
Wild Species Veronica officinalis L. and Veronica saturejoides Vis. ssp. saturejoides—Biological Potential of Free Volatiles

 ,
,  , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. GC and GC-MS Analyses
2.3. Micromorphological Traits
2.4. Antiphytoviral Activity
2.4.1. Virus and Plant Hosts
2.4.2. Antiphytoviral Activity Assay
2.5. Cell Culture
2.6. Cell Proliferation Assay
2.7. Antioxidant Activity of Essential Oils and Hydrosols
2.7.1. ORAC
2.7.2. DPPH
2.8. Statistical Analyses
3. Results and Discussion
3.1. Composition of Free Volatile Components
3.2. Glandular and Non-Glandular Trichomes
3.3. Antiphytoviral Activity
3.4. Antiproliferative Activity
3.5. Antioxidant Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Albach, D.C.; Martinez-Ortega, M.M.; Chase, M.W. Veronica: Parallel morphological evolution and phylogeography in the Mediterranean. Plant Syst. Evol. 2004, 246, 177–194. [Google Scholar] [CrossRef]
- Albach, D.C.; Martínez-Ortega, M.M.; Fischer, M.A.; Chase, M.W. A new classification of the tribe Veroniceae-problems and a possible solution. Taxon 2004, 53, 429–452. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Tayeboon, G.S.; Sharifi-Rad, J.; Iriti, M.; Varoni, E.M.; Razazi, S. Inhibitory activity on type 2 diabetes and hypertension key-enzymes, and antioxidant capacity of Veronica persica phenolic-rich extracts. Cell. Mol. Biol. 2016, 62, 80–85. [Google Scholar] [CrossRef]
- Nikolić, T. Flora Croatica Database. Available online: http://hirc.botanic.hr/fcd (accessed on 22 May 2021).
- Reynolds, T. The evolution of chemosystematics. Phytochemistry 2007, 68, 2887–2895. [Google Scholar] [CrossRef] [PubMed]
- Loreto, F.; Dicke, M.; Schnitzler, J.P.; Turlings, T.C.J. Plant volatiles and the environment. Plant Cell Environ. 2014, 37, 1905–1908. [Google Scholar] [CrossRef]
- Salehi, B.; Shetty, M.S.; Anil Kumar, N.V.; Živković, J.; Calina, D.; Docea, A.O.; Emamzadeh-Yazdi, S.; Kılıç, C.S.; Goloshvili, T.; Nicola, S.; et al. Veronica plants—Drifting from farm to traditional healing, food application, and phytopharmacology. Molecules 2019, 24, 2454. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.; Chen, K.X.; Zhang, L.Q.; Li, Y.M. Review of the Ethnopharmacology, Phytochemistry, and Pharmacology of the Genus Veronica. Am. J. Chin. Med. 2019, 47, 1193–1221. [Google Scholar] [CrossRef]
- Dunkić, V.; Kosalec, I.; Kosir, I.; Potočnik, T.; Cerenak, A.; Koncic, M.; Vitali, D.; Muller, I.; Kopricanec, M.; Bezić, N.; et al. Antioxidant and antimicrobial properties of Veronica spicata L. (Plantaginaceae). Curr. Drug Targets 2015, 16, 1660–1670. [Google Scholar] [CrossRef]
- Ertas, A.; Boga, M.; Kizil, M.; Ceken, B.; Goren, A.C.; Hasimi, N.; Demirci, S.; Topcu, G.; Kolak, U. Chemical profile and biological activities of Veronica thymoides subsp. pseudocinerea. Pharm. Biol. 2015, 53, 334–339. [Google Scholar] [CrossRef] [Green Version]
