
Satureja montana Essential Oil, Zein Nanoparticles and Their Combination as a Biocontrol Strategy to Reduce Bacterial Spot Disease on Tomato Plants

 ,
,  ,
,  , ,
, ,  ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Essential Oil Obtention, Synthesis of Zein Nanoparticles, and EO-Zein NPs
2.2. Plant Growth Conditions and Treatments
2.3. Xeu Quantification
2.4. Quantification of Reactive Oxygen Species
2.4.1. H2O2 Concentration in Leaves
2.4.2. Superoxide Radical Concentration in Leaves
2.5. Non-Enzymatic Antioxidant Capacity (NEAC)
2.5.1. Total Phenol Content Quantification
2.5.2. Antiradical Activity
2.6. Antioxidant Enzyme Activity
2.7. Gene Expression Analysis
2.8. Statistical Analysis
3. Results
3.1. NP Formation Efficiency, EO Encapsulation Efficiency and NP Stability during Storage
3.2. Xeu Quantification and Disease Symptom Monitoring
3.3. ROS Quantification
3.4. Non-Enzymatic Antioxidant Capacity
3.5. Antioxidant Enzymatic Activity
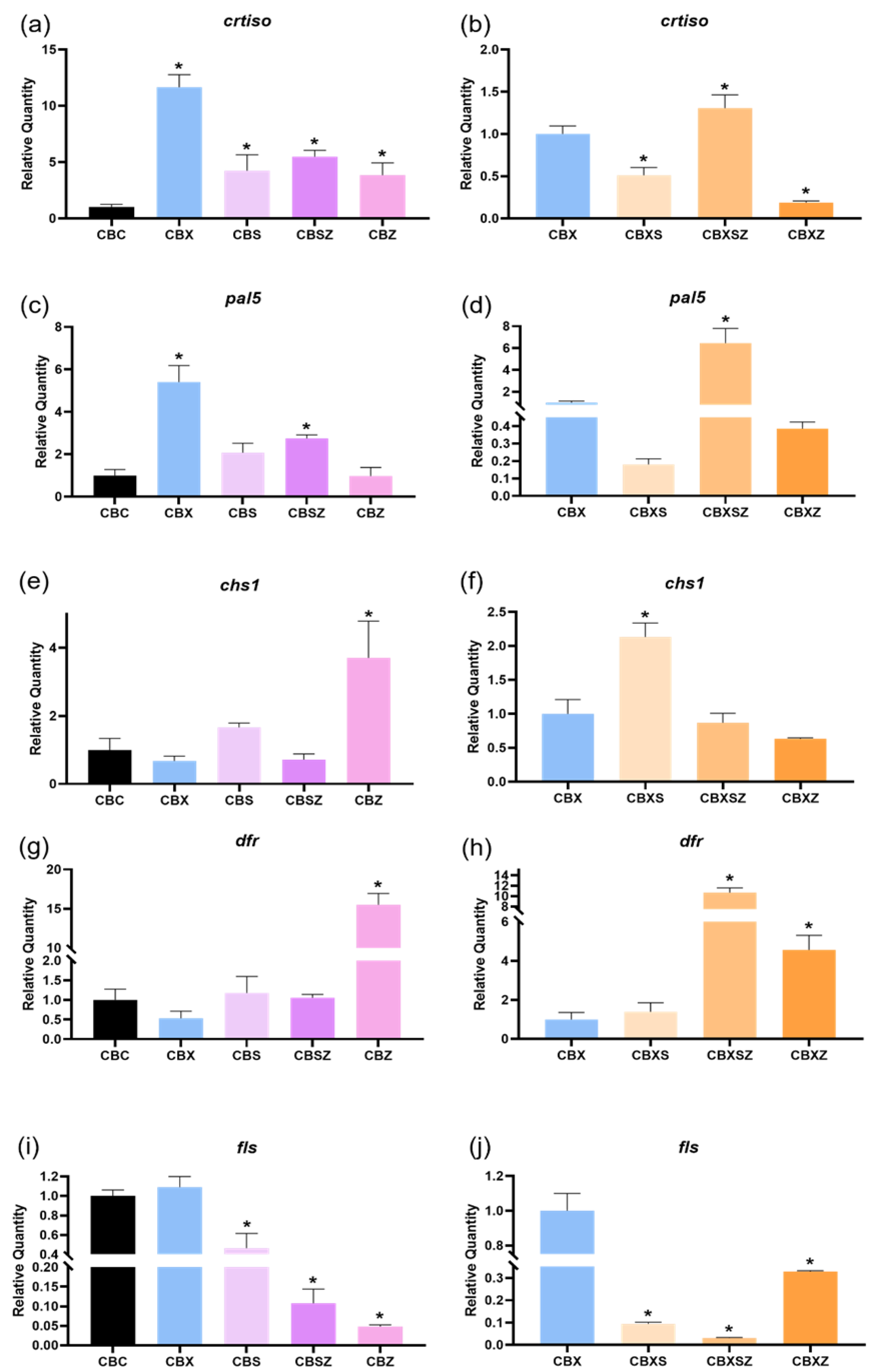
3.6. Gene Expression
3.6.1. Phenylpropanoid Pathway
3.6.2. Enzymatic Pathway
3.6.3. Hormone Pathway
3.7. Principle Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Guan, Z.; Biswas, T.; Wu, F. The US Tomato Industry: An Overview of Production and Trade: FE1027, 9/2017, 1st ed.; EDIS: Gainesville, FL, USA, 2018. [Google Scholar]
- Horvath, D.M.; Stall, R.E.; Jones, J.B.; Pauly, M.H.; Vallad, G.E.; Dahlbeck, D.; Staskawicz, B.J.; Scott, J.W. Transgenic resistance confers effective field level control of bacterial spot disease in tomato. PLoS ONE 2012, 7, e42036. [Google Scholar] [CrossRef]
- Potnis, N.; Timilsina, S.; Strayer, A.; Shantharaj, D.; Barak, J.D.; Paret, M.L.; Vallad, G.E.; Jones, J.B. Bacterial spot of tomato and pepper: Diverse Xanthomonas species with a wide variety of virulence factors posing a worldwide challenge. Mol. Plant Pathol. 2015, 16, 907–920. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.B.; Bouzar, H.; Stall, R.E.; Amlira, E.C.; Roberts, P.D.; Bowen, B.W.; Sudberry, J.; Strickler, P.M.; Chun, J. Systematic analysis of Xanthomonads (Xanthomonas spp.) associated with pepper and tomato lesions. Intl. J. Syst. Evol. Microbiol. 2000, 50, 1211–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egel, D.S.; Jones, J.B.; Minsavage, G.V.; Creswell, T.; Ruhl, G.; Maynard, E.; Marchino, C. Distribution and characterization of Xanthomonas strains causing bacterial spot of tomato in Indiana. Plant Health Prog. 2018, 19, 319–321. [Google Scholar] [CrossRef]
- Itako, A.T.; Tolentino, J.B.; Silva, T.A.F.D.; Soman, J.M.; Maringoni, A.C. Chemical products induce resistance to Xanthomonas perforans in tomato. Braz. J. Microbiol. 2015, 46, 701–706. [Google Scholar] [CrossRef]
