
Heat Treatment in Two Tomato Cultivars: A Study of the Effect on Physiological and Growth Recovery

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Heat Treatment Conditions
2.2. Measurement of Heat Tolerance among Tomato Seedlings
2.3. Measurement of Chlorophyll Contents and Photosynthetic Rate in Seedlings under Heat Treatment
2.4. Determination of Electrolyte Leakage Potential in Seedlings Leaves under HT
2.5. Extraction of Free Total Proline Content in Seedlings Leaves under HT
2.6. Proline Content and Seedlings Growth with Different Leaf Damage Levels at Recovery
2.7. Plant Materials and Heat Treatment Conditions
2.8. Data Collection on Reproductive Parameters at Growth Period
2.9. Statistical Analysis
3. Results
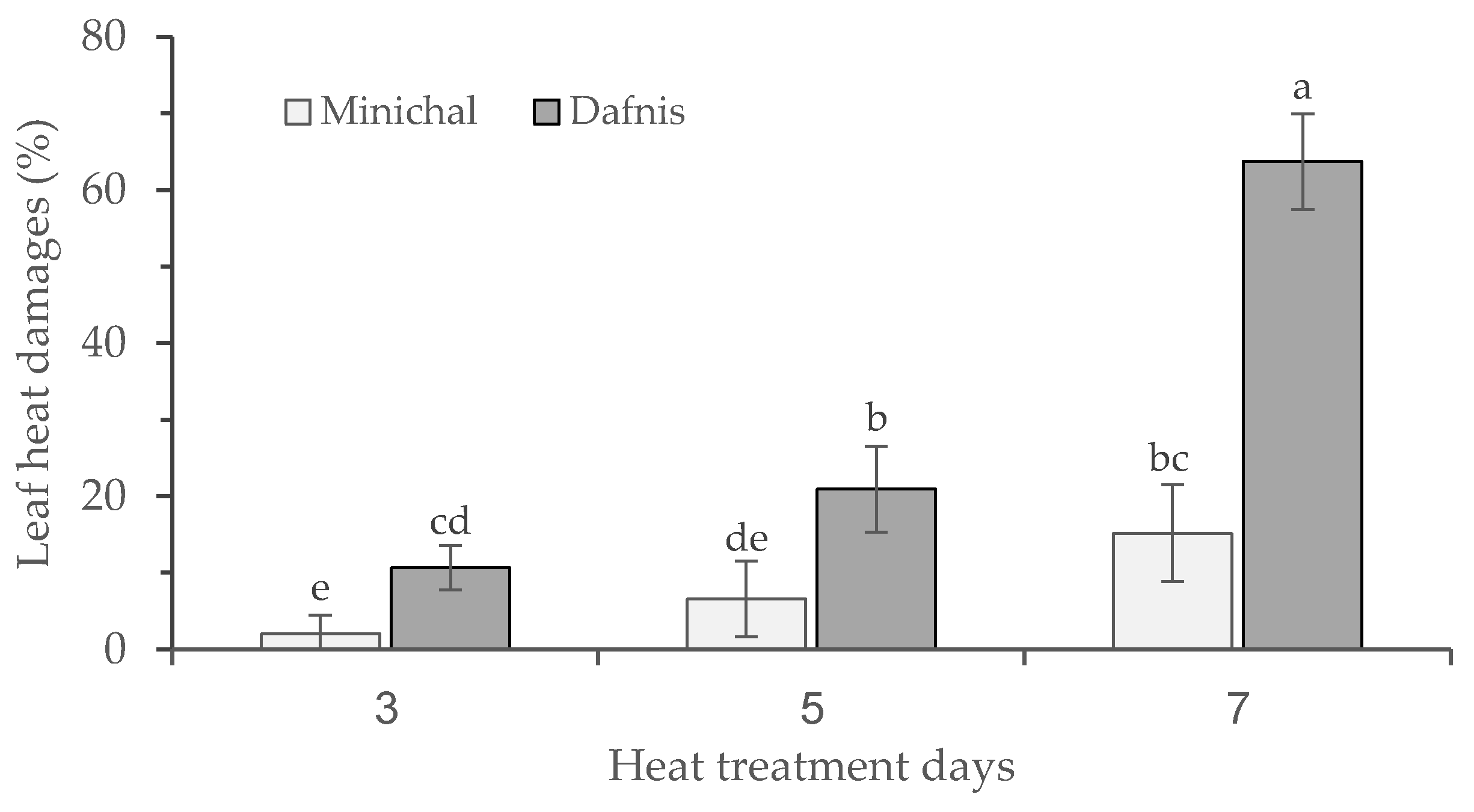
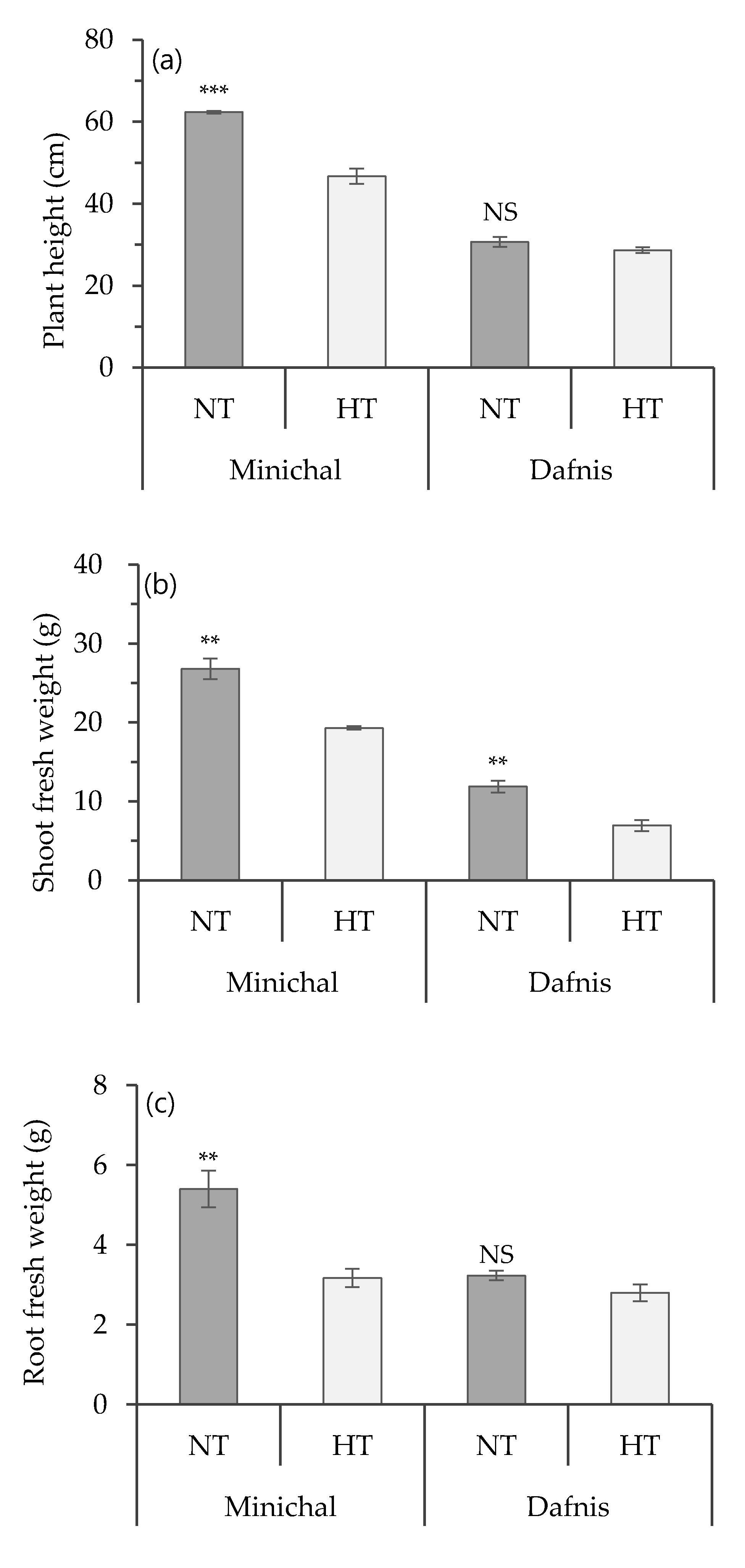
3.1. Screening for Heat Tolerance in Tomato Seedlings
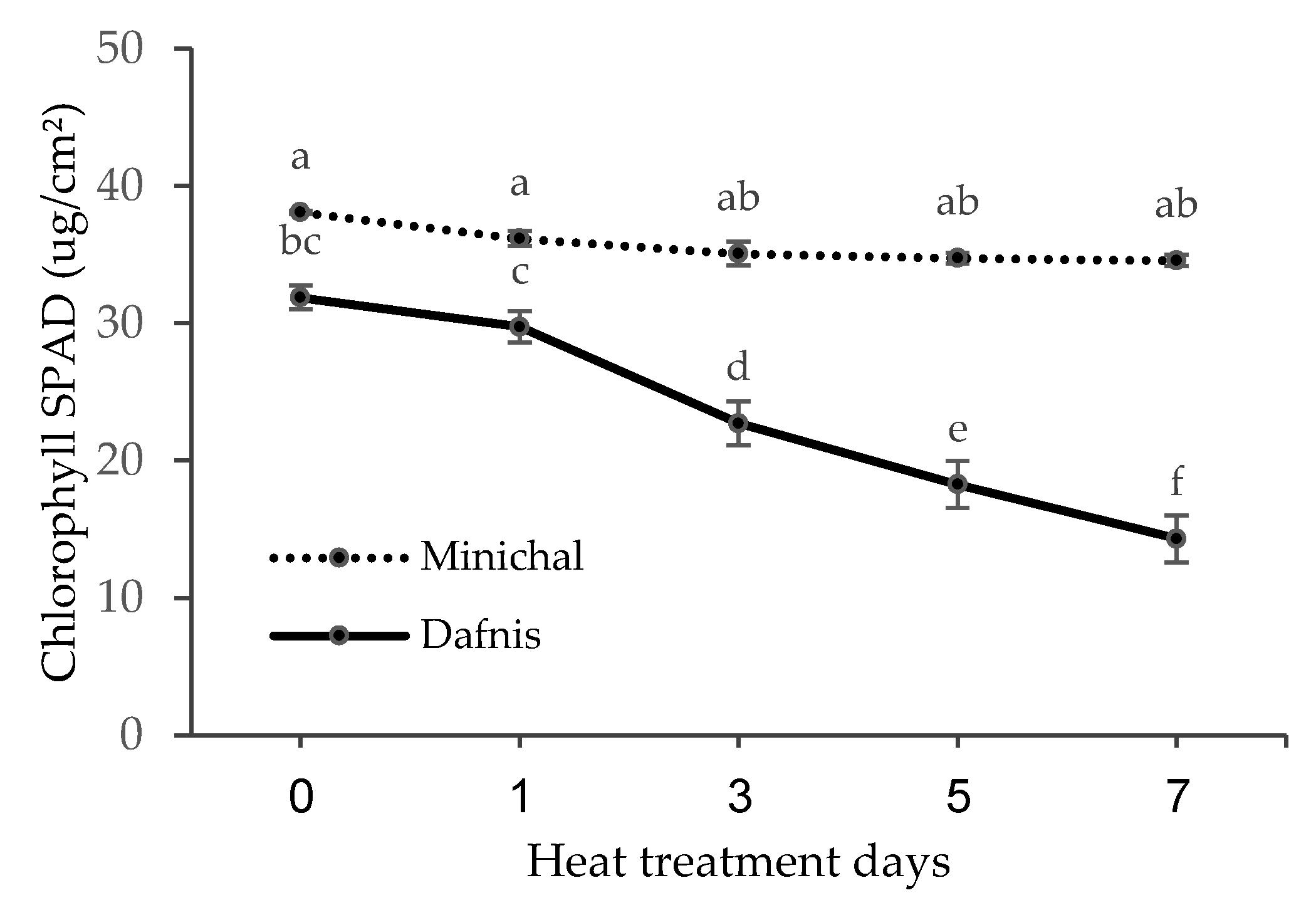
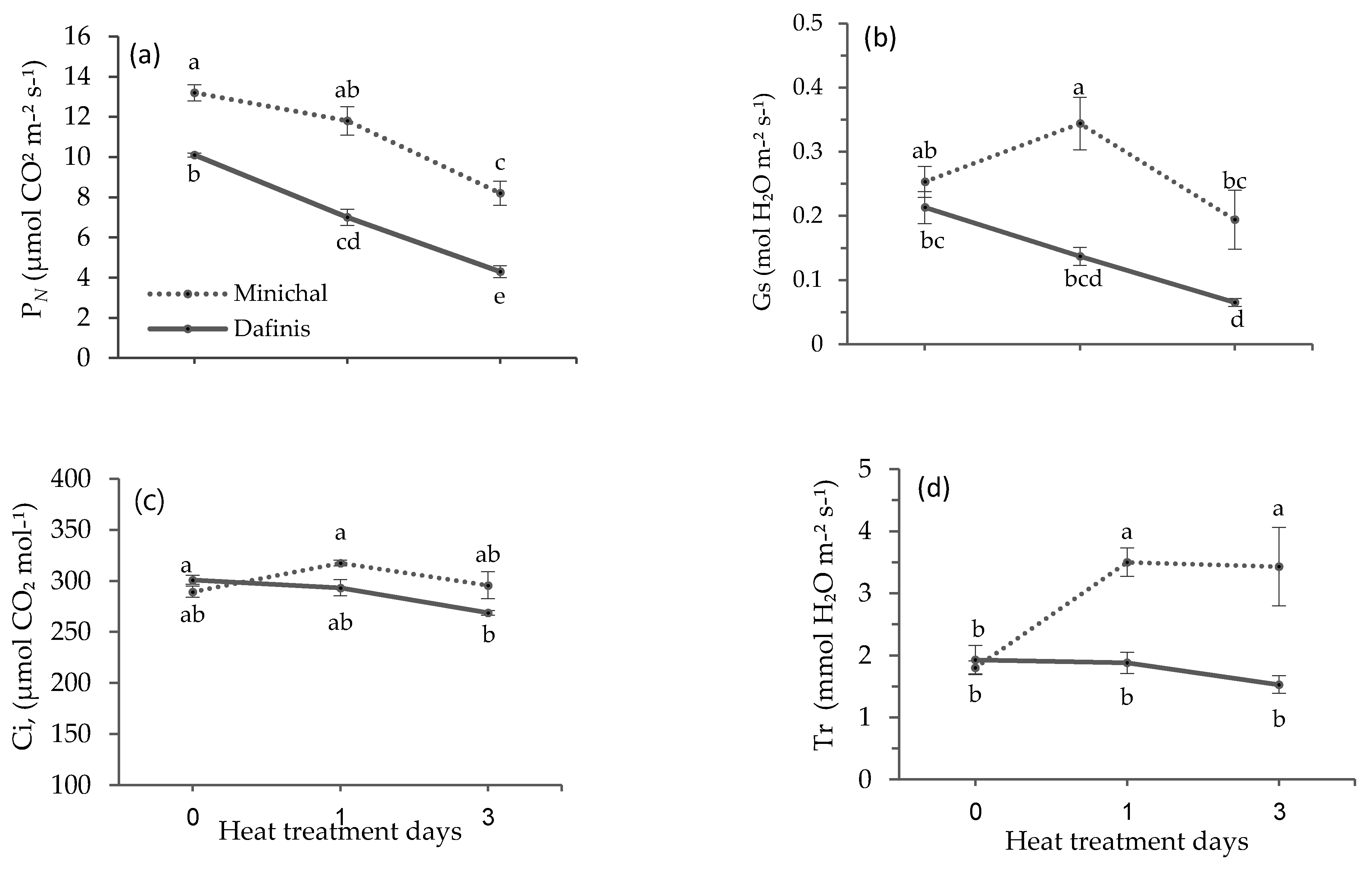