- Mocan, A.; Vodnar, D.C.; Vlase, L.; Crișan, O.; Gheldiu, A.M.; Crișan, G. Phytochemical characterization of Veronica officinalis L., V. teucrium L. and V. orchidea Crantz from Romania and their antioxidant and antimicrobial properties. Int. J. Mol. Sci. 2015, 16, 21109–21127. [Google Scholar] [CrossRef]
- Valyova, M.; Hadjimitova, V.; Stoyanov, S.; Ganeva, Y.; Petkov, I. Free radical scavenging activity of extracts from Bulgarian Veronica officinalis L. and GC-MS analysis of ethanol extract. Internet J. Aesthetic Antiaging Med. 2008, 2, 2–6. [Google Scholar] [CrossRef]
- Kwak, J.H.; Kim, H.J.; Lee, K.H.; Kang, S.C.; Zee, O.P. Antioxidative iridoid glycosides and phenolic compounds from Veronica peregrina. Arch. Pharm. Res. 2009, 32, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Ishikawa, M. Replication of Tobamovirus RNA. Annu. Rev. Phytopathol. 2016, 54, 55–78. [Google Scholar] [CrossRef] [Green Version]
- Othman, B.A.; Shoman, S.A. Antiphytoviral Activity of the Plectranthus tenuiflorus on Some Important Viruses. Int. J. Agric. Biol. 2004, 6, 844–849. [Google Scholar]
- Rusak, G.; Krajačić, M.; Krsnik-Rasol, M.; Gutzeit, H.O. Quercetin influences response in Nicotiana megalosiphon infected by satellite-associated cucumber mosaic virus/Quercetin beeinf lusst die Reaktion von Nikotiana magalosiphon gegenüber Satelliten-assoziierten Gurkenmosaikvirus-Infektionen. J. Plant Dis. Prot. 2007, 114, 145–150. [Google Scholar] [CrossRef]
- Krcatović, E.; Rusak, G.; Bezić, N.; Krajačić, M. Inhibition of tobacco mosaic virus infection by quercetin and vitexin. Acta Virol. 2008, 52, 119–124. [Google Scholar]
- Moeini, A.; van Reenen, A.; Van Otterlo, W.; Cimmino, A.; Masi, M.; Lavermicocca, P.; Valerio, F.; Immirzi, B.; Santagata, G.; Malinconico, M.; et al. α-costic acid, a plant sesquiterpenoid from Dittrichia viscosa, as modifier of Poly (lactic acid) properties: A novel exploitation of the autochthone biomass metabolite for a wholly biodegradable system. Ind. Crops Prod. 2020, 146, 112134. [Google Scholar] [CrossRef]
- Vuko, E.; Dunkić, V.; Bezić, N.; Ruščić, M.; Kremer, D. Chemical Composition and Antiphytoviral Activity of Essential Oil of Micromeria graeca. Nat. Prod. Commun. 2012, 7, 1227–1230. [Google Scholar] [CrossRef] [Green Version]
- Dunkić, V.; Bezić, N.; Vuko, E.; Cukrov, D. Antiphytoviral activity of Satureja montana L. ssp. variegata (host) P. W. Ball essential oil and phenol compounds on CMV and TMV. Molecules 2010, 15, 6713–6721. [Google Scholar] [CrossRef] [Green Version]
- Dunkić, V.; Bezić, N.; Vuko, E. Antiphytoviral activity of essential oil from endemic species Teucrium arduini. Nat. Prod. Commun. 2011, 6, 1385–1388. [Google Scholar] [CrossRef] [Green Version]