- Liao, Y.Y.; Strayer-Scherer, A.L.; White, J.; Mukherjee, A.; De La Torre-Roche, R.; Ritchie, L.; Colee, J.; Vallad, G.E.; Freeman, J.H.; Jones, J.B.; et al. Nano-Magnesium oxide: A novel bactericide against copper-tolerant Xanthomonas perforans causing tomato bacterial spot. Phytopathology 2019, 109, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Zhang, S.; Huang, Y.; Jones, J.B. Evaluation of a small molecule compound 3-indolylacetonitrile for control of bacterial spot on tomato. Crop Prot. 2019, 120, 7–12. [Google Scholar] [CrossRef]
- Vallad, G.E.; Pernezny, K.L.; Balogh, B.; Wen, A.; Figueiredo, J.F.L.; Jones, J.B.; Momol, T.; Muchovej, R.M.; Havranek, N.; Abdallah, N.; et al. Comparison of kasugamycin to traditional bactericides for the management of bacterial spot on tomato. HortScience 2010, 45, 1834–1840. [Google Scholar] [CrossRef]
- Strayer-Scherer, A.; Liao, Y.Y.; Abrahamian, P.; Timilsina, S.; Paret, M.; Momol, T.; Jones, J.; Vallad, G.E. Integrated Management of Bacterial Spot on Tomato in Florida, 1st ed.; EDIS: Gainesville, FL, USA, 2019; p. 8. [Google Scholar] [CrossRef]
- Jamiołkowska, A. Natural compounds as elicitors of plant resistance against diseases and new biocontrol strategies. Agronomy 2020, 10, 173. [Google Scholar] [CrossRef] [Green Version]
- Arasu, M.V.; Al-Dhabi, N.A.; Choi, K.C.; Bensy, A.D.V.; Rajaselvam, J. Bioactive potential of Albizia lebbeck extract against phytopathogens and protective properties on tomato plant against speck disease in greenhouse. Physiol. Mol. Plant Pathol. 2021, 117, 101750. [Google Scholar] [CrossRef]
- Liberto, M.G.D.; Seimandi, G.M.; Fernández, L.N.; Ruiz, V.E.; Svetaz, L.A.; Derita, M.G. Botanical Control of Citrus Green Mold and Peach Bro wn Rot on Fruits Assays Using a Persicaria acuminata Phytochemically Characterized Extract. Plants 2021, 10, 425. [Google Scholar] [CrossRef] [PubMed]
- Tamm, L.; Thürig, B.; Fliessbach, A.; Goltlieb, A.E.; Karavani, S.; Cohen, Y. Elicitors and soil management to induce resistance against fungal plant diseases. NJAS Wagening. J. Life Sci. 2011, 58, 131–137. [Google Scholar] [CrossRef]
- Burketova, L.; Trda, L.; Ott, P.G.; Valentova, O. Bio-based resistance inducers for sustainable plant protection against pathogens. Biotechnol. Adv. 2015, 33, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, V.K.; Kang, S.R.; Xu, H.; Lee, S.G.; Baek, K.H.; Kang, S.C. Potential roles of essential oils on controlling plant pathogenic bacteria Xanthomonas species: A review. Plant Pathol. J. 2011, 27, 207–224. [Google Scholar] [CrossRef] [Green Version]
- Pepa, T.D.; Elshafie, H.S.; Capasso, R.; De Feo, V.; Camele, I.; Nazzaro, F.; Scognamiglio, M.R.; Caputo, L. Antimicrobial and phytotoxic activity of Origanum heracleoticum and O. majorana essential oils growing in Cilento (Southern Italy). Molecules 2019, 24, 2576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkan, D.; Yemenicioğlu, A. Potential application of natural phenolic antimicrobials and edible film technology against bacterial plant pathogens. Food Hydrocoll. 2016, 55, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Abo-Elyousr, K.A.; Bagy, H.M.K.; Hashem, M.; Alamri, S.A.; Mostafa, Y.S. Biological control of the tomato wilt caused by Clavibacter michiganensis subsp. michiganensis using formulated plant growth-promoting bacteria. Egypt. J. Biol. Pest Control 2019, 29, 54. [Google Scholar] [CrossRef]
