
Key Proteins and Metabolic Pathways Involved in 24-Epibrasionlide Improving Drought Tolerance of Rhododendron delavayi Franch

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Determination of Instantaneous Gas Exchange and Physiological Traits
2.3. Protein Extraction and Digestion
2.4. High pH Reverse Phase Separation
2.5. Nano-HPLC-MS/MS Analysis
2.6. Data Analysis
3. Results
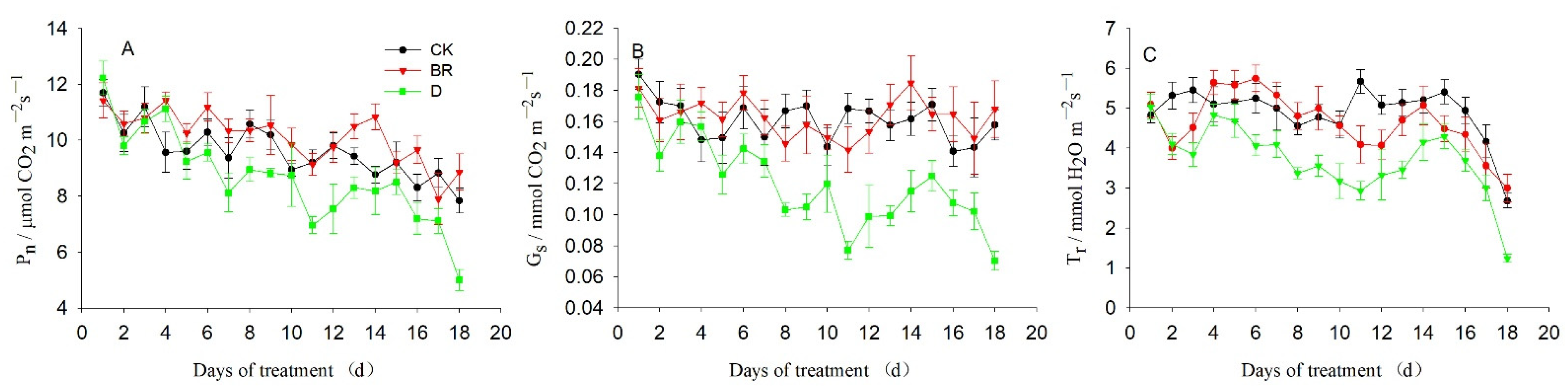
3.1. Effects of BR and Drought on the Photosynthetic Performance of R. delavayi
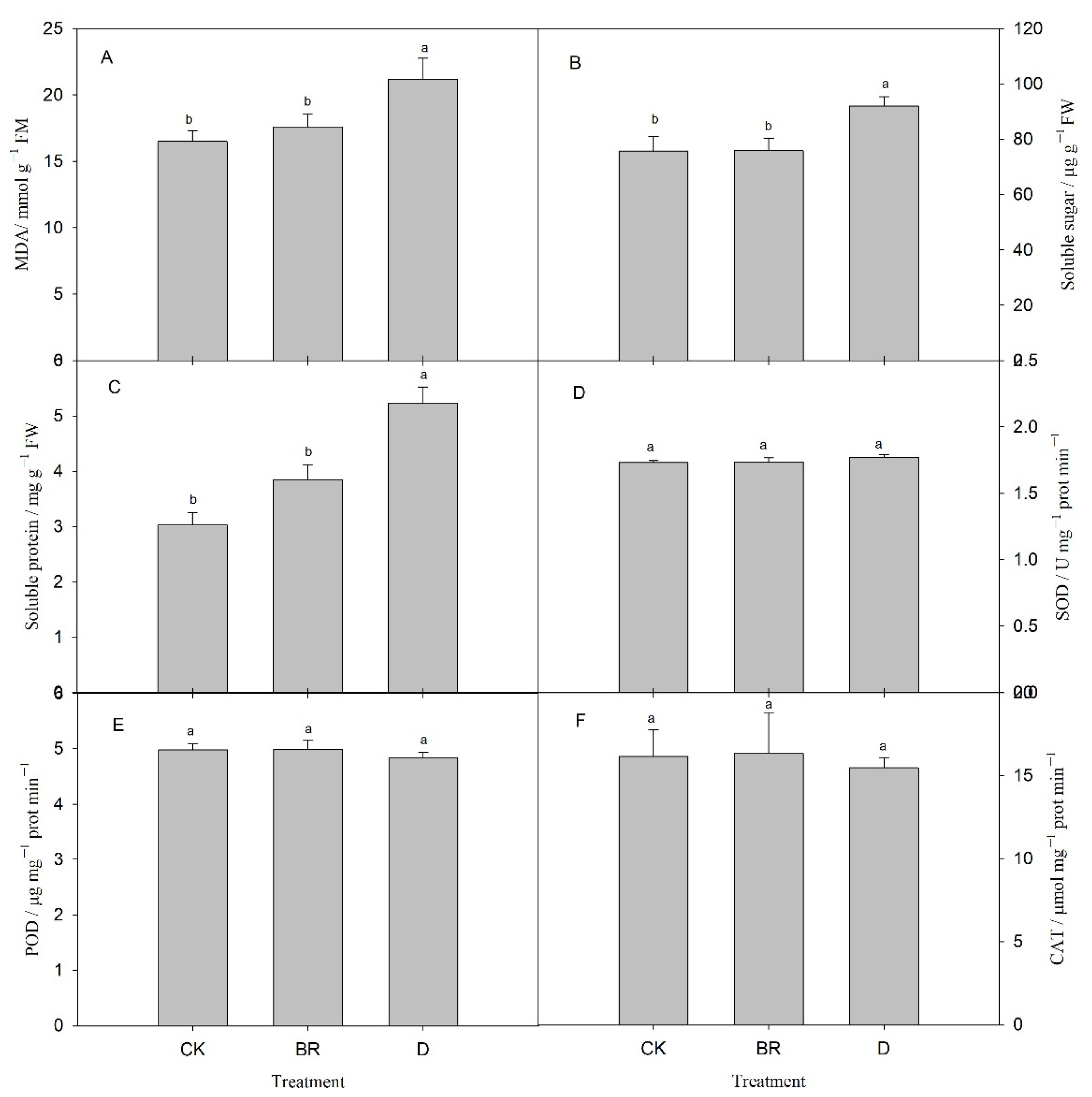
3.2. Effects of BR and Drought on Physiological and Biochemical Substances of R. delavayi
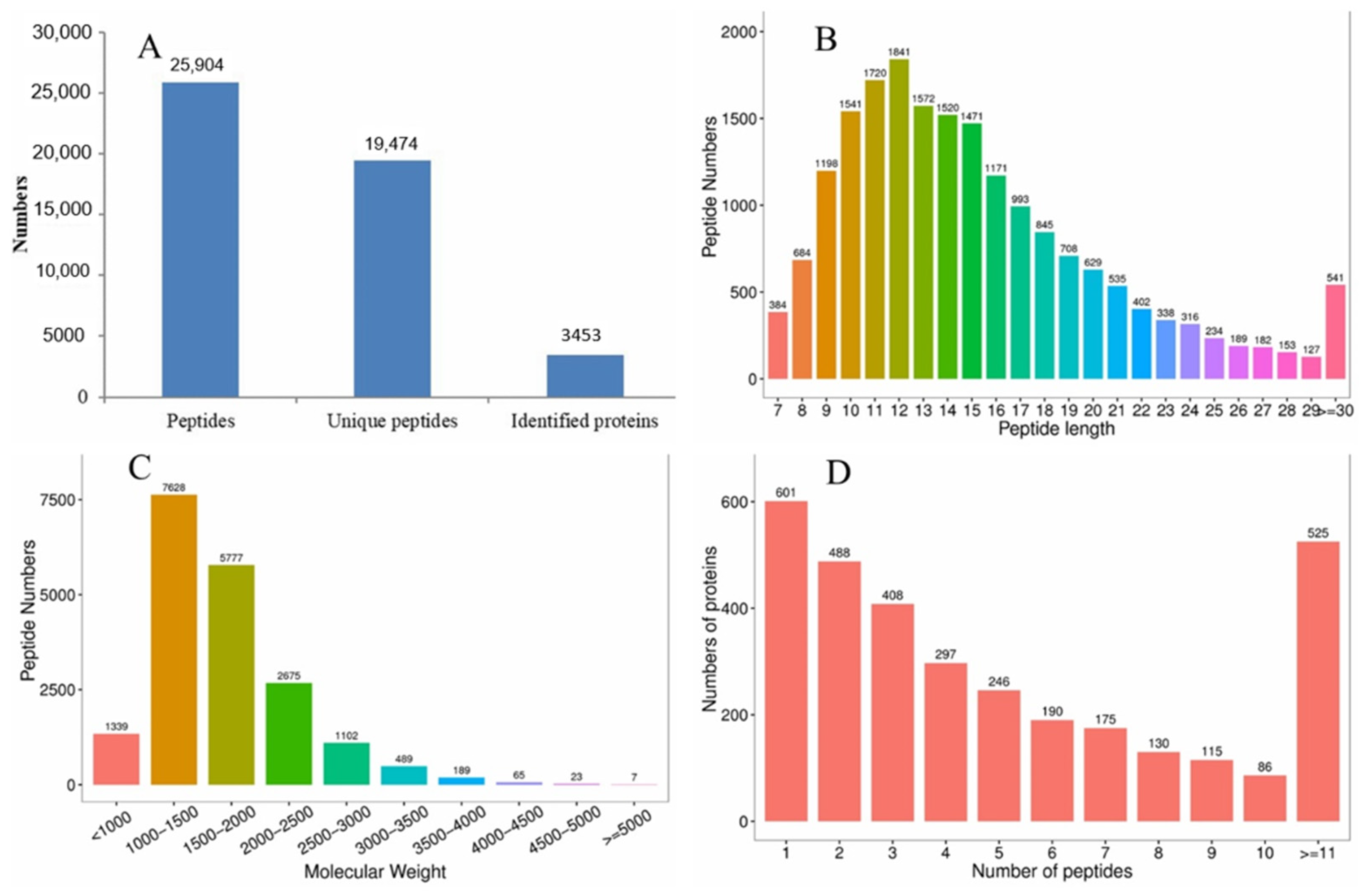
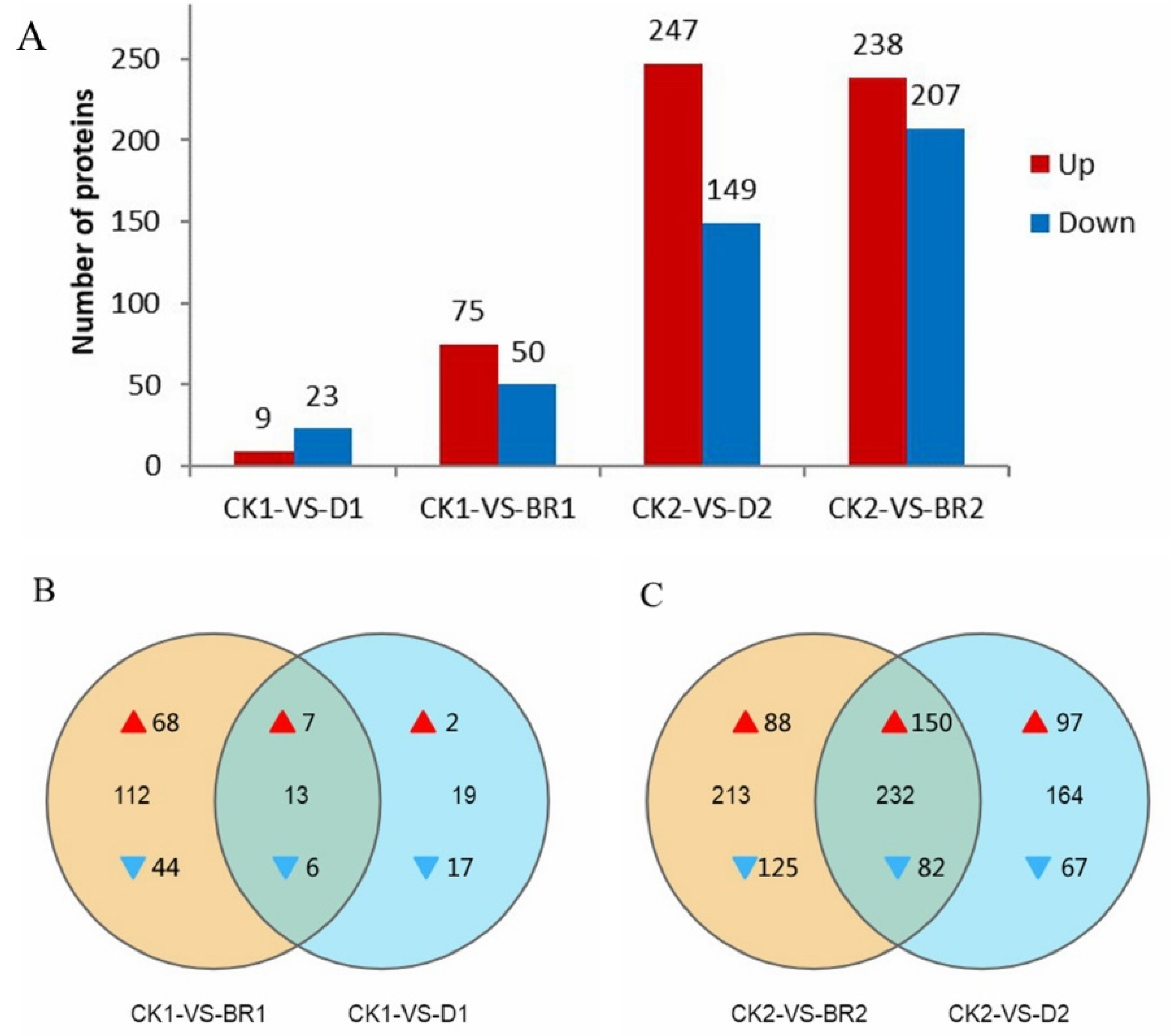
3.3. Quantitative Proteomic Analysis
3.4. Identification of Differentially Expressed Proteins (DEPs)
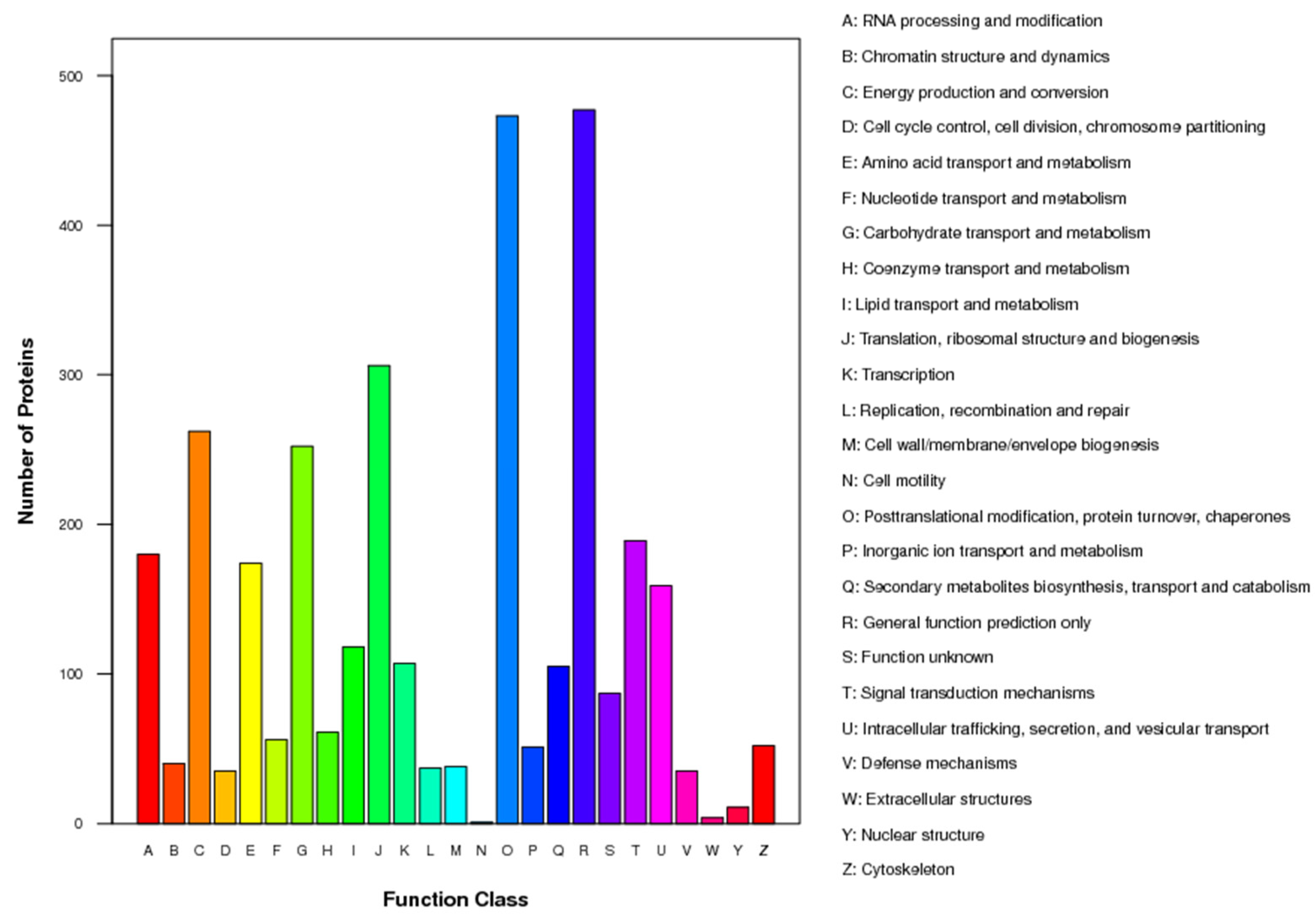
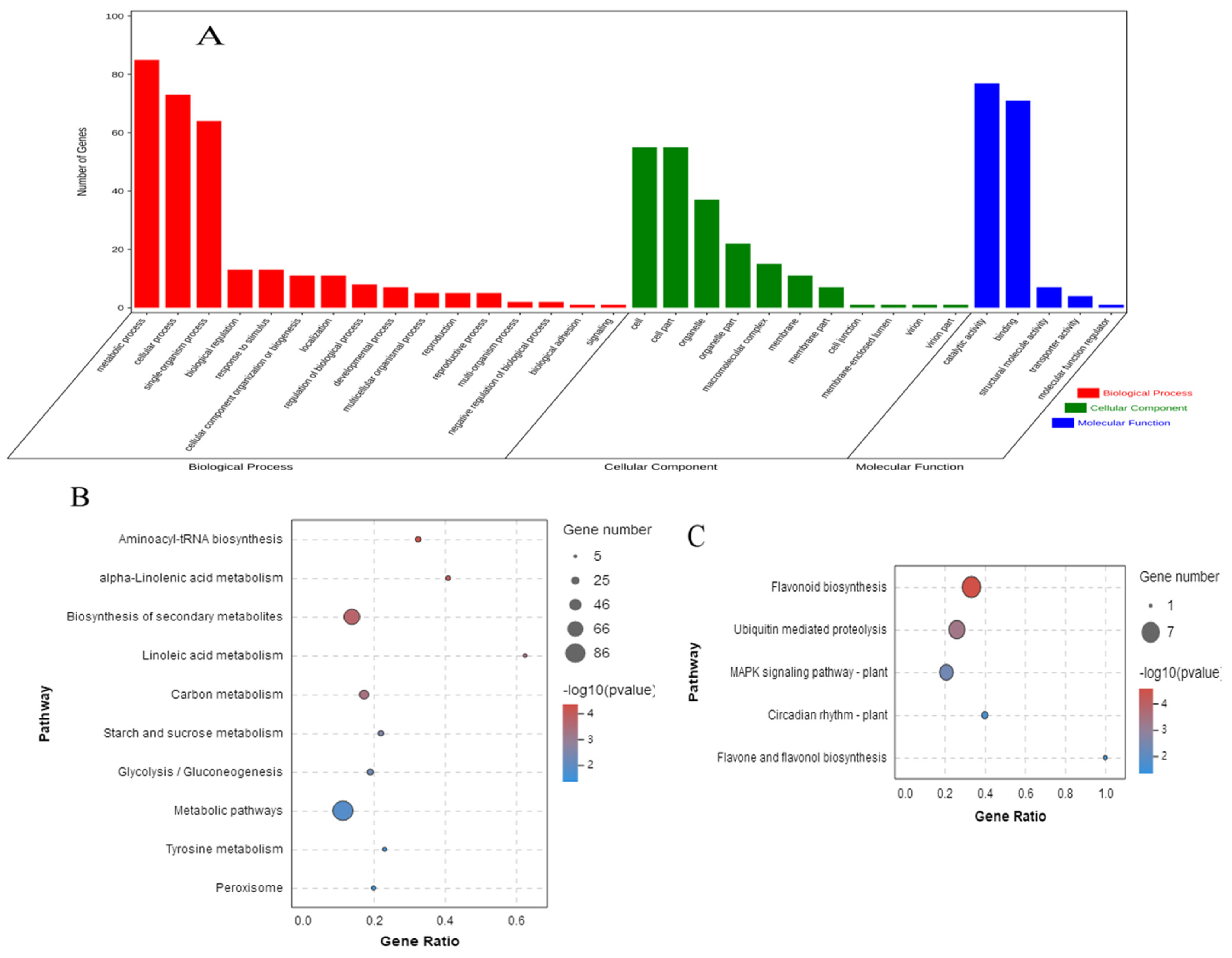
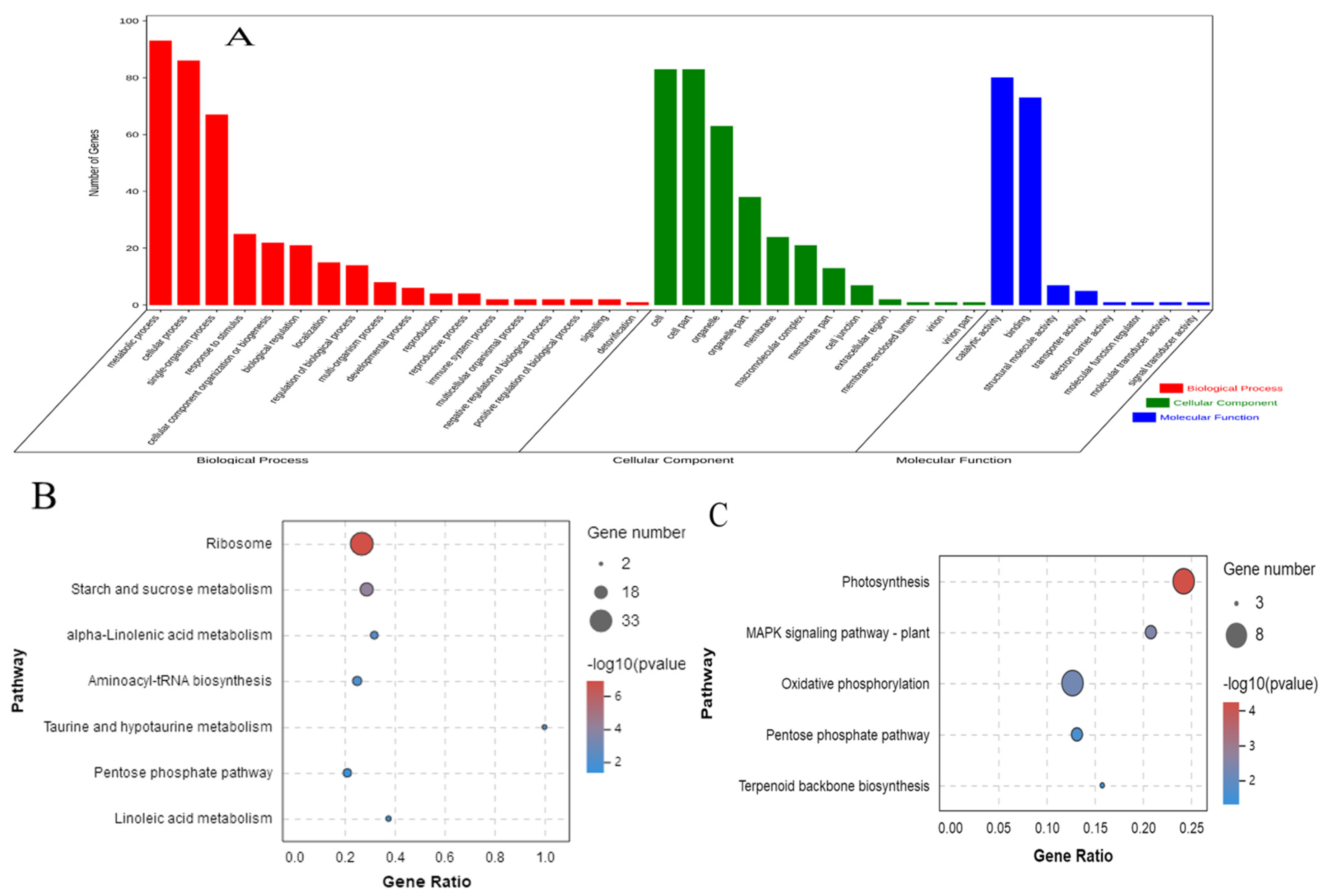
3.5. Functional Classification of DEPs
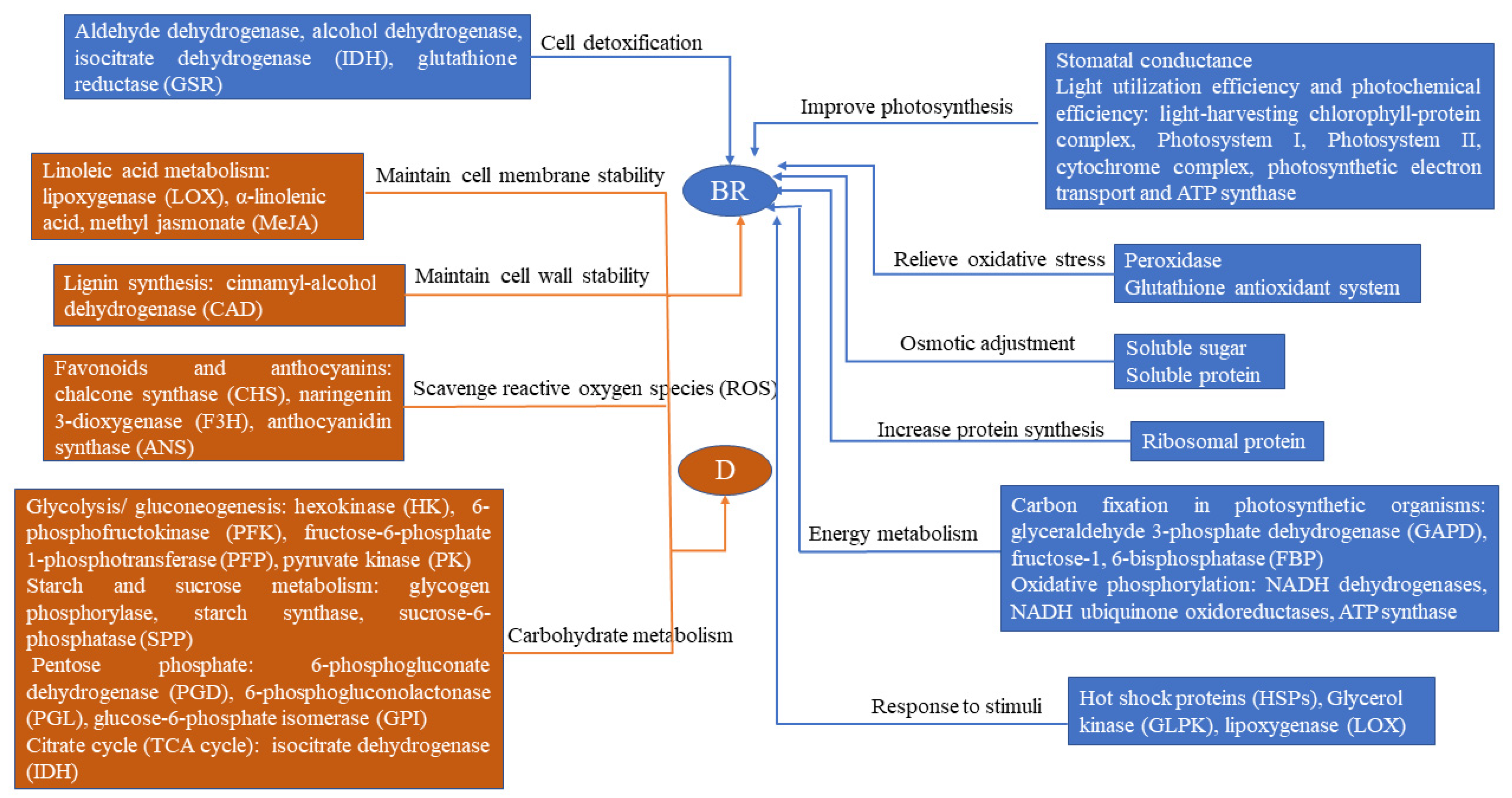
4. Discussion
4.1. Response of Photosynthetic and Physiological Performances to Drought
4.2. Significant Changes of Ribosomal Proteins in the BR Treatment
4.3. Proteins in Relation to Energy Metabolism
4.4. Carbohydrate Metabolism Is Important for R. delavayi to Cope with Drought Stress
4.5. Proteins Involved in Cell Detoxification
4.6. Proteins Related to Lipid Metabolism
4.7. Lignin Synthesis in the Response of R. delavayi to Drought Stress
4.8. Roles of Flavonoid Biosynthesis Proteins in Drought
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hosseini, M.S.; Samsampour, D.; Zahedi, S.M.; Zamanian, K.; Rahman, M.M.; Mostofa, M.G.; Tran, L.S.P. Melatonin alleviates drought impact on growth and essential oil yield of lemon verbena by enhancing antioxidant responses, mineral balance, and abscisic acid content. Physiol. Plant 2021, 172, 1363–1375. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, S.M.; Hosseini, M.S.; Daneshvar Hakimi Meybodi, N.; Peijnenburg, W. Mitigation of the effect of drought on growth and yield of pomegranates by foliar spraying of different sizes of selenium nanoparticles. J. Sci. Food Agric. 2021, 101, 5202–5213. [Google Scholar] [CrossRef] [PubMed]