- Dunkić, V.; Vuko, E.; Bezić, N.; Kremer, D.; Ruščić, M. Composition and antiviral activity of the essential oils of Eryngium alpinum and E. amethystinum. Chem. Biodivers. 2013, 10, 1894–1902. [Google Scholar] [CrossRef] [PubMed]
- Bezić, N.; Vuko, E.; Dunkić, V.; Ruščić, M.; Blažević, I.; Burčul, F. Antiphytoviral Activity of Sesquiterpene-Rich Essential Oils from Four Croatian Teucrium species. Molecules 2011, 16, 8119–8129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuko, E.; Rusak, G.; Dunkić, V.; Kremer, D.; Kosalec, I.; Rada, B.; Bezić, N. Inhibition of satellite RNA associated cucumber mosaic virus infection by essential oil of Micromeria croatica (Pers.) Schott. Molecules 2019, 24, 1342. [Google Scholar] [CrossRef] [Green Version]
- Vuko, E.; Dunkić, V.; Ruščić, M.; Nazlić, M.; Mandić, N.; Soldo, B.; Šprung, M.; Fredotović, Ž. Chemical Composition and New Biological Activities of Essential Oil and Hydrosol of Hypericum perforatum L. ssp. veronense (Schrank) H. Lindb. Plants 2021, 10, 1014. [Google Scholar] [CrossRef]
- Lu, M.; Han, Z.; Xu, Y.; Yao, L. In Vitro and In Vivo Anti-Tobacco Mosaic Virus Activities of Essential Oils and Individual Compounds. J. Microbiol. Biotechnol. 2013, 23, 771–778. [Google Scholar] [CrossRef] [Green Version]
- Bishop, C.D. Antiviral Activity of the Essential Oil of Melaleuca alternifolia (Maiden amp; Betche) Cheel (Tea Tree) Against Tobacco Mosaic Virus. J. Essent. Oil Res. 1995, 7, 641–644. [Google Scholar] [CrossRef]
- Nazlić, M.; Kremer, D.; Jurišić-Grubešić, R.; Soldo, B.; Vuko, E.; Stabentheiner, E.; Ballian, D.; Bogunić, F.; Dunkić, V. Endemic Veronica saturejoides Vis. ssp. saturejoides–Chemical Composition and Antioxidant Activity of Free Volatile Compounds. Plants 2020, 9, 1646. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing: Carol Stream, IL, USA, 2017; ISBN 978-1-932633-21-4. [Google Scholar]
- Babushok, V.I.; Linstrom, P.J.; Zenkevich, I.G. Retention Indices for Frequently Reported Compounds of Plant Essential Oils. J. Phys. Chem. Ref. Data 2011, 40, 1–47. [Google Scholar] [CrossRef] [Green Version]
- Li, F. Analysis of chemical constituents of essential oil in Veronica linariifolia by gas chromatography-mass spectrometry. Chin. J. Anal. Chem. 2002, 30, 822–825. [Google Scholar]
- Çelik, E.; Çelik, G.Y.; Meysun, A. Essential Oil Composition and Antibacterial Activity of Some Plant Species. J. Appl. Biol. Sci. 2010, 1, 45–48. [Google Scholar] [CrossRef] [Green Version]
- Kremer, D.; Müller, I.D.; Dunkić, V.; Vitali, D.; Stabentheiner, E.; Oberländer, A.; Bezić, N.; Kosalec, I. Chemical traits and antimicrobial activity of endemic Teucrium arduini L. from Mt Biokovo (Croatia). Cent. Eur. J. Biol. 2012, 7, 941–947. [Google Scholar] [CrossRef] [Green Version]
- Payne, W.W. A glossary of plant hair thermonology. Brittonia 1978, 30, 239–255. [Google Scholar] [CrossRef]