- Franz, C.; Novak, J. Sources of essential oils. In Handbook of Essential Oils; Baser, K.H.C., Buchbauer, G., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 41–83. [Google Scholar]
- Mollova, S.; Fidan, H.; Antonova, D.; Bozhilov, D.; Stanev, S.; Kostova, I.; Stoyanova, A. Chemical composition and antimicrobial and antioxidant activity of Helichrysum italicum (Roth) G. Don subspecies essential oils. Turk. J. Agric. For. 2020, 44, 371–378. [Google Scholar] [CrossRef]
- Magi, G.; Marini, E.; Facinelli, B. Antimicrobial activity of essential oils and carvacrol, and synergy of carvacrol and erythromycin, against clinical, erythromycin-resistant Group A Streptococci. Front. Microbiol. 2015, 6, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chouhan, S.; Sharma, K.; Guleria, S. Antimicrobial activity of some essential oils—Present status and future perspectives. Medicines 2017, 4, 58. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Chen, Q.; Liang, Q.; Zhang, M.; Chen, W.; Chen, H.; Yun, Y.; Zhong, Q.; Chen, W. Antimicrobial Activity and Proposed Action Mechanism of Linalool against Pseudomonas fluorescens. Front. Microbiol. 2021, 12, 49. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Cho, M.J.; Kang, S.C. Control of plant pathogenic bacteria of Xanthomonas spp. by the essential oil and extracts of Metasequoia glyptostroboides Miki ex Hu In Vitro and In Vivo. J. Phytopathol. 2010, 158, 479–486. [Google Scholar] [CrossRef]
- Altundag, S.; Aslim, B.E.L.M.A.; Ozturk, S. In Vitro Antimicrobial Activities of Essential Oils from Origanum minutiflorum and Sideritis erytrantha subsp. erytrantha on Phytopathogenic Bacteria. J. Essent. Oil Res. 2011, 23, 4–8. [Google Scholar] [CrossRef]
- Costa, P.; Medronho, B.; Goncalves, S.; Romano, A. Cyclodextrins enhance the antioxidant activity of essential oil. Ind. Crop. Prod. 2015, 70, 341–346. [Google Scholar] [CrossRef]
- Cunha, A.P.; Ribeiro, J.A.; Roque, O.R. Plantas Aromáticas em Portugal. Caracterização e Utilizações, 1st ed.; Fundação Calouste Gulbenkian: Lisbon, Portugal, 2007. [Google Scholar]
- Maccelli, A.; Vitanza, L.; Imbriano, A.; Frascetti, C.; Filippi, A.; Goldoni, P.; Maurizi, L.; Ammendolia, M.G.; Crestoni, M.E.; Fornarini, S.; et al. Satureja montana L. essential oil/chemical profiles/phitochemical screening, antimicrobial activity and O/W nanoemulsion formulations. Pharmaceutics 2020, 12, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitanza, L.; Maccelli, A.; Marazzato, M.; Scazzocchio, F.; Comanducci, A.; Fornarini, A.; Crestoni, M.E.; Filippi, A.; Frascetti, C.; Rinaldi, F.; et al. Satureja montana L. essential oil and its activity alone or in combination with gentamicin. Microb. Pathog. 2018, 126, 323–331. [Google Scholar] [CrossRef]
