
Unraveling the Interaction between Arbuscular Mycorrhizal Fungi and Camellia Plants


 ,
, 
Abstract
:1. Introduction
2. AMF Diversity in Rhizosphere of Camellia spp.
2.1. Morphological Identification
2.2. Molecular Identification
3. AMF Colonization of Camellia Plants and Its Influencing Factors
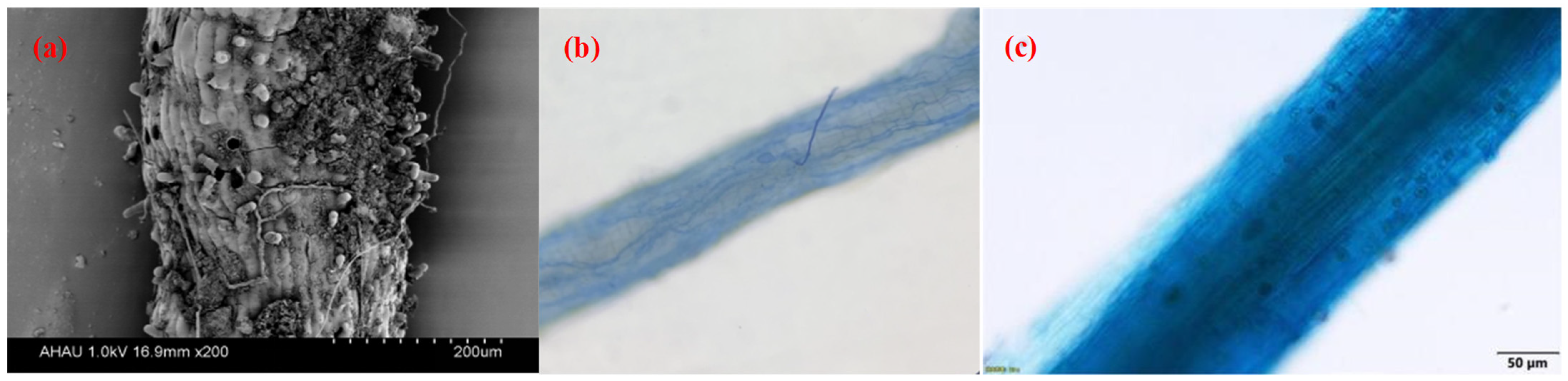
3.1. Root AMF Colonization of Camellia Plants
3.2. Factors Affecting AMF Colonization
3.2.1. Seasonal Variations
3.2.2. Soil Factors
4. AMF Diversity in Rhizosphere of Camellia spp.
4.1. Plant Growth and Development
4.2. Nutrient Uptake
4.3. Stress Resistance
4.4. Food Quality
5. Conclusions
- (1)
- The rhizosphere of Camellia plants in open-field under non-AMF inoculation conditions has an AMF community based on morphological identification. Indigenous AMF colonizes roots of Camellia plants to form a symbiotic association. Due to the limitation of morphological identification, more work around high-throughput sequencing should be performed as much as possible to accurately identify the AMF community and provide a basis for screening of the suitable dominant strains for its application. In addition, future work needs to screen effective AMF strains in promoting plant growth of Camellia under non-sterilized soil conditions and in different soil types.
- (2)
- Among Camellia plants, tea plants are rich in natural tea polyphenols, caffeine and other active ingredients; seeds of C. oleifera can be extracted as oil (tea oil) for consumption; the flower size, number and brightness of C. japonica (an ornamental plant) are important indicators for ornamental purposes. Earlier studies on AMF and Camellia plants focused on plant growth, nutrients and stress resistance. However, few studies have addressed the effects of AMF on functional constituents of tea and the oil yield and composition in the seeds of C. oleifera. The effects of AMF on the ornamental properties of C. japonica are also unknown. Future experiments should focus on the above aspects.
- (3)
- AMF promote the absorption of nutrients (especially P) from the soil of Camellia plants, while the underlying mechanism is unknown. In addition, tea plants are typically grown in extremely acidic soil conditions where aluminum is relatively rich, resulting in aluminum stress in tea plants. Future work needs to revolve around whether and how AMF affects the aluminum tolerance of tea plants.