- Fredotović, Ž.; Šprung, M.; Soldo, B.; Ljubenkov, I.; Budić-Leto, I.; Bilušić, T.; Cikeš-Čulić, V.; Puizina, J. Chemical composition and biological activity of Allium cepa L. and Allium × cornutum (Clementi ex Visiani 1842) methanolic extracts. Molecules 2017, 22, 448. [Google Scholar] [CrossRef] [Green Version]
- Mensor, L.L.; Menezes, F.S.; Leitão, G.G.; Reis, A.S.; dos Santos, T.C.; Coube, C.S.; Leitão, S.G. Screening of Brazilian plant extracts for antioxidant activity by the use of DPPH free radical method. Phyther. Res. 2001, 15, 127–130. [Google Scholar] [CrossRef]
- Payet, B.; Sing, A.S.C.; Smadja, J. Assessment of antioxidant activity of cane brown sugars by ABTS and DPPH radical scavenging assays: Determination of their polyphenolic and volatile constituents. J. Agric. Food Chem. 2005, 53, 10074–10079. [Google Scholar] [CrossRef]
- Hazzoumi, Z.; Moustakime, Y.; Joutei, K.A. Essential Oil and Glandular Hairs: Diversity and Roles. In Essential Oils—Oils of Nature; El-Shemy, H.A., Ed.; IntechOpen: London, UK, 2019; pp. 1–16. [Google Scholar]
- Haratym, W.; Weryszko-Chmielewska, E. Ultrastructural and histochemical analysis of glandular trichomes of Marrubium vulgare L. (Lamiaceae). Flora Morphol. Distrib. Funct. Ecol. Plants 2017, 231, 11–20. [Google Scholar] [CrossRef]
- Baran, P.; Aktaş, K.; Özdemir, C. Structural investigation of the glandular trichomes of endemic Salvia smyrnea L. S. Afr. J. Bot. 2010, 76, 572–578. [Google Scholar] [CrossRef] [Green Version]
- Kristen, U.; Lockhausen, J. The leaf glands of Veronica beccabunga L.: Ultrastructure and a possible pathway of secretion. Isr. J. Plant Sci. 2015, 34, 147–156. [Google Scholar] [CrossRef]
- Kurer, G.A. Kutikularfalten und Protuberanzen an Haaren und Epidermen; Druck, Gebr. Leeman: Dresden, Germany, 1917. [Google Scholar]
- Kraehmer, H.; Baur, P. Weed Anatomy; Wiley-Blackwell: Hoboken, NJ, USA, 2013; ISBN 978-0-470-65986-1. [Google Scholar]
- Ruščić, M.; Vuko, E.; Bezić, N.; Kremer, D.; Dunkić, V. Croatian Micromeria fruticulosa-essential oil composition and antiphytoviral activity. Nat. Prod. Commun. 2017, 12, 1133–1135. [Google Scholar] [CrossRef] [Green Version]
- An, M.; Zhou, T.; Guo, Y.; Zhao, X.; Wu, Y. Molecular Regulation of Host Defense Responses Mediated by Biological Anti-TMV Agent Ningnanmycin. Viruses 2019, 11, 815. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Luo, Y.; Qin, S.; Xi, L.; Wan, B.; Du, L. Induction of systemic resistance against tobacco mosaic virus by Ningnanmycin in tobacco. Pestic. Biochem. Physiol. 2014, 111, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Song, B.; Bhadury, P.S.; Jin, L.; Hu, D.; Yang, S. Antiviral activity and mechanism of action of novel thiourea containing chiral phosphonate on tobacco mosaic virus. Int. J. Mol. Sci. 2011, 12, 4522–4535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, D.; Wang, D.; Chen, K.; Huang, M.; Xie, X.; Li, X. Activation of biochemical factors in CMV-infected tobacco by ningnanmycin. Pestic. Biochem. Physiol. 2019, 156, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Tomassini, L.; Brkic, D.; Serafini, M.; Nicoletti, M. Constituents of Veronica hederifolia and Veronica polita. Fitoterapia 1995, 66, 382. [Google Scholar]
