific Publishing Company: Singapore, 2004. [Google Scholar]
- Lee, J.S. Stomatal opening mechanism of CAM plants. J. Plant Biol. 2010, 53, 19–23. [Google Scholar] [CrossRef] [Green Version]
- Chen, C. Evaluation of the effect of temperature on a stem elongation model of Phalaenopsis. Horticulturae 2019, 5, 76. [Google Scholar] [CrossRef] [Green Version]
- Runkle, E.; Wang, Y.T.; Blanchard, M.; Lopez, R. Growing the best Phalaenopsis. Part 1: An introduction to potted Phalaenopsis orchids. Orchids 2007, 76, 24–28. [Google Scholar]
- Rural Development Administration (RDA). A Manual of Environmental Management for Smart Greenhouses; RDA: Jeonju, Korea, 2018; pp. 223–245.
- Blanchard, M.G.; Runkle, E.S. Temperature during the day, but not during the night, controls flowering of Phalaenopsis orchids. J. Exp. Bot. 2006, 57, 4043–4049. [Google Scholar] [CrossRef] [Green Version]
- Ministry for Agriculture, Food and Rural Affairs (MAFRA). Annual Report of Floriculture Cultivation Statistics 2019; MAFRA: Sejong, Korea, 2020.
- Rural Development Administration (RDA). Study for Seedling Production and Distribution of Domestic Orchid Variety for Cymbidium and Phalaenopsis; RDA: Jeonju, Korea, 2016.
- Rural Development Administration (RDA). Orchid (Phalaenopsis) Export Guideline; RDA: Jeonju, Korea, 2018.
- Animal and Plant Quarantine Agency (APQA). U.S. Export Quarantine Guidelines for Korean Cymbidium and Phalaenopsis Young Plants Planted in Growing Media; APQA: Gimcheon, Korea, 2017.
- Animal and Plant Health Inspection Service (APHIS). Importation of Moth Orchids (Phalaenopsis spp.) in Approved Growing Media from Taiwan; UDSA APHIS: Riverdale, MD, USA, 2003.
- Blanchard, M.; Lopez, R.; Runkle, E.; Wang, Y.T. Growing the best Phalaenopsis. Part 4: A complete production schedule. Orchids 2007, 76, 266–271. [Google Scholar]
- Chen, W.H.; Wang, Y.T. Phalaenopsis orchid culture. Taiwan Sugar 1996, 43, 11–16. [Google Scholar]
- Hou, J.Y.; Setter, T.L.; Chang, Y.C.A. Effects of simulated dark shipping on photosynthetic status and post-shipping performance in Phalaenopsis Sogo Yukidian ‘V3’. J. Am. Soc. Hortic. Sci. 2010, 135, 183–190. [Google Scholar] [CrossRef] [Green Version]
- National Institute of Horticultural and Herbal Science (NIHHS). The Optimal Condition of Medium Drying Treatment for Long-Distance Ship Export of Phalaenopsis Orchids. Available online: http://www.nongsaro.go.kr/portal/ps/psb/psbb/farmUseTechDtl.ps?sTchnlgyRealmCode=&sKidofcomdtySeCode=&sTchnlgyPrcuseTyCode=&sRsrchRealmCode=&sRealmName=&sPsitnCode=&sPsitnArea=&sDtlType=&pageIndex=1&noSpr=&menuId=PS00072&farmPrcuseSeqNo=100000157500&sType=sj&sReSearchType=sj&sValue=%ED%8C%94%EB%A0%88%EB%86%89%EC%8B%9C%EC%8A%A4&sReSearchValue=&_sReSearchYn=on (accessed on 14 July 2021).