- García-Díaz, M.; Gil-Serna, J.; Patiño, B.; García-Cela, E.; Magan, N.; Medina, A. Assessment of the effect of Satureja montana and Organum virens essential oils on Aspergillus flavus growth and aflatoxin production at different water activities. Toxins 2020, 12, 142. [Google Scholar] [CrossRef] [Green Version]
- Kotan, R.; Dadasoğlu, F.; Karagoz, K.; Cakir, A.; Ozer, H.; Kordali, S.; Cakmakci, R.; Dikbas, N. Antibacterial activity of the essential oil and extracts of Satureja hortensis against plant pathogenic bacteria and their potential use as seed disinfectants. Sci. Hortic. 2013, 153, 34–41. [Google Scholar] [CrossRef]
- Maurya, A.K.; Sharma, A.; Kumar, K.; Chander, R.; Kumar, A.; Kumar, D.; Padwad, Y.S.; Chand, G.; Agnihotri, V.K. Comparative studies of essential oils composition and cytotoxic activity of Valeriana jatamansi Jones. J. Essent. Oil Res. 2021, 1–8. [Google Scholar] [CrossRef]
- Wang, H.; Zhu, W.; Huang, Y.; Li, Z.; Jiang, Y.; Xie, Q. Facile encapsulation of hydroxycamptothecin nanocrystals into zein-based nanocomplexes for active targeting in drug delivery and cell imaging. Acta Biomater. 2017, 61, 88–100. [Google Scholar] [CrossRef]
- Patel, A.; Hu, Y.; Tiwari, J.K.; Velikov, K.P. Synthesis and characterisation of zein–curcumin colloidal particles. Soft Matter 2010, 6, 6192–6199. [Google Scholar] [CrossRef]
- Paliwal, R.; Palakurthi, S. Zein in controlled drug delivery and tissue engineering. J. Control. Release 2014, 189, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Bidyarani, N.; Srivastav, A.K.; Gupta, S.K.; Kumar, U. Synthesis and physicochemical characterization of rhamnolipid-stabilized carvacrol-loaded zein nanoparticles for antimicrobial application supported by molecular docking. J. Nanoparticle Res. 2020, 22, 307. [Google Scholar] [CrossRef]
- Yilmaz, M.T.; Akman, P.K.; Bozkurt, F.; Karasu, S. An effective polydopamine coating to improve stability and bioactivity of carvacrol-loaded zein nanoparticles. Int. J. Food Sci. Technol. 2021, 56, 6011–6024. [Google Scholar] [CrossRef]
- Merino, N.; Berdejo, D.; Bento, R.; Salman, H.; Lanz, M.; Maggi, F.; Sánchez-Gómez, S.; García-Gonzaloa, D.; Pagán, R. Antimicrobial efficacy of Thymbra capitata (L.) Cav. essential oil loaded in self-assembled zein nanoparticles in combination with heat. Ind. Crop. Prod. 2019, 133, 98–104. [Google Scholar] [CrossRef]
- Monteiro, R.A.; Camara, M.C.; de Oliveira, J.L.; Campos, E.V.R.; Carvalho, L.B.; de Freitas Proença, P.L.; Guilger-Casagrande, M.; Lima, R.; do Nascimento, J.; Gonçalves, K.C.; et al. Zein based-nanoparticles loaded botanical pesticides in pest control: An enzyme stimuli-responsive approach aiming sustainable agriculture. J. Hazard. Mater. 2021, 417, 126004. [Google Scholar] [CrossRef]
- Matsuda, Y.; Suzuki, T.; Sato, E.; Sato, M.; Koizumi, S.; Unno, K.; Kato, T.; Nakai, K. Novel preparation of zein microspheres conjugated with PS-K available for cancer immunotherapy. Chem. Pharm. Bull. 1989, 37, 757–759. [Google Scholar] [CrossRef] [Green Version]