- Farouk, S.; Omar, M.M. Sweet basil growth, physiological and ultrastructural modification, and oxidative defense system under water deficit and silicon forms treatment. J. Plant Growth Regul. 2020, 39, 1307–1331. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, M.; Morishita, H.; Urano, K.; Shiozaki, N.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Yoshiba, Y. Effects of free proline accumulation in petunias under drought stress. J. Exp. Bot. 2005, 56, 1975–1981. [Google Scholar] [CrossRef] [Green Version]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaleghi, A.; Naderi, R.; Brunetti, C.; Maserti, B.E.; Salami, S.A.; Babalar, M. Morphological, physiochemical and antioxidant responses of Maclura pomifera to drought stress. Sci. Rep. 2019, 9, 19250. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Basra, S.; Islam-ud-Din. Improving water relations and gas exchange with brassinosteroids in rice under drought stress. J. Agron. Crop Sci. 2010, 195, 262–269. [Google Scholar] [CrossRef]
- Soares, C.; de Sousa, A.; Pinto, A.; Azenha, M.; Teixeira, J.; Azevedo, R.A.; Fidalgo, F. Effect of 24-epibrassinolide on ROS content, antioxidant system, lipid peroxidation and Ni uptake in Solanum nigrum L. under Ni stress. Environ. Exp. Bot. 2016, 122, 115–125. [Google Scholar] [CrossRef]
- Gruszka, D. The brassinosteroid signaling pathway—New key players and interconnections with other signaling networks crucial for plant development and stress tolerance. Int. J. Mol. Sci. 2013, 14, 8740–8774. [Google Scholar] [CrossRef] [Green Version]
- Saini, S.; Sharma, I.; Pati, P. Versatile roles of brassinosteroid in plants in the context of its homoeostasis, signaling and crosstalks. Front. Plant Sci. 2015, 6, 950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakirova, F.; Allagulova, C.; Maslennikova, D.; Fedorova, K.; Yuldashev, R.; Lubyanova, A.; Bezrukova, M.; Avalbaev, A. Involvement of dehydrins in 24-epibrassinolide-induced protection of wheat plants against drought stress. Plant Physiol. Biochem. 2016, 108, 539–548. [Google Scholar] [CrossRef]