- Su, B.N.; Zhu, Q.X.; Jia, Z.J. Aquaticol, a novel bis-sesquiterpene from Veronica anagallis-aquatica. Tetrahedron Lett. 1999, 40, 357–358. [Google Scholar] [CrossRef]
- Graham, J.G.; Quinn, M.L.; Fabricant, D.S.; Farnsworth, N.R. Plants used against cancer—An extension of the work of Jonathan Hartwell. J. Ethnopharmacol. 2000, 73, 347–377. [Google Scholar] [CrossRef]
- Chari, V.M.; Grayer-Barkmeijer, R.J.; Harborne, J.B.; Osterdahl, B.-G. An acylated allose-containing 8-hydroxyflavone glycoside from Veronica filiformis. Phytochemistry 1981, 20, 1977–1979. [Google Scholar] [CrossRef]
- Taskova, R.; Handjieva, N.; Peev, D.; Popov, S. Iridoid glucosides from three Veronica species. Phytochemistry 1998, 49, 1323–1327. [Google Scholar] [CrossRef]
- Harput, U.S.; Saracoglu, I.; Inoue, M.; Ogihara, Y. Anti-inflammatory and cytotoxic activities of five Veronica species. Biol. Pharm. Bull. 2002, 25, 483–486. [Google Scholar] [CrossRef] [Green Version]
- Saracoglu, I.; Harput, U.S.; Inoue, M.; Ogihara, Y. New phenylethanoid glycosides from Veronica pectinata var glandulosa and their free radical scavenging activities. Chem. Pharm. Bull. 2002, 50, 665–668. [Google Scholar] [CrossRef] [Green Version]
- Saracoǧlu, I.; Harput-Hudaverdi, U.Ş.; Nagatsu, A.; Varel, M. Cytotoxic Activity and Structural Aspects of Iridoids Isolated from Veronica thymoides subsp. pseudocinerea. In Proceedings of the Natural Products with Pharmaceutical, Nutraceutical, Cosmetic, and Agrochemical Interest, 7th Joint Meeting of AFERP, ASP, GA, and PSE (49th Annual Meeting of the American Society of Pharmacognosy), Athens, Greece, 8 July 2008; p. 1046. [Google Scholar]
- Saracoǧlu, I.; Harput-Hudaverdi, U.Ş. In vitro Anticancer Activity and Structure-Activity Relationships of Iridoid Glucosides. In Proceedings of the 9th International Symposium on Pharmaceutical Sciences, Ankara, Turkey, 23–26 June 2009. [Google Scholar]
- Moreno-Escobar, J.A.; Bazalda, S.; Villarreal, M.L.; Bonilla-Barbosa, J.R.; Mendoza, S.; Rodríguez-López, V. Cytotoxic and antioxidant activities of selected Lamiales species from Mexico. Pharm. Biol. 2011, 49, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Jiang, R.; Sun, L.W. Studies on the anti-tumor activity in vitro of the flavonoid extract in round leaf speedwell. In Proceedings of the First International Conference on Cellular, Molecular Biology, Biophysics and Bioengineering, Lushan, China, 17–18 September 2010. [Google Scholar]