- Moretti, C.; Amatulli, M.T.; Buonaurio, R. PCR-based assay for the detection of Xanthomonas euvesicatoria causing pepper and tomato bacterial spot. Lett. Appl. Microbiol. 2009, 49, 466–471. [Google Scholar] [CrossRef]
- Zhao, S.; Fernald, R.D. Comprehensive algorithm for quantitative real-time polymerase chain reaction. J. Comput. Biol. 2005, 12, 1047–1064. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Gajewska, E.; Sklodowska, M. Effect of nickel on ROS content and antioxidative enzyme activities in wheat leaves. BioMetals 2007, 20, 27–36. [Google Scholar] [CrossRef]
- Costa-Santos, M.; Mariz-Ponte, N.; Dias, M.C.; Moura, L.; Marques, G.; Santos, C. Effect of Bacillus spp. and Brevibacillus sp. on the Photosynthesis and Redox Status of Solanum lycopersicum. Horticulturae 2021, 7, 24. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef]
- Harkat-Madouri, L.; Asma, B.; Madani, K.; Said, Z.B.O.S.; Rigou, P.; Grenier, D.; Allalou, H.; Remini, H.; Adjaoud, A.; Boulekbache-Makhlouf, L. Chemical composition, antibacterial and antioxidant activities of essential oil of Eucalyptus globulus from Algeria. Ind. Crop. Prod. 2015, 78, 148–153. [Google Scholar] [CrossRef]
- Mariz-Ponte, N.; Mendes, R.J.; Sario, S.; Melo, P.; Santos, C. Moderate UV-A supplementation benefits tomato seed and seedling invigoration: A contribution to the use of UV in seed technology. Sci. Hortic. 2018, 235, 357–366. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Løvdal, T.; Lillo, C. Reference gene selection for quantitative real-time PCR normalization in tomato subjected to nitrogen, cold, and light stress. Anal. Biochem. 2009, 387, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Mariz-Ponte, N.; Mendes, R.J.; Sario, S.; De Oliveira, J.F.; Melo, P.; Santos, C. Tomato plants use non-enzymatic antioxidant pathways to cope with moderate UV-A/B irradiation: A contribution to the use of UV-A/B in horticulture. J. Plant Physiol. 2018, 221, 32–42. [Google Scholar] [CrossRef]
- Murshed, R.; Lopez-Lauri, F.; Sallanon, H. Effect of water stress on antioxidant systems and oxidative parameters in fruits of tomato (Solanum lycopersicon L, cv. Micro-tom). Physiol. Mol. Biol. Plants 2013, 19, 363–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horváth, E.; Csiszár, J.; Gallé, Á.; Poór, P.; Szepesi, Á.; Tari, I. Hardening with salicylic acid induces concentration-dependent changes in abscisic acid biosynthesis of tomato under salt stress. J. Plant Physiol. 2015, 183, 54–63. [Google Scholar] [CrossRef]
- Niwa, T.; Suzuki, T.; Takebayashi, Y.; Ishiguro, R.; Higashiyama, T.; Sakakibara, H.; Ishiguro, S. Jasmonic acid facilitates flower opening and floral organ development through the upregulated expression of SlMYB21 transcription factor in tomato. Biosci. Biotechnol. Biochem. 2018, 82, 292–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, D.F.; Lopes, E.A.; Moraes, A.R.F.; Soares, M.S.; Visôtto, L.E.; Oliveira, C.R.; Valente, V.M.M. Formulation of botanicals for the control of plant-pathogens: A review. Crop Prot. 2018, 110, 135–140. [Google Scholar] [CrossRef]