- Li, X.J.; Guo, X.; Zhou, Y.H.; Shi, K.; Zhou, J.; Yu, J.Q.; Xia, X.J. Overexpression of a brassinosteroid biosynthetic gene Dwarf enhances photosynthetic capacity through activation of Calvin cycle enzymes in tomato. BMC Plant Biol. 2016, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ahammed, G.J.; Gao, C.J.; Ogweno, J.O.; Zhou, Y.H.; Xia, X.J.; Mao, W.H.; Shi, K.; Yu, J.Q. Brassinosteroids induce plant tolerance against phenanthrene by enhancing degradation and detoxification in Solanum lycopersicum L. Ecotoxicol. Environ. Saf. 2012, 80, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Fang, R.; He, M.; Hu, L.; Yang, H.; Chamberlain, D.F. Rhododendron Linnaeus, Sp. Pl. 1: 392. 1753. Flora China 2005, 14, 260–455. [Google Scholar]
- Cai, Y.; Wang, J.; Li, S.; Zhang, L.; Peng, L.; Xie, W.; Liu, F. Photosynthetic response of an alpine plant, Rhododendron delavayi Franch, to water stress and recovery: The role of mesophyll conductance. Front. Plant Sci. 2015, 6, 1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Xu, P.; Cai, Y.; Ma, L.; Li, S.; Li, S.; Xie, W.; Song, J.; Peng, L.; Yan, H.; et al. The draft genome assembly of Rhododendron delavayi Franch. var. delavayi. Gigascience 2017, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.F.; Peng, L.C.; Li, S.F.; Zhang, L.; Xie, W.J.; Song, J.; Wang, J.H. 24-epibrassionlide improves photosynthetic response of Rhododendron delavayi to drought. Nord. J. Bot. 2020, 38, 1–9. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, S.; Wu, H.; Yang, Y.; Xu, H. Biochemical and proteomics analyses of antioxidant enzymes reveal the potential stress tolerance in Rhododendron chrysanthum Pall. Biol. Direct 2017, 12, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Zhang, S.; Huang, Y.; Cai, H.; Zhu, X. Understory and canopy additions of nitrogen differentially affect carbon and nitrogen metabolism of Psychotria rubra in an evergreen broad-leaved forest. Sci. Total Environ. 