- Saracoglu, I.; Oztunca, F.H.; Nagatsu, A.; Harput, U.S. Iridoid content and biological activities of Veronica cuneifolia subsp. cuneifolia and V. cymbalaria. Pharm. Biol. 2011, 49, 1150–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohadjerani, M.; Asadollahi, S. Veronica crista-galli Steven and Veronica persica Poir. as anticancer and antioxidant plants in-vitro. Trends Phytochem. Res. 2019, 3, 61–66. [Google Scholar] [CrossRef]
- Assaeed, A.; Elshamy, A.; El Gendy, A.E.N.; Dar, B.; Al-Rowaily, S.; Abd-ElGawad, A. Sesquiterpenes-rich essential oil from above ground parts of Pulicaria somalensis exhibited antioxidant activity and allelopathic effect on weeds. Agronomy 2020, 10, 399. [Google Scholar] [CrossRef] [Green Version]
- Chappell, J.; Coates, R.M. Sesquiterpenes. Compr. Nat. Prod. II Chem. Biol. 2010, 1, 609–641. [Google Scholar] [CrossRef]
- Aazza, S.; Lyoussi, B.; Miguel, M. Antioxidant activity of some Morrocan hydrosols. J. Med. Plants Res. 2011, 5, 6688–6696. [Google Scholar] [CrossRef]
- Harput, U.S.; Saracoglu, I.; Genc, Y. Comparative Bioactivity Studies on Four Veronica species. Fabad J. Pharm. Sci. 2009, 34, 67–72. [Google Scholar]
- Harput, U.Ş.; Genç, Y.; Khan, N.; Saracoglu, I. Radical scavenging effects of different Veronica species. Rec. Nat. Prod. 2011, 5, 100–107. [Google Scholar]
- Živković, J.; Ćebović, T.; Maksimović, Z. In vivo and in vitro antioxidant effects of three Veronica species. Cent. Eur. J. Biol. 2012, 7, 559–568. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Sharifi-Rad, M.; Salehi, B.; Iriti, M.; Roointan, A.; Mnayer, D.; Soltani-Nejad, A.; Afshari, A. In vitro and in vivo assessment of free radical scavenging and antioxidant activities of Veronica persica Poir. Cell. Mol. Biol. 2018, 64, 57–64. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
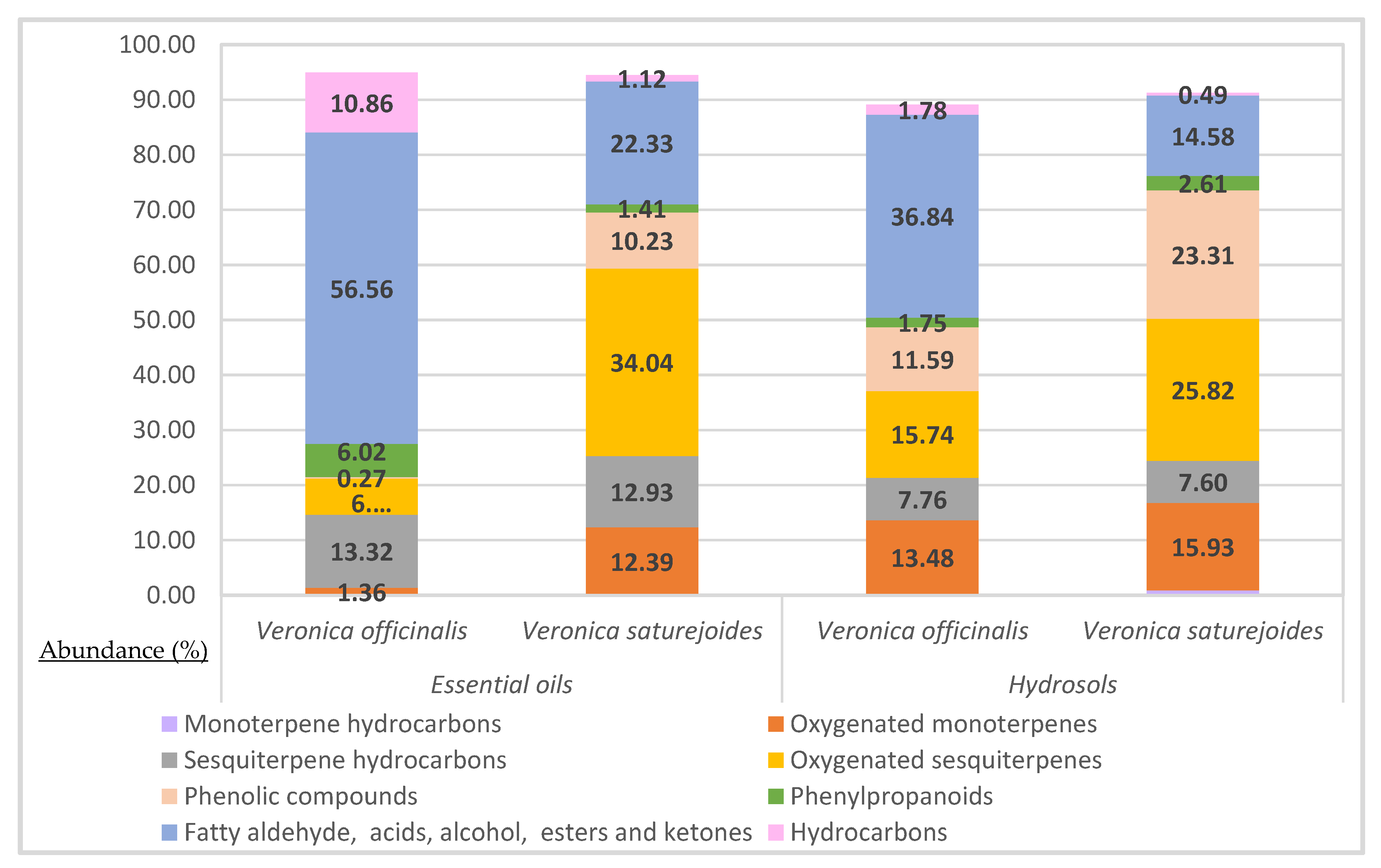
| Locality | Coordinates | Altitude a.s.l. (m) | Date of Collection | |
|---|---|---|---|---|
| Veronica officinalis L. | Kamešnica Mountain | 43°43′03.1″ N 16°50′34.1″ E | 1445 m | July 2021 |
| Veronica saturejoides Vis. ssp. saturejoides | Dinara Mountain | 44°3′2.6″ N; 16°22′52.9″ E | 1650 m | July 2021 |
| V. officinalis V. saturejoides | V. officinalis V. saturejoides | |||||
|---|---|---|---|---|---|---|
| Essential Oils | Hydrosols | |||||
| Component | RI1 | RI2 | Mean ± SD (%) | Mean ± SD (%) | ||
| Monoterpene hydrocarbons | NI | NI | 0.15 | 0.89 | ||
| α-Thujene | 924 | 1012 | NI | NI | NI | 0.23 ± 0.01 |
| α-Pinene * | 935 | 1017 | NI | NI | NI | 0.66 ± 0.03 |
| β -Phellandrene | 1002 | 1195 | NI | NI | 0.15 ± 0.03 | - |
| Oxygenated monoterpenes | 1.36 | 12.39 | 13.48 | 15.93 | ||
| 1,8-Cineole | 1026 | 1210 | NI | NI | 0.76 ± 0.01 | NI |
| γ-Terpinene | 1057 | 1225 | NI | NI | 2.61 ± 0.01 | NI |
| Linalool | 1095 | 1506 | 0.52 ± 0.01 b | 0.89 ± 0.05 a | 4.72 ± 0.01 a | 1.39 ± 0.01 b |
| allo-Ocimene | 1128 | 1390 | 0.22 ± 0.15 | NI | NI | NI |
| Camphor | 1151 | 1499 | NI | NI | 0.72 ± 0.01 | NI |