- Podaralla, S.; Perumal, O. Influence of Formulation Factors in the Precipitation of Zein Nanoparticles. AAPS PharmSciTech 2012, 13, 919–927. [Google Scholar] [CrossRef] [Green Version]
- Parris, N.; Cooke, P.H.; Hicks, K.B. Encapsulation of Essential Oils in Zein Nanospherical Particles. J. Agric. Food Chem. 2005, 53, 4788–4792. [Google Scholar] [CrossRef]
- Poyatos-Racionero, E.; Guarí-Borràs, G.; Ruiz-Rico, M.; Morellá-Aucejo, Á.; Aznar, E.; Barat, J.M.; Martínez-Máñez, R.; Marcos, M.D.; Bernardos, A. Towards the Enhancement of Essential Oil Components’ Antimicrobial Activity Using New Zein Protein-Gated Mesoporous Silica Microdevices. Int. J. Mol. Sci. 2021, 22, 3795. [Google Scholar] [CrossRef]
- Xiao, D.; Davidson, P.M.; Zhong, Q. Spray-dried zein capsules with coencapsulated nisin and thymol as antimicrobial delivery system for enhanced antilisterial properties. J. Agric. Food Chem. 2011, 59, 7393–7404. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Luo, Y.; Wang, Q. Antioxidant and antimicrobial properties of essential oils encapsulated in zein nanoparticles prepared by liquid–liquid dispersion method. LWT 2012, 48, 283–290. [Google Scholar] [CrossRef]
- Li, K.K.; Yin, S.W.; Yang, X.Q.; Tang, C.H.; Wei, Z.H. Fabrication and characterization of novel antimicrobial films derived from thymol-loaded zein–sodium caseinate (SC) nanoparticles. J. Agric. Food Chem. 2012, 60, 11592–11600. [Google Scholar] [CrossRef]
- Gonçalves, J.; Torres, N.; Silva, S.; Gonçalves, F.; Noro, J.; Cavaco-Paulo, A.; Ribeiro, A.; Silva, C. Zein impart hydrophobic and antimicrobial properties to cotton textiles. React. Funct. Polym. 2020, 154, 104664. [Google Scholar] [CrossRef]
- Irache, J.M.; González-Navarro, C.J. Zein nanoparticles as vehicles for oral delivery purposes. Nanomedicine 2017, 12, 1209–1211. [Google Scholar] [CrossRef]
- Ibáñez, M.D.; López-Gresa, M.P.; Lisón, P.; Rodrigo, I.; Bellés, J.M.; González-Mas, M.C.; Blázquez, M.A. Essential Oils as Natural Antimicrobial and Antioxidant Products in the Agrifood Indus. Nereis 2020, 12, 55–69. [Google Scholar] [CrossRef]
- Choudhary, P.; Aggarwal, P.R.; Rana, S.; Nagarathnam, R.; Muthamilarasan, M. Molecular and metabolomic interventions for identifying potential bioactive molecules to mitigate diseases and their impacts on crop plants. Physiol. Mol. Plant Pathol. 2021, 114, 101624. [Google Scholar] [CrossRef]
- Torres, M.A. ROS in biotic interactions. Physiol. Plant. 2010, 138, 414–429. [Google Scholar] [CrossRef]
- Bhattarai, K.; Louws, F.J.; Williamson, J.D.; Panthee, D.R. Differential response of tomato genotypes to Xanthomonas-specific pathogen-associated molecular patterns and correlation with bacterial spot (Xanthomonas perforans) resistance. Hortic. Res. 2016, 3, 16035. [Google Scholar] [CrossRef]