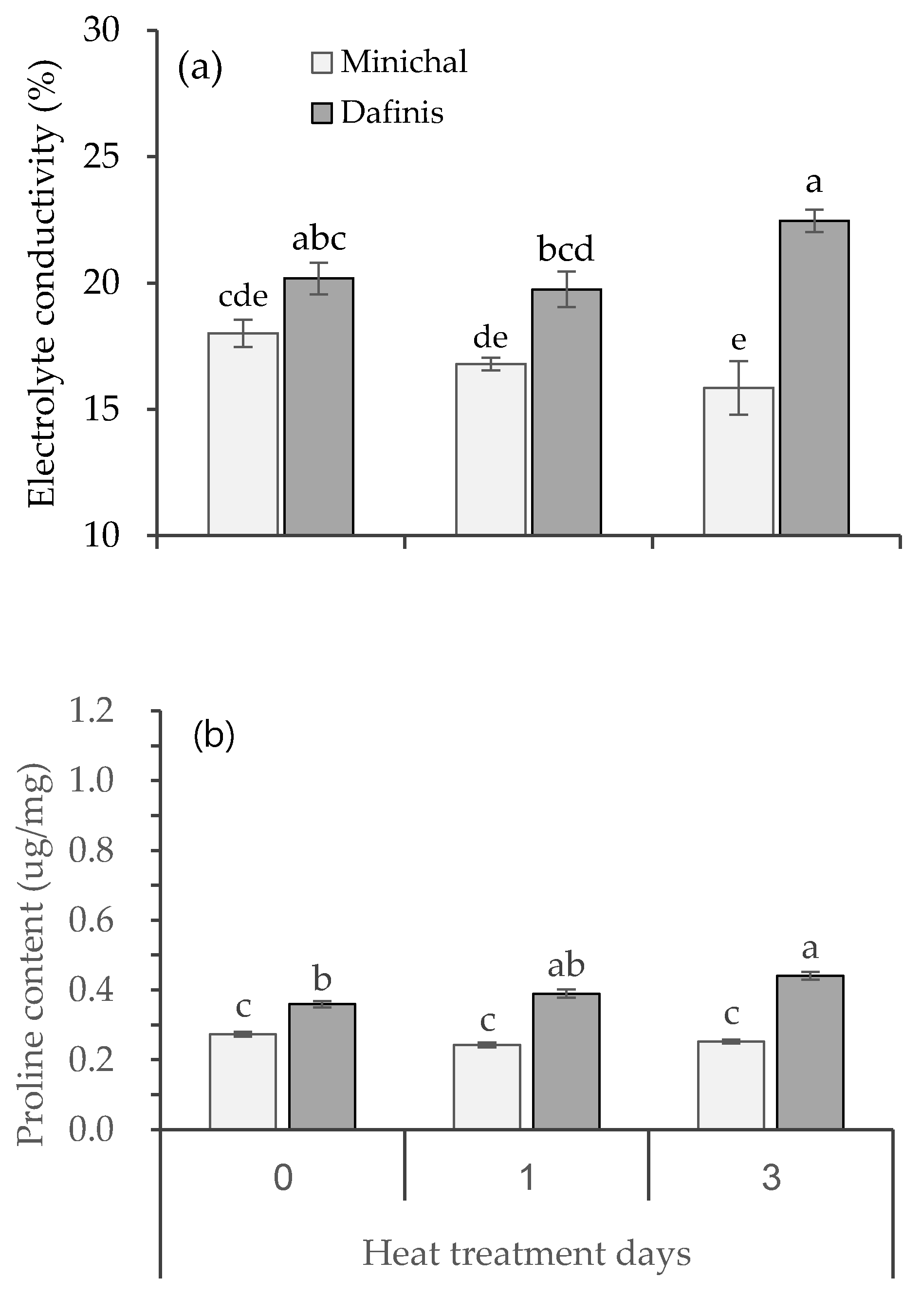
3.2. The Difference in Physiological Responses to High Temperature between Heat-Susceptible and Tolerant Seedlings
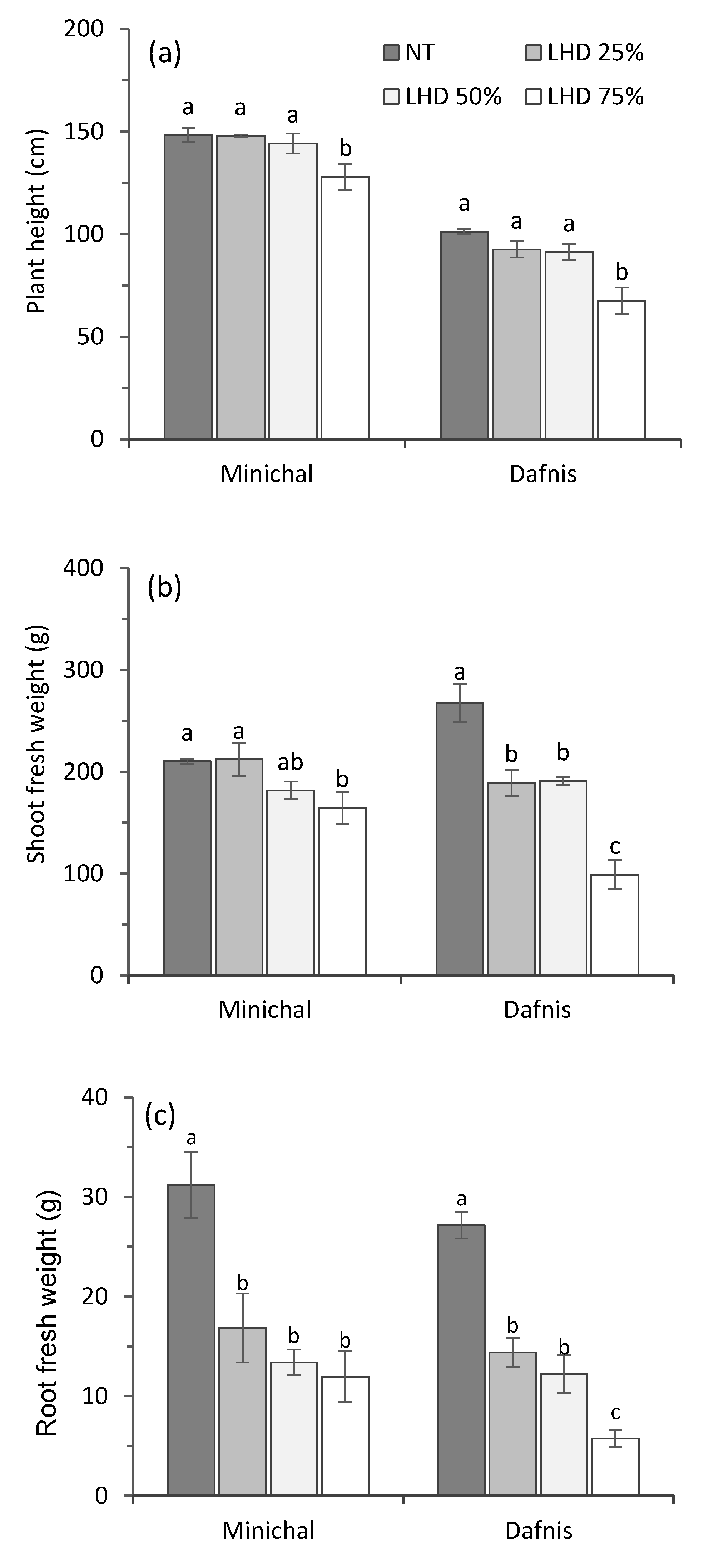
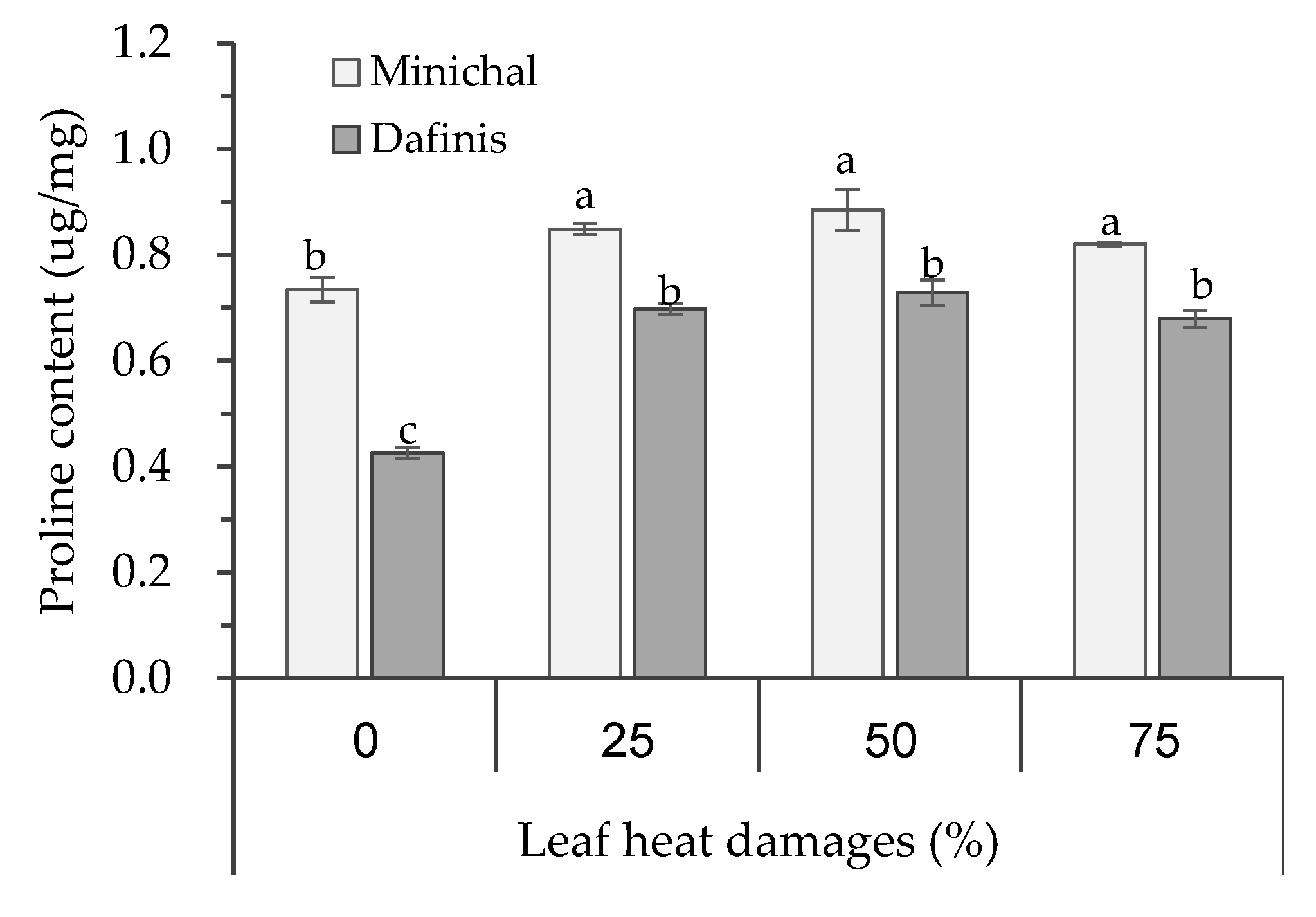
3.3. Effects of Leaf Heat Damage Levels on the Growth and Proline Content of Heat-Susceptible and Tolerant Cultivars at Recovery
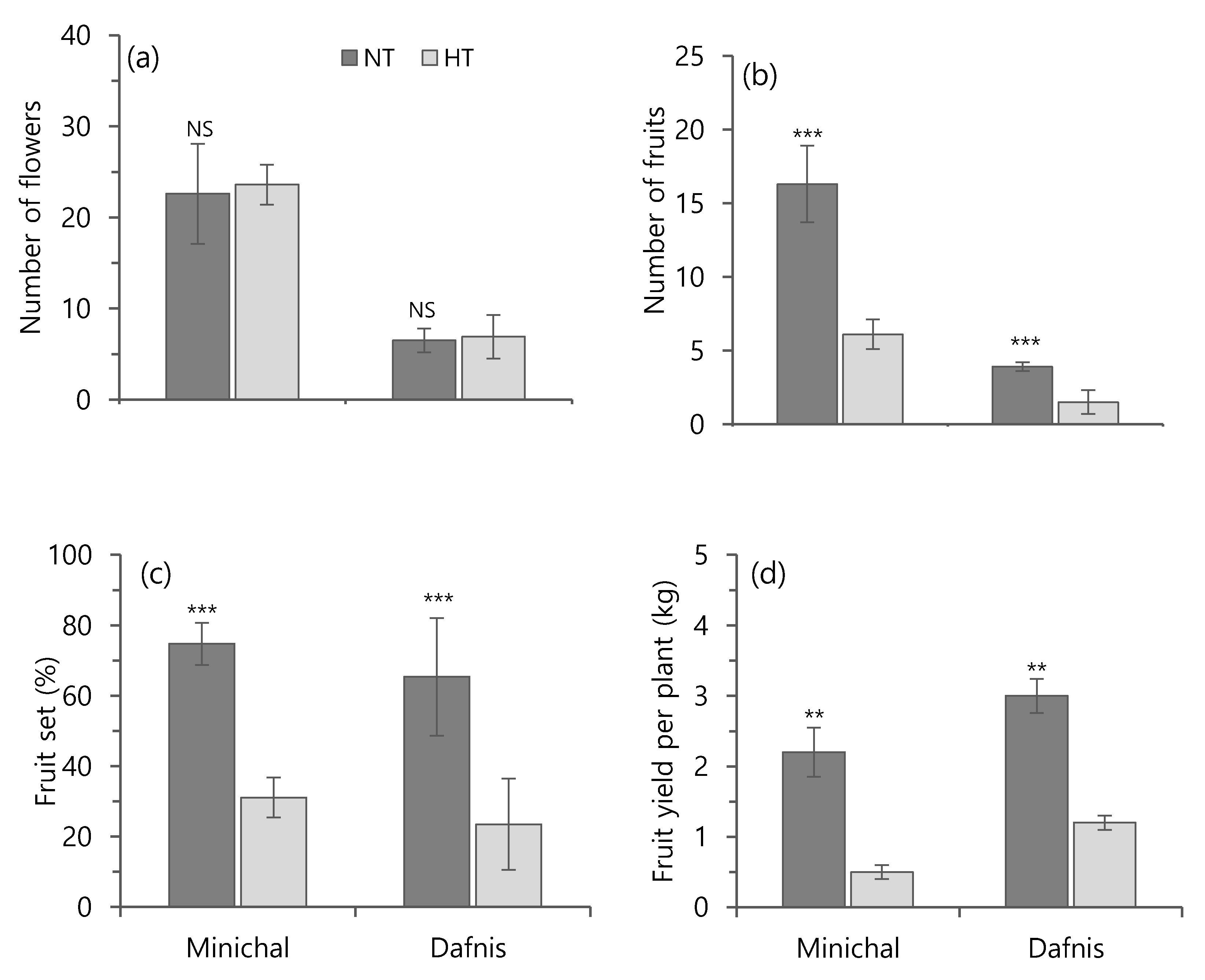
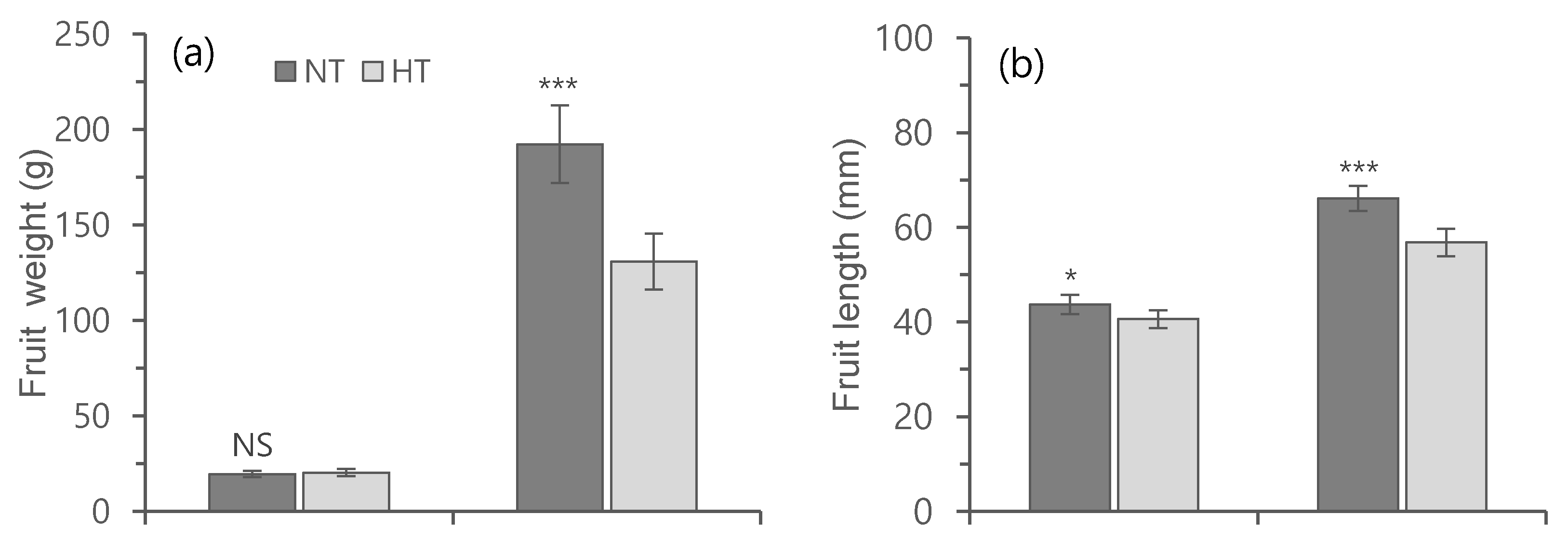
3.4. Effect of Heat Treatment on the Development of Flowers, Fruit Parameters, and Yield of Heat-Susceptible and Tolerant Cultivars
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Camejo, D.; Rodríguez, P.; Angeles Morales, M.; Dell’Amico, J.M.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. Plant Physiol. 2005, 162, 281–2896. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Wu, Z.; Wang, X.; Rosenqvist, E.; Wang, Y.; Zhao, T.; Ottosen, C.O. Evaluation of temperature stress tolerance in cultivated and wild tomatoes using photosynthesis and chlorophyll fluorescence. Hortic. Environ. Biotechnol. 2018, 59, 499–509. [Google Scholar] [CrossRef]