| Borneol | 1176 | 1719 | NI | NI | 1.59 ± 0.01 | NI |
| α-Terpineol | 1184 | 1660 | NI | 0.88 ± 0.01 | 3.08 ± 0.03 a | 2.79 ± 0.01 b |
| trans-1(7),8-p-Mentadien-2-ol | 1187 | 1803 | 0.62 ± 0.01 | 10.62 ± 0.02 | NI | 11.75 ± 0.01 |
| Sesquiterpene hydrocarbons | 13.32 | 12.93 | 7.76 | 7.6 | ||
| α-Copaene | 1377 | 1484 | 0.78 ± 0.01 | NI | NI | NI |
| E-Caryophyllene * | 1424 | 1585 | 6.78 ± 0.04 b | 7.63 ± 0.01 a | 0.56 ± 0.01 b | 1.39 ± 0.01 a |
| allo-Aromadendrene | 1465 | 1662 | 1.32 ± 0.01 a | 0.87 ± 0.01 b | 2.59 ± 0.01 b | 3.87 ± 0.01 a |
| β-Chamigrene | 1478 | 1724 | NI | NI | 0.27 ± 0.12 | NI |
| Germacrene D | 1481 | 1692 | NI | 2.61 ± 0.01 | NI | 2.34 ± 0.01 |
| δ-Selinene | 1492 | 1756 | 3.32 ± 0.01 | NI | 4.34 ± 0.01 | NI |
| δ-Cadinene | 1517 | 1745 | 1.12 ± 0.01 b | 1.82 ± 0.01 a | NI | NI |
| Oxygenated sesquiterpenes | 6.54 | 34.04 | 15.74 | 25.82 | ||
| Spathulenol | 1577 | 2101 | NI | 1.8 ± 0.01 | 5.25 ± 0.01 | NI |
| Caryophyllene oxide * | 1581 | 1955 | 1.42 ± 0.01 b | 23.65 ± 0.01 a | 7.52 ± 0.01 b | 21.28 ± 0.01 a |
| Viridiflorol | 1592 | 2099 | NI | NI | NI | 1.53 ± 0.01 |
| γ-Eudesmol | 1632 | 2175 | 1.82 ± 0.01 a | 0.2 ± 0.03 b | NI | 0.23 ± 0.01 |
| α-Muurolol | 1645 | 2181 | 3.30 ± 0.01 b | 7.86 ± 0.01 a | 2.38 ± 0.01 | 2.37 ± 0.01 |
| α-Cadinol | 1655 | 2208 | NI | NI | NI | 0.41 ± 0.01 |
| α-Bisabolol | 1685 | 2210 | NI | NI | 0.59 ± 0.03 | NI |
| α-Bisabolol oxide | 1748 | 2511 | NI | 0.53 ± 0.01 | NI | NI |
| Phenolic compounds | 0.27 | 10.23 | 11.59 | 23.31 | ||
| p-Vinyl guaiacol | 1313 | 2156 | 0.27 ± 0.03 | NI | 11.59 ± 0.01 | NI |
| Methyl eugenol | 1403 | 2005 | NI | 10.23 ± 0.01 | NI | 23.31 ± 0.01 |
| Phenylpropanoids | 6.02 | 1.41 | 1.75 | 2.61 | ||
| Z-Methyl isoeugenol | 1451 | 2045 | 1.46 ± 0.03 a | 1.41 ± 0.01 b | 1.75 ± 0.01 b | 2.16 ± 0.03 a |
| Benzyl benzoate | 1760 | 2613 | 4.56 ± 0.01 | NI | NI | NI |
| Fatty aldehyde, acids, alcohol, esters and ketones | 56.56 | 22.33 | 36.84 | 14.58 | ||
| Isopentyl acetate | 863 | 1127 | NI | 6.24 ± 0.01 | 0.24 ± 0.09 b | 0.59 ± 0.01 a |
| Benzaldehyde | 952 | 1508 | 0.98 ± 0.01 b | 4.29 ± 0.01 a | 9.25 ± 0.01 a | 8.87 ± 0.01 b |
| Benzene acetaldehyde | 1036 | 1633 | 0.48 ± 0.01 | NI | 4.75 ± 0.01 a | 3.68 ± 0.01 b |
| n-Nonanal | 1100 | 1389 | 0.89 ± 0.02 | NI | 1.68 ± 0.01 a | 0.28 ± 0.01 b |
| Hexyl 2-methyl butanoate | 1233 | 1425 | NI | NI | 0.21 ± 0.01 a | 0.15 ± 0.03 b |
| Menthyl acetate | 1294 | 1550 | NI | NI | 1.58 ± 0.02 | NI |
| (E)-β-Damascone | 1384 | 1819 | NI | NI | 6.69 ± 0.01 | NI |
| β-Ionone | 1487 | 1935 | 17.88 ± 0.01 | NI | 10.74 ± 0.01 | NI |
| Hexahydrofarnesyl acetone | 1839 | 2113 | 13.92 ± 0.01 a | 6.18 ± 0.01 b | 0.25 ± 0.03 b | 0.39 ± 0.03 a |
| 1-Hexadecanol | 1874 | 2371 | 1.79 ± 0.01 a | 1.51 ± 0.03 b | NI | NI |
| Hexadecanoic acid | 1959 | 2912 | 20.62 ± 0.01 a | 4.11 ± 0.01 b | 1.45 ± 0.01 a | 0.62 ± 0.02 b |