- Chandrashekar, S.; Umesha, S. Induction of antioxidant enzymes associated with bacterial spot pathogenesis in tomato. Int. J. Food Agric. Vet. Sci. 2012, 2, 22–34. [Google Scholar]
- Silveira, P.R.; Nascimento, K.J.T.; Andrade, C.C.L.; Bispo, W.M.S.; Oliveira, J.R.; Rodrigues, F.A. Physiological changes in tomato leaves arising from Xanthomonas gardneri infection. Physiol. Mol. Plant Pathol. 2015, 92, 130–138. [Google Scholar] [CrossRef]
- Zeiss, D.R.; Mhlongo, M.I.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A. Metabolomic profiling of the host response of tomato (Solanum lycopersicum) following infection by Ralstonia solanacearum. Int. J. Mol. Sci. 2019, 20, 3945. [Google Scholar] [CrossRef] [Green Version]
- García-Andrade, J.; González, B.; Gonzalez-Guzman, M.; Rodriguez, P.L.; Vera, P. The role of ABA in plant immunity is mediated through the PYR1 receptor. Int. J. Mol. Sci. 2020, 21, 5852. [Google Scholar] [CrossRef]
- Lievens, L.; Pollier, J.; Goossens, A.; Beyaert, R.; Staal, J. Abscisic acid as pathogen effector and immune regulator. Front. Plant Sci. 2017, 8, 587. [Google Scholar] [CrossRef]
- Sánchez-Vallet, A.; López, G.; Ramos, B.; Delgado-Cerezo, M.; Riviere, M.P.; Llorente, F.; Fernández, P.V.; Miedes, E.; Estevez, J.M.; Grant, M.; et al. Disruption of abscisic acid signaling constitutively activates Arabidopsis resistance to the necrotrophic fungus Plectosphaerella cucumerina. Plant Physiol. 2012, 160, 2109–2124. [Google Scholar] [CrossRef] [Green Version]
- Ramegowda, V.; Senthil-Kumar, M.; Udayakumar, M.; Mysore, K.S. A high-throughput virus-induced gene silencing protocol identifies genes involved in multi-stress tolerance. BMC Plant Biol. 2013, 13, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Li, Y.; Zhang, Y.; Sheng, J.; Zhu, H.; Shen, L. Transcriptome analysis reveals that SlNPR1 mediates tomato fruit resistance against Botrytis cinerea by modulating phenylpropanoid metabolism and balancing ROS homeostasis. Postharvest Biol. Tec. 2021, 172, 111382. [Google Scholar] [CrossRef]
- Mysore, K.S.; D’Ascenzo, M.D.; He, X.; Martin, G.B. Overexpression of the disease resistance gene Pto in tomato induces gene expression changes similar to immune responses in human and fruitfly. Plant Physiol. 2003, 132, 1901–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene. | Primer | Sequence | Annealing Temperature (°C) | Product Size (bp) | Reference |
|---|---|---|---|---|---|
| ubi | Forward | GGACGGACGTACTCTAGCTGAT | 60 | 134 | [51] |
| Reverse | AGCTTTCGACCTCAAGGGTA | ||||
| tub | Forward | AACCTCCATTCAGGAGATGTTT | 60 | 180 | [51] |
| Reverse | TCTGCTGTAGCATCCTGGTATT | ||||
| crtiso | Forward | GTTTGTAATCTTGGGTTTCCAGCA | 60 | 117 | This Study |
| Reverse | TTGCCTTGTGGGTTTCAAGC | ||||
| fls | Forward | ATAGCTCCACAACCAGGTGC | 60 | 108 | [52] |
| Reverse | TCCATTTGGCCTCACCACTC | ||||