2020, 724, 138183. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Galmés, J.; Medrano, H.; Ribas-Carbó, M. Keeping a positive carbon balance under adverse conditions: Responses of photosynthesis and respiration to water stress. Physiol. Plant 2006, 127, 343–352. [Google Scholar] [CrossRef]
- Hu, W.H.; Yan, X.H.; Xiao, Y.A.; Zeng, J.J.; Qi, H.J.; Ogweno, J.O. 24-Epibrassinosteroid alleviate drought-induced inhibition of photosynthesis in Capsicum annuum. Sci. Hortic. 2013, 150, 232–237. [Google Scholar] [CrossRef]
- Kolomeichuk, L.; Efimova, M.; Zlobin, I.; Kreslavskii, V.; Murgan, O.; Kovtun, I.; Khripach, V.; Allakhverdiev, S. 24-Epibrassinolide alleviates the toxic effects of NaCl on photosynthetic processes in potato plants. Photosynth. Res. 2020, 146, 151–163. [Google Scholar] [CrossRef]
- Anjum, S.A.; Wang, L.C.; Farooq, M.; Hussain, M.; Xue, L.L.; Zou, C.M. Brassinolide application improves the drought tolerance in maize through modulation of enzymatic antioxidants and leaf gas exchange. J. Agron. Crop Sci. 2011, 197, 177–185. [Google Scholar] [CrossRef]
- Li, H.; Yao, W.; Fu, Y.; Li, S.; Guo, Q. De novo assembly and discovery of genes that are involved in drought tolerance in Tibetan Sophora moorcroftiana. PLoS ONE 2015, 10, e111054. [Google Scholar] [CrossRef] [PubMed]
- Das, T.; Meena, M.; Mandavia, M.K.; Sapre, S.S. Influence of NaCl salt stress on physiological, biochemical changes and isoenzyme pattern in wheat (Triticum aestivum L.) genotypes. Res. Environ. Life Sci. 2015, 8, 825–828. [Google Scholar]
- Gruszka, D. Crosstalk of the brassinosteroid signalosome with phytohormonal and stress signaling components maintains a balance between the processes of growth and stress tolerance. Int. J. Mol. Sci. 2018, 19, 2675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachiri, H.; Djebbar, R.; Mekliche, A. The effect of drought, heat and combined stress on antioxidant enzymes in bread wheat genotypes (Triticum aestivum L.). Analele Univ. Oradea Fasc. Biol. 2020, 27, 56–63. [Google Scholar]