| Hydrocarbons | 10.86 | 1.12 | 1.78 | 0.49 | ||
| Eicosane * | 2000 | 2000 | 4.21 ± 0.01 a | 1.12 ± 0.01 b | 1.51 ± 0.04 | NI |
| Heneicosane * | 2100 | 2100 | 0.98 ± 0.17 | NI | 0.27 ± 0.01 | NI |
| Docosane * | 2200 | 2200 | 2.13 ± 0.01 | NI | NI | NI |
| Tricosane * | 2300 | 2300 | NI | NI | NI | NI |
| Tetracosane * | 2400 | 2400 | 0.83 ± 0.01 | NI | NI | 0.49 ± 0.01 |
| Pentacosane * | 2500 | 2500 | 2.71 ± 0.04 | NI | NI | NI |
| Total identification (%) | 94.93 | 95.45 | 89.09 | 91.23 | ||
| LLN | ||||
|---|---|---|---|---|
| dpi | Mean ± SD | Mean ± SD | ||
| 3rd | C | 5.86 ± 2.39 | C | 6.55 ± 1.24 |
| V. off H | 0.83 ± 0.31 * | V.off EO | 2.57 ± 0.18 * | |
| V. sat H | 1.50 ± 0.62 * | V.sat EO | n.a. | |
| 5th | C | 8.95 ± 2.95 | C | 8.39 ± 2.20 |
| V. off H | 1.36 ± 0.58 * | V.off EO | 2.75 ± 0.58 * | |
| V. sat H | 2.24 ± 0.63 * | V.sat EO | n.a. | |
| 7th | C | 10.55 ± 3.62 | C | 9.67 ± 2.24 |
| V. off H | 2.10 ± 0.87 * | V.off EO | 3.39 ± 0.50 * | |
| V. satH | 3.02 ± 0.89 * | V.sat EO | n.a. | |
| Essential Oils (Mean ± SD) | ||
|---|---|---|
| Antioxidant Assay | V. officinalis | V. saturejoides ssp. saturejoides |
| ORAC (Trolox eq) | 58.75 ± 3.42 b | 263.29 ± 4.89 a |
| DPPH (% inhibition) | 15.51 ± 1.67 | 19.11 ± 4.09 |
| DPPH (IC 50) | 31.34 ± 2.91 b | 15.99 ± 4.17 a |
| Hydrosols (mean ± SD) | ||
| Antioxidant Assay | V. officinalis | V. saturejoides ssp. saturejoides |
| ORAC (Trolox eq) | 0.307 ± 0.011 | 0.258 ± 0.013 |
| DPPH (% inhibition) | 27.74 ± 0.77 b | 38.39 ± 5.83 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazlić, M.; Fredotović, Ž.; Vuko, E.; Fabijanić, L.; Kremer, D.; Stabentheiner, E.; Ruščić, M.; Dunkić, V. Wild Species Veronica officinalis L. and Veronica saturejoides Vis. ssp. saturejoides—Biological Potential of Free Volatiles. Horticulturae 2021, 7, 295. https://doi.org/10.3390/horticulturae7090295
Nazlić M, Fredotović Ž, Vuko E, Fabijanić L, Kremer D, Stabentheiner E, Ruščić M, Dunkić V. Wild Species Veronica officinalis L. and Veronica saturejoides Vis. ssp. saturejoides—Biological Potential of Free Volatiles. Horticulturae. 2021; 7(9):295. https://doi.org/10.3390/horticulturae7090295
Chicago/Turabian StyleNazlić, Marija, Željana Fredotović, Elma Vuko, Lea Fabijanić, Dario Kremer, Edith Stabentheiner, Mirko Ruščić, and Valerija Dunkić. 2021. "Wild Species Veronica officinalis L. and Veronica saturejoides Vis. ssp. saturejoides—Biological Potential of Free Volatiles" Horticulturae 7, no. 9: 295. https://doi.org/10.3390/horticulturae7090295
APA StyleNazlić, M., Fredotović, Ž., Vuko, E., Fabijanić, L., Kremer, D., Stabentheiner, E., Ruščić, M., & Dunkić, V. (2021). Wild Species Veronica officinalis L. and Veronica saturejoides Vis. ssp. saturejoides—Biological Potential of Free Volatiles. Horticulturae, 7(9), 295. https://doi.org/10.3390/horticulturae7090295