| dfr | Forward | TGCCCTTTCACTAATTACCGCT | 60 | 107 | This Study |
| Reverse | CCTTGGGGTGCTCATACAGG | ||||
| pal5 | Forward | TGGAGGAGAATTTGAAGAATGCTG | 60 | 136 | This Study |
| Reverse | TCCCTTTCCACCACTTGTAGC | ||||
| chs1 | Forward | ACCAACAAGGTTGCTTTGCC | 60 | 135 | [52] |
| Reverse | GAGATTCACTGGGTCCACGG | ||||
| apxcyto | Forward | GTTGAAGGTCGCTTGCCG | 60 | 118 | [53] |
| Reverse | CCAAGGTATGGGCACCAG | ||||
| sod | Forward | GCCACTGCCTCTGCTAATTCA | 60 | 104 | [52] |
| Reverse | CCAAATTGTTTCTTTTGGGTTCTCC | ||||
| aao | Forward | CCAGGCACAAACACAATCAA | 60 | 154 | [54] |
| Reverse | GTCGTAAATAATATCAGACTG | ||||
| opr3 | Forward | ATGGACTCTAATCCACTCAGCCTTG | 60 | 152 | [55] |
| Reverse | TCACTGCCAAGTCTGCCTGCTTCAG |
| Sample | Time (Days) | Size (nm) | SD | PDI | SD | Zeta-Potential | SD |
|---|---|---|---|---|---|---|---|
| Zein NPs | 0 | 142.3 | 4.88 | 0.222 | 0.025 | 30.3 | 1.01 |
| 7 | 132.9 | 7.53 | 0.364 | 0.034 | 28.8 | 1.41 | |
| 30 | 122.7 | 11.87 | 0.361 | 0.024 | 28.9 | 0.451 | |
| 60 | 128.8 | 9.383 | 0.324 | 0.008 | 30.1 | 0.764 | |
| 90 | 122.1 | 7.804 | 0.261 | 0.02 | 29.8 | 2.04 | |
| EO-Zein NPs | 0 | 159.1 | 3.99 | 0.338 | 0.108 | 30.3 | 1.13 |
| 7 | 152.5 | 5.233 | 0.254 | 0.017 | 26.9 | 2.112 | |
| 30 | 149.3 | 3.782 | 0.232 | 0.003 | 28.3 | 2.4 | |
| 60 | 151.7 | 3.755 | 0.218 | 0.01 | 30.5 | 0.721 | |
| 90 | 150.5 | 3.922 | 0.211 | 0.007 | 30.1 | 0.764 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira-Pinto, P.R.; Mariz-Ponte, N.; Sousa, R.M.O.F.; Torres, A.; Tavares, F.; Ribeiro, A.; Cavaco-Paulo, A.; Fernandes-Ferreira, M.; Santos, C. Satureja montana Essential Oil, Zein Nanoparticles and Their Combination as a Biocontrol Strategy to Reduce Bacterial Spot Disease on Tomato Plants. Horticulturae 2021, 7, 584. https://doi.org/10.3390/horticulturae7120584
Oliveira-Pinto PR, Mariz-Ponte N, Sousa RMOF, Torres A, Tavares F, Ribeiro A, Cavaco-Paulo A, Fernandes-Ferreira M, Santos C. Satureja montana Essential Oil, Zein Nanoparticles and Their Combination as a Biocontrol Strategy to Reduce Bacterial Spot Disease on Tomato Plants. Horticulturae. 2021; 7(12):584. https://doi.org/10.3390/horticulturae7120584
Chicago/Turabian StyleOliveira-Pinto, Paulo R., Nuno Mariz-Ponte, Rose Marie O. F. Sousa, Ana Torres, Fernando Tavares, Artur Ribeiro, Artur Cavaco-Paulo, Manuel Fernandes-Ferreira, and Conceição Santos. 2021. "Satureja montana Essential Oil, Zein Nanoparticles and Their Combination as a Biocontrol Strategy to Reduce Bacterial Spot Disease on Tomato Plants" Horticulturae 7, no. 12: 584. https://doi.org/10.3390/horticulturae7120584
APA StyleOliveira-Pinto, P. R., Mariz-Ponte, N., Sousa, R. M. O. F., Torres, A., Tavares, F., Ribeiro, A., Cavaco-Paulo, A., Fernandes-Ferreira, M., & Santos, C. (2021). Satureja montana Essential Oil, Zein Nanoparticles and Their Combination as a Biocontrol Strategy to Reduce Bacterial Spot Disease on Tomato Plants. Horticulturae, 7(12), 584. https://doi.org/10.3390/horticulturae7120584