- Moin, M.; Bakshi, A.; Sheshu madhav, M.; Kirti, P. Expression profiling of ribosomal protein gene family in dehydration stress responses and characterization of transgenic rice plants overexpressing RPL23A for water-use efficiency and tolerance to drought and salt stresses. Front. Chem. 2017, 5, 97. [Google Scholar] [CrossRef] [Green Version]
- Fromont-Racine, M.; Senger, B.; Saveanu, C.; Fasiolo, F. Ribosome assembly in eukaryotes. Gene 2003, 313, 17–42. [Google Scholar] [CrossRef]
- Ferreyra, M.L.F.; Pezza, A.; Biarc, J.; Burlingame, A. Plant L10 ribosomal proteins have different roles during development and translation under ultraviolet-B stress. Plant Physiol. 2010, 153, 1878–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehari, T.G.; Xu, Y.; Magwanga, R.O.; Umer, M.J.; Kirungu, J.N.; Cai, X.; Hou, Y.; Wang, Y.; Yu, S.; Wang, K.; et al. Genome wide identification and characterization of light-harvesting Chloro a/b binding (LHC) genes reveals their potential role in enhancing drought tolerance in Gossypium hirsutum. J. Cott. Res. 2021, 4, 15. [Google Scholar] [CrossRef]
- Zenda, T.; Liu, S.; Wang, X.; Liu, G.; Jin, H.; Dong, A.; Yang, Y.; Duan, H. Key maize drought-responsive genes and pathways revealed by comparative transcriptome and physiological analyses of contrasting inbred lines. Int. J. Mol. Sci. 2019, 20, 1268. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.P. How mitochondria produce reactive oxygen species? Biochem. J. 2009, 417, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Siedow, J.N.; Umbach, A.L. The mitochondrial cyanide-resistant oxidase: Structural conservation amid regulatory diversity. BBA Bioenerg. 2000, 1459, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.; Peng, Y.; Zhao, X.; Wu, B.; Chen, F.; Ren, B.; Zhuang, Z.; Gao, Q.; Ding, Y. Comparative proteomics analysis of the seedling root response of drought-sensitive and drought-tolerant maize varieties to drought stress. Int. J. Mol. Sci. 2019, 20, 2793. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, S.M.; Andrade, M.O.; Gomes, A.P.S.; DaMatta, F.M.; Baracat-Pereira, M.C.; Fontes, E.P.B. Arabidopsis and tobacco plants ectopically expressing the soybean antiquitin-like ALDH7 gene display enhanced tolerance to drought, salinity, and oxidative stress. J. Exp. Bot. 2006, 57, 1909–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gullner, G.; Komives, T.; Király, L.; Schröder, P. Glutathione S-transferase enzymes in plant-pathogen interactions. Front. Plant Sci. 2018, 871, 1836. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Zhang, X.; Lu, C.; Zeng, X.; Li, Y.; Fu, D.; Wu, G. Non-specific lipid transfer proteins in plants: Presenting new advances and an integrated functional analysis. J. Exp. Bot. 2015, 66, 5663–5681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafari, M.; Ebadi, A.; Sedghi, M.; Jahanbakhsh, S.; Miransari, M. Alleviating effect of 24- epibrassinolide on seed oil content and fatty acid composition under drought stress in safflower. J. Food Compos. Anal. 2020, 92, 103544. [Google Scholar] [CrossRef]
- Iba, K. Acclimative response to temperature stress in higher plants: Approaches of gene engineering for temperature tolerance. Annu. Rev. Plant Biol. 2002, 53, 225–245. [Google Scholar] [CrossRef] [Green Version]
- Wildhagen, H.; Paul, S.; Allwright, M.; Smith, H.K.; Malinowska, M.; Schnabel, S.K.; Paulo, M.J.; Cattonaro, F.; Vendramin, V.; Scalabrin, S.; et al. Genes and gene clusters related to genotype and drought-induced variation in saccharification potential, lignin content and wood anatomical traits in Populus nigra. Tree Physiol. 2018, 38, 320–339. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Li, Y.; Guo, T. Expression of flavonoid biosynthesis genes and accumulation of flavonoid in wheat leaves in response to drought stress. Plant Physiol. Biochem. 2014, 80, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Liang, M.-X.; DeWald, D.; Weimer, B.; Peel, M.D.; Bugbee, B.; Michaelson, J.; Davis, E.; Wu, Y. Identification of dehydration responsive genes from two non-nodulated alfalfa cultivars using Medicago truncatula microarrays. Acta Physiol. Plant 2008, 30, 183–199. [Google Scholar] [CrossRef]