- Bastide, B.; Sipes, D.; Hann, J.; Ting, I.P. Effect of severe water stress on aspects of Crassulacean acid metabolism in Xerosicyos. Plant Physiol. 1993, 103, 1089–1096. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.Y.; Pandey, D.M.; Hahn, E.J.; Paek, K.Y. Effect of drought on physiological aspects of Crassulacean acid metabolism in Doritaenopsis. Plant Sci. 2004, 167, 1219–1226. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Li, T.; Tan, G.F.; Huang, Y.; Wang, F.; Xiong, A.S. Effects of dark treatment and regular light recovery on the growth characteristics and regulation of chlorophyll in water dropwort. Plant Growth Regul. 2018, 85, 293–303. [Google Scholar] [CrossRef]
- Rahdari, P.; Hosseini, S.M.; Tavakoli, S. The studying effect of drought stress on germination, proline, sugar, lipid, protein and chlorophyll content in purslane (Portulaca oleracea L.) leaves. J. Med. Plants Res. 2012, 6, 1539–1547. [Google Scholar]
- Yang, X.; Chen, X.; Ge, Q.; Li, B.; Tong, Y.; Zhang, A.; Li, Z.; Kuang, T.; Lu, C. Tolerance of photosynthesis to photoinhibition, high temperature and drought stress in flag leaves of wheat: A comparison between a hybridization line and its parents grown under field conditions. Plant Sci. 2006, 171, 389–397. [Google Scholar] [CrossRef]
- Thenot, F.; Méthy, M.; Winkel, T. The photochemical reflectance index (PRI) as a water-stress index. Int. J. Remote Sens. 2002, 23, 5135–5139. [Google Scholar] [CrossRef]
- Lee, M.C.; Chang, D.C.; Wu, C.W.; Wang, Y.T.; Chang, Y.S. Phalaenopsis efficiently acclimate to highlight environment through orchid mycorrhization. Sci. Hortic. 2014, 179, 184–190. [Google Scholar] [CrossRef]
- Bolhar-Nordenkampf, H.R.; Long, S.P.; Baker, N.R.; Oquist, G.; Schreiber, U.; Lechner, E.G. Chlorophyll fluorescence as a probe of the photosynthetic competence of leaves in the field: A review of current instrumentation. Funct. Ecol. 1989, 3, 497–514. [Google Scholar] [CrossRef]
- Hou, J.Y.; Miller, W.B.; Chang, Y.C.A. Effects of simulated dark shipping on the carbohydrate status and post-shipping performance of Phalaenopsis. J. Am. Soc. Hortic. Sci. 2011, 136, 364–371. [Google Scholar] [CrossRef]
- Zeiger, E. Light perception in guard cells. Plant Cell Environ. 1990, 13, 739–744. [Google Scholar] [CrossRef]
- Vitale, L.; Vitale, E.; Costanzo, G.; De Maio, A.; Arena, C. Photo-protective mechanisms and the role of poly (ADP-ribose) polymerase activity in a facultative CAM plant exposed to long-term water deprivation. Plants 2020, 9, 1192. [Google Scholar] [CrossRef]
- Ceusters, J.; Borland, A.M.; Godts, C.; Londers, E.; Croonenborghs, S.; Van Goethem, D.; De Proft, M.P. Crassulacean acid metabolism under severe light limitation: A matter of plasticity in the shadows? Exp. Bot. 2011, 62, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Shavrukov, Y.; Kurishbayev, A.; Jatayev, S.; Shvidchenko, V.; Zotova, L.; Koekemoer, F.; Groot, S.D.; Soole, K.; Langridge, P. Early flowering as a drought escape mechanism in plants: How can it aid wheat production? Front. Plant Sci. 2017, 8, 1950. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Wu, C.; Zhou, B. Drought stress induces flowering and enhances carbohydrate accumulation in Averrhoa carambola. Hortic. Plant J. 2017, 3, 60–66. [Google Scholar] [CrossRef]