- Camejo, D.; Jiménez, A.; Alarcón, J.J.; Torres, W.; Gómez, J.M.; Sevilla, F. Changes in photosynthetic parameters and antioxidant activities following heat-shock treatment in tomato plants. Funct. Plant Biol. 2006, 33, 177–187. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Kjær, K.H.; Rosenqvist, E.; Ottosen., C.O.; Wu, Z. Screening and validation of tomato genotypes under heat stress using Phenotyping tomato for heat tolerance Functional Plant Biology K Fv/Fm to reveal the physiological mechanism of heat tolerance. Environ. Exp. Bot. 2015, 118, 1–11. [Google Scholar] [CrossRef]
- Hu, Y.; Wu, Q.; Sprague, S.A.; Park, J.; Oh, M.; Rajashekar, C.B.; Koiwa, H.; A Nakata, P.; Cheng, N.; Hirschi, K.D.; et al. Tomato expressing Arabidopsis glutaredoxin gene AtGRXS17 confers tolerance to chilling stress via modulating cold responsive components. Hortic. Res. 2015, 2, 15051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Chen, X.; Chen, D.; Li, J.; Zhang, Y.; Wang, A. A comparison of the low temperature transcriptomes of two tomato genotypes that differ in freezing tolerance: Solanum lycopersicum and Solanum habrochaites. J. BMC Plant Biol. 2015, 15, 132. [Google Scholar] [CrossRef] [Green Version]
- Rajametov, S.; Yang, E.Y.; Cho, M.C.; Chae, S.Y.; Kim, J.H.; Nam, C.W.; Chae, W.B. Traits Affecting Low Temperature Tolerance in Tomato and its Application to Breeding Program. Plant Breed. Biotech. 2019, 7, 350–359. [Google Scholar] [CrossRef]
- Bolger, A.; Scossa, F.; Bolger, M.E.; Lanz, C.; Maumus, F.; Tohge, T.; Quesneville, H.; Alseekh, S.; Sørensen, I.; Lichtenstein, G.; et al. The genome of the stress-tolerant wild tomato species Solanum pennellii. Nat. Genet. 2014, 46, 1034–1038. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, X.D.; Li, T.L.; Cao, X.J. Photosynthesis and ultrastructure of photosynthetic apparatus in tomato leaves under elevated temperature. Photosynthetica 2014, 52, 430–436. [Google Scholar] [CrossRef]