- Xu, Y.; Gao, S.; Yang, Y.; Huang, M.; Cheng, L.; Wei, Q.; Fei, Z.; Gao, J.; Hong, B. Transcriptome sequencing and whole genome expression profiling of chrysanthemum under dehydration stress. BMC Genom. 2013, 14, 662–676. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Han, Y.; Torres-Jerez, I.; Wang, M.; Tang, Y.; Monteros, M.; Udvardi, M. System responses to long-term drought and re-watering of two contrasting alfalfa varieties. Plant J. 2011, 68, 871–889. [Google Scholar] [CrossRef]
- Cai, Y.F.; Wang, J.H.; Zhang, L.; Song, J.; Peng, L.C.; Zhang, S.B. Physiological and transcriptomic analysis highlight key metabolic pathways in relation to drought tolerance in Rhododendron delavayi. Physiol. Mol. Biol. Plants 2019, 25, 991–1008. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Abrahan, C.; Colquhoun, T.A.; Liu, C.J. A proteolytic regulator controlling chalcone synthase stability and flavonoid biosynthesis in arabidopsis. Plant Cell 2017, 29, 1157–1174. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.; Fu, L.; Tie, W.; Yan, Y.; Wu, C.; Hu, W.; Zhang, J. Extensive post-transcriptional regulation revealed by transcriptomic and proteomic integrative analysis in cassava under drought. J. Agric. Food Chem. 2019, 67, 3521–3534. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | DEPs with Pathway Annotation | All Proteins with Pathway Annotation | p-Value | Q-Value | Pathway ID |
|---|---|---|---|---|---|
| Specific to BR treatment | |||||
| Ribosome | 28 | 123 | 0 | 0 | ko03010 |
| Ether lipid metabolism | 2 | 2 | 0.006 | 0.172 | ko00565 |
| Photosynthesis | 7 | 33 | 0.009 | 0.187 | ko00195 |
| Oxidative phosphorylation | 9 | 63 | 0.041 | 0.627 | ko00190 |
| Specific to D treatment | |||||
| Flavonoid biosynthesis | 6 | 21 | 0.001 | 0.088 | ko00941 |
| Ubiquitin mediated proteolysis | 5 | 23 | 0.012 | 0.268 | ko04120 |