- Mohr, P.G.; Cahill, D.M. Abscisic acid influences the susceptibility of Arabidopsis thaliana to Pseudomonas syringae pv. tomato and Peronospora parasitica. Funct. Plant Biol. 2003, 30, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Pourtau, N.; Marès, M.; Purdy, S.; Quentin, N.; Ruël, A.; Wingler, A. Interactions of abscisic acid and sugar signalling in the regulation of leaf senescence. Planta 2004, 219, 765–772. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Simulated Shipping Duration (d) | Light Condition | Uppermost Mature Leaf | No. of New Leaves | Spike Length (cm) z | 1st Floret or Bud y | No. of Flower Buds | Days to Flower Visible Bud | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Length (cm) | Width (cm) | Length (cm) | Width (cm) | |||||||||||
| 0 | 20.1 | 7.0 | 1.1 | 54.1 | ab x | 78.6 | a | 62.5 | a | 7.2 | a | 125 | c | |
| 10 | Light | 20.9 | 6.7 | 1.2 | 32.9 | cd | 33.2 | cd | 28.7 | bcd | 3.6 | bc | 150 | abc |
| Dark | 20.6 | 6.8 | 1.2 | 48.8 | ab | 11.8 | cd | 9.4 | cd | 2.0 | c | 168 | a | |
| 20 | Light | 20.4 | 7.2 | 1.0 | 58.2 | a | 42.7 | bc | 36.9 | abc | 5.7 | ab | 140 | bc |
| Dark | 19.7 | 6.9 | 0.8 | 53.4 | ab | 5.5 | d | 7.0 | d | 2.4 | c | 171 | a | |
| 30 | Light | 20.8 | 6.9 | 1.2 | 48.7 | abc | 38.0 | bcd | 32.0 | bcd | 3.3 | bc | 149 | abc |
| Dark | 20.5 | 6.7 | 0.8 | 38.0 | bcd | 15.1 | cd | 13.2 | cd | 3.5 | bc | 150 | abc | |
| 40 | Light | 20.6 | 7.0 | 0.9 | 44.8 | a–d | 5.4 | d | 12.6 | cd | 2.9 | c | 163 | ab |
| Dark | 21.4 | 7.0 | 0.6 | 29.0 | cd | 4.0 | d | 4.2 | d | 1.6 | c | 168 | a | |
| 50 | Light | 20.0 | 6.9 | 0.6 | 53.5 | ab | 69.6 | ab | 54.4 | ab | 6.9 | a | 127 | c |
| Dark | 21.6 | 6.7 | 0.6 | 35.6 | d | 2.2 | d | 2.9 | d | 1.4 | c | 173 | a | |
| Significance | ||||||||||||||
| Duration (A) | NS | NS | NS | * | ** | NS | * | NS | ||||||
| Light condition (B) | NS | NS | NS | * | *** | *** | *** | *** | ||||||
| A × B | NS | NS | NS | * | * | NS | * | NS | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, J.H.; Oh, W. Drought and Darkness during Long-Term Simulated Shipping Delay Post-Shipping Flowering of Phalaenopsis Sogo Yukidian ‘V3’. Horticulturae 2021, 7, 483. https://doi.org/10.3390/horticulturae7110483
Jeong JH, Oh W. Drought and Darkness during Long-Term Simulated Shipping Delay Post-Shipping Flowering of Phalaenopsis Sogo Yukidian ‘V3’. Horticulturae. 2021; 7(11):483. https://doi.org/10.3390/horticulturae7110483
Chicago/Turabian StyleJeong, Ju Hui, and Wook Oh. 2021. "Drought and Darkness during Long-Term Simulated Shipping Delay Post-Shipping Flowering of Phalaenopsis Sogo Yukidian ‘V3’" Horticulturae 7, no. 11: 483. https://doi.org/10.3390/horticulturae7110483
APA StyleJeong, J. H., & Oh, W. (2021). Drought and Darkness during Long-Term Simulated Shipping Delay Post-Shipping Flowering of Phalaenopsis Sogo Yukidian ‘V3’. Horticulturae, 7(11), 483. https://doi.org/10.3390/horticulturae7110483