- Poudyal, D.; Rosenqvist, E.; Ottosen, C.-O. Phenotyping from lab to field–tomato lines screened for heat stress using Fv/Fm maintain high fruit yield during thermal stress in the field. Funct. Plant Biol. 2019, 46, 44–55. [Google Scholar] [CrossRef]
- Zhou, R.; Kjaer, K.H.; Rosenqvist, E.; Yu, X.; Wu, Z.; Ottosen, C.O. Physiological response to heat stress during seedling and anthesis stage in tomato genotypes differing in heat tolerance. Agron. Crop Sci. 2016, 203, 68–80. [Google Scholar] [CrossRef]
- Zhou, R.; Kong, L.; Wu, Z.; Rosenqvist, E.; Wang, Y.; Zhao, L.; Zhao, T.; Ottosen, C. Physiological response of tomatoes at drought, heat and their combination followed by recovery. Physiol. Plant. 2019, 165, 144–154. [Google Scholar] [CrossRef]
- Abdul-Baki, A.A.; Stommel, J.R. Pollen viability and fruit set of tomato genotypes under optimum and high-temperature regimes. Hortic. Sci. 1995, 30, 115–117. [Google Scholar]
- Xu, J.; Wolters-Arts, M.; Mariani, C.; Huber, H.; Rieu, I. Heat stress affects vegetative and reproductive performance and trait correlations in tomato (Solanum lycopersicum). Euphytica 2017, 213, 156. [Google Scholar] [CrossRef] [Green Version]
- Rajametov, S.; Yang, E.Y.; Cho, M.C.; Chae, S.Y.; Chae, W.B. Physiological traits associated with high temperature tolerance differ by fruit types and sizes in tomato (Solanum lycopersicum L.). Hortic. Environ. Biotechnol. 2020, 61, 837–847. [Google Scholar] [CrossRef]
- Ro, S.; Chea, L.; Ngoun, S.; Stewart, Z.; Roeurn, S.; Theam, P.; Lim, S.; Sor, R.; Kosal, M.; Roeun, M.; et al. Response of tomato genotypes under different high temperatures in field and greenhouse conditions. Plants 2021, 10, 449. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Ottosen, C.-O.; Rosenqvist, E.; Zhao, L.; Wang, Y.; Yu, W.; Zhao, T.; Wu, Z. Drought stress had a predominant effect over heat stress on three tomato cultivars subjected to combined stress. BMC Plant Biol. 2017, 17, 24. [Google Scholar] [CrossRef] [Green Version]
- Rieu, I.; Twel, D.; Firon, N. Pollen development at high temperature: From acclimation to collapse. Plant Physiol. 2017, 173, 1967–1976. [Google Scholar] [CrossRef] [Green Version]
- Shah Jahan, M.; Wang, Y.; Sheng Sh Zhong, M.; Chen, Z.; Wu, J.; Sun, J.; Guo, S. Exogenous salicylic acid increases the heat tolerance in Tomato (Solanum lycopersicum L) by enhancing photosynthesis efficiency and improving antioxidant defense system through scavenging of reactive oxygen species. Sci. Hortic. 2019, 247, 421–429. [Google Scholar] [CrossRef]
- Claussen, W. Proline as a measure of stress in tomato plants. Plant Sci. 2005, 168, 241–248. [Google Scholar] [CrossRef]
- Korea Soil Information System. Available online: https://soil.rda.go.kr (accessed on 14 May 2021).