| Biosynthesis of secondary metabolites | 40 | 468 | 0.012 | 0.268 | ko01110 |
| Alpha-linolenic acid metabolism | 4 | 22 | 0.045 | 0.741 | ko00592 |
| Shared between BR and D treatments | |||||
| Starch and sucrose metabolism | 12 | 59 | 0.003 | 0.110 | ko00500 |
| Biosynthesis of secondary metabolites | 53 | 468 | 0.004 | 0.110 | ko01110 |
| Aminoacyl-tRNA biosynthesis | 9 | 40 | 0.004 | 0.110 | ko00970 |
| Alpha-linolenic acid metabolism | 6 | 22 | 0.007 | 0.140 | ko00592 |
| Pentose phosphate pathway | 8 | 38 | 0.011 | 0.165 | ko00030 |
| Tyrosine metabolism | 6 | 26 | 0.017 | 0.218 | ko00350 |
| Linoleic acid metabolism | 3 | 8 | 0.023 | 0.255 | ko00591 |
| Carbon metabolism | 22 | 179 | 0.033 | 0.321 | ko01200 |
| Glycolysis/Gluconeogenesis | 12 | 84 | 0.041 | 0.334 | ko00010 |
| MAPK signaling pathway-plant | 5 | 24 | 0.043 | 0.334 | ko04016 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, Y.-F.; Zhang, L.; Peng, L.-C.; Li, S.-F.; Song, J.; Xie, W.-J.; Wang, J.-H. Key Proteins and Metabolic Pathways Involved in 24-Epibrasionlide Improving Drought Tolerance of Rhododendron delavayi Franch. Horticulturae 2021, 7, 501. https://doi.org/10.3390/horticulturae7110501
Cai Y-F, Zhang L, Peng L-C, Li S-F, Song J, Xie W-J, Wang J-H. Key Proteins and Metabolic Pathways Involved in 24-Epibrasionlide Improving Drought Tolerance of Rhododendron delavayi Franch. Horticulturae. 2021; 7(11):501. https://doi.org/10.3390/horticulturae7110501
Chicago/Turabian StyleCai, Yan-Fei, Lu Zhang, Lv-Chun Peng, Shi-Feng Li, Jie Song, Wei-Jia Xie, and Ji-Hua Wang. 2021. "Key Proteins and Metabolic Pathways Involved in 24-Epibrasionlide Improving Drought Tolerance of Rhododendron delavayi Franch" Horticulturae 7, no. 11: 501. https://doi.org/10.3390/horticulturae7110501
APA StyleCai, Y. -F., Zhang, L., Peng, L. -C., Li, S. -F., Song, J., Xie, W. -J., & Wang, J. -H. (2021). Key Proteins and Metabolic Pathways Involved in 24-Epibrasionlide Improving Drought Tolerance of Rhododendron delavayi Franch. Horticulturae, 7(11), 501. https://doi.org/10.3390/horticulturae7110501