- Karapanos, I.C.; Akoumianakis, K.A.; Olympio, C.M.; Harold, C.P. Tomato pollen respiration in relation to in vitro germination and pollen tube growth under favorable and stress-inducing temperatures. Sex Plant Reprod. 2010, 23, 219–224. [Google Scholar] [CrossRef]
- Golam, F.; Prodhan, Z.H.; Nezhadahmadi, A.; Rahman, M. Heat tolerance in tomato. Life Sci. J. 2012, 9, 1936–1950. [Google Scholar]
- Din, J.U.; Khan, S.U.; Khan, A.; Qayyum, A.; Abbasi, K.S.; Jenks, M.A. Evaluation of potential morpho-physiological and biochemical indicators in selecting heat-tolerant tomato (Solanum lycopersicum Mill.) genotypes. Hortic. Environ. Biotechnol. 2015, 56, 769–776. [Google Scholar] [CrossRef]
- Fan, X.X.; Xu, Z.G.; Liu, X.Y.; Tang, C.M.; Wang, L.W.; Han, X. Effects of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Sci. Hortic. 2013, 153, 50–55. [Google Scholar] [CrossRef]
- Morphological, Physiological and Biochemical Responses of Gerbera Cultivars to Heat Stress. Korean J. Hortic. Sci. 2016, 34, 1–14. [CrossRef]
- Lu, T.; Meng, Z.; Zhang, G.; Qi, M.; Sun, Z.; Liu, Y.; Li, T. Sub-high Temperature and High Light Intensity Induced Irreversible Inhibition on Photosynthesis System of Tomato Plant (Solanum lycopersicum L.). Front Plant Sci. 2017, 8, 365. [Google Scholar] [CrossRef] [Green Version]
- Herde, O.; Pena-Cortes, H.; Fuss, H.; Willmitzer, L.; Fisahn, J. Effects of mechanical wounding, current application and heat treatment on chlorophyll fluorescence and pigment composition in tomato plants. Physiol. Plant. 1999, 105, 179–184. [Google Scholar] [CrossRef]
- Vijayalakshmi, K.; Fritz, A.K.; Paulsen, G.M.; Bai, G.; Pandravada, S.; Gill, B.S. Modeling and mapping QTL for senescence-related traits in winter wheat under high temperature. Mol. Breed. 2010, 26, 163–175. [Google Scholar] [CrossRef]
- Gao, Y.; Guo, Y.K.; Lin, S.H.; Fang, Y.Y.; Bai, J.G. Hydrogen peroxide pretreatment alters the activity of antioxidant enzymes and protects chloroplast ultrastructure in heat-stressed cucumber leaves. Sci. Hortic. 2010, 126, 20–26. [Google Scholar] [CrossRef]
- Nankishore, A.; Farrell, A.D. The response of contrasting tomato genotypes to combined heat and drought stress. J. Plant Physiol. 2016, 202, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Yuan, L.; Zhang, S.; Wang, J.; Xie, S.; Zhao, M.; Chen, G.; Sun, R.; Wang, C. Comprehensive Evaluation for Cold Tolerance in Wucai (Brassica campestris L.) by the Performance Index on an Absorption Basis (PIabs). Agronomy 2019, 9, 61. [Google Scholar] [CrossRef] [Green Version]
- Alsamir, M.; Ahmad, N.M.; Keitel, C.; Mahmood, T.; Trethowan, R. Identification of high-temperature tolerant and agronomically viable tomato (S. lycopersicum) genotypes from a diverse germplasm. Adv. Crop Sci. Tech. 2017, 5, 299. [Google Scholar] [CrossRef]
- Hong, Z.L.; Lakkineni, K.; Zhang, Z.M.; Verma, D.P.S. Removal of feedback inhibition of D(1)-pyrroline-5-carboxylate synthetase results in increased proline accumulation and protection of plants from osmotic stress. Plant Physiol. 2000, 122, 1129–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harsh, A.; Sharma, Y.K.; Joshi, U.; Rampuria, S.; Singh, G.; Kumar, S.; Sharma, R. Effect of short-term heat stress on total sugars, proline and some antioxidant enzymes in moth bean (Vigna aconitifolia). Ann. Agric. Sci. 2016, 61, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Rivero, R.M.; Ruiz, J.M.; Romero, L. Importance of N source on heat stress tolerance due to the accumulation of proline and quaternary ammonium compounds in tomato plants. Plant Biol. 2004, 6, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, S.R.; Kim, Y.H.; Lee, J.G. Detection of temperature stress using chlorophyll fluorescence parameters and stress-related chlorophyll and proline content in paprika (Capsicum annuum L.) seedlings. Korean J. Hortic. Sci. Technol. 2018, 36, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, J.M.; Sanchez, E.; Garcia, P.C.; Lopez-Lefebre, L.R.; Rivero, R.M.; Romero, L. Proline metabolism and NAD kinase activity in greenbean plants subjected to cold-shock. Phytochemistry 2002, 59, 473. [Google Scholar] [CrossRef]
- Gosavi, G.U.; Jadhav, A.S.; Kale, A.A.; Gadakh, S.R.; Pawar, B.D.; Chimote, V.P. Effect of heat stress on proline, chlorophyll content, heat shock proteins and antioxidant enzyme activity in sorghum (Sorghum bicolor) at seedlings stage. Indian J. Biotechnol. 2014, 13, 356–363. [Google Scholar]
- AVRDC. 2017. Available online: https://avrdc.org/seed/improved-lines/processingdual-purpose-tomato/ (accessed on 19 August 2020).
- Solankey, S.S.; Akhtar, S.; Neha, P.; Kumari, M.; Kherwa, R. Screening and identification of heat tolerant tomato genotypes for Bihar. J. Pharm. Phytochem. 2018, 7, 97–100. [Google Scholar]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajametov, S.N.; Yang, E.Y.; Jeong, H.B.; Cho, M.C.; Chae, S.Y.; Paudel, N. Heat Treatment in Two Tomato Cultivars: A Study of the Effect on Physiological and Growth Recovery. Horticulturae 2021, 7, 119. https://doi.org/10.3390/horticulturae7050119
Rajametov SN, Yang EY, Jeong HB, Cho MC, Chae SY, Paudel N. Heat Treatment in Two Tomato Cultivars: A Study of the Effect on Physiological and Growth Recovery. Horticulturae. 2021; 7(5):119. https://doi.org/10.3390/horticulturae7050119
Chicago/Turabian StyleRajametov, Sherzod Nigmatullaevich, Eun Young Yang, Hyo Bong Jeong, Myeong Cheoul Cho, Soo Young Chae, and Niroj Paudel. 2021. "Heat Treatment in Two Tomato Cultivars: A Study of the Effect on Physiological and Growth Recovery" Horticulturae 7, no. 5: 119. https://doi.org/10.3390/horticulturae7050119
APA StyleRajametov, S. N., Yang, E. Y., Jeong, H. B., Cho, M. C., Chae, S. Y., & Paudel, N. (2021). Heat Treatment in Two Tomato Cultivars: A Study of the Effect on Physiological and Growth Recovery. Horticulturae, 7(5), 119. https://doi.org/10.3390/horticulturae7050119