- (4)
- In-depth study on the mechanism of AMF on enhancing stress tolerance of Camellia plants at physiological and molecular levels.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, L.; Zou, Y.N.; Tian, Z.H.; Wu, Q.S.; Kuča, K. Effects of beneficial endophytic fungal inoculants on plant growth and nutrient absorption of trifoliate orange seedlings. Sci. Hortic. 2021, 277, 109815. [Google Scholar] [CrossRef]
- Meng, L.L.; He, J.D.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Mycorrhiza-released glomalin-related soil protein fractions contribute to soil total nitrogen in trifoliate orange. Plant Soil Environ. 2020, 66, 183–189. [Google Scholar] [CrossRef]
- Wu, Q.S.; Gao, W.Q.; Srivastava, A.K.; Zhang, F.; Zou, Y.N. Nutrient acquisition and fruit quality of Ponkan mandarin in response to AMF inoculation. Ind. J. Agric. Sci. 2020, 90, 1563–1567. [Google Scholar]
- Zou, Y.N.; Zhang, F.; Srivastava, A.K.; Wu, Q.S.; Kuča, K. Arbuscular mycorrhizal fungi regulate polyamine homeostasis in roots of trifoliate orange for improved adaptation to soil moisture deficit stress. Front. Plant Sci. 2021, 11, 600792. [Google Scholar] [CrossRef]
- Eid, K.E.; Abbas, M.H.H.; Mekawi, E.M.; EINagar, M.M.; Abdelhafez, A.A.; Amin, B.H.; Mohamed, I.; Ali, M.B. Arbuscular mycorrhiza and environmentally biochemicals enhance the nutritional status of Helianthus tuberosus and induce its resistance against Sclerotium rolfsii. Ecotox. Environ. Saf. 2019, 186, 109783. [Google Scholar] [CrossRef]
- Karthikeyan, A.; Muthukumar, T.; Udaiyan, K. Response of tea (Camellia sinensis (L). Kuntze) to arbuscular mycorrhizal fungi under plantation nursery conditions. Biol. Agric. Hortic. 2005, 22, 305–319. [Google Scholar] [CrossRef]
- Zhang, F.; Zou, Y.N.; Wu, Q.S. Quantitative estimation of water uptake by mycorrhizal extraradical hyphae in citrus under drought stress. Sci. Hortic. 2018, 229, 132–136. [Google Scholar] [CrossRef]
- Wu, Q.S.; Srivastava, A.K.; Zou, Y.N. AMF-induced tolerant to drought stress in citrus: A review. Sci. Hortic. 2013, 164, 77–87. [Google Scholar] [CrossRef]
- González-Chávez, M.C.; Carrillo-González, R.; Wright, S.F.; Nichols, K.A. The role of glomalin, a protein produced by arbuscular mycorrhizal fungi, in sequestering potentially toxic elements. Environ. Pollut. 2004, 130, 317–323. [Google Scholar] [CrossRef]
- Cornejo, P.; Meier, S.; Borie, G.; Rillig, M.C.; Borie, F. Glomalin related soil protein in a Mediterranean ecosystem affected by a copper smelter and its contribution to Cu and Zn sequestration. Sci. Total Environ. 2008, 406, 154–160. [Google Scholar] [CrossRef]
- He, J.D.; Chi, G.G.; Zou, Y.N.; Shu, B.; Wu, Q.S.; Srivastava, A.K.; Kuča, K. Contribution of glomalin-related soil proteins to soil organic carbon in trifoliate orange. Appl. Soil Ecol. 2020, 154, 103592. [Google Scholar] [CrossRef]
- Cheng, H.Q.; Giri, B.; Wu, Q.S.; Zou, Y.N.; Kuča, K. Arbuscular mycorrhizal fungi mitigate drought stress in citrus by modulating root microenvironment. Arch. Agron. Soil Sci. 2021. [Google Scholar] [CrossRef]
- Luo, C.Q.; Tan, X.F.; Ling, L.L. A Classification summary on plant of genus Camellia. J. Cent. South For. Univ. 1999, 19, 78–81. [Google Scholar]
- Mondal, T.K.; Bhattacharya, A.; Laxikumaran, M.; Ahuja, P.S. Recent advance of tea (Camellia sinensis) biotechnology. Plant Cell Tiss. Org. 2004, 76, 195–254. [Google Scholar] [CrossRef]
- Singh, S.; Pandey, A.; Kumar, B.; Palni, L.M.S. Enhancement in growth and quality parameters of tea [Camellia sinensis (L.) O. Kuntze] through inoculation with arbuscular mycorrhizal fungi in an acid soil. Biol. Fert. Soils 2010, 46, 427–433. [Google Scholar] [CrossRef]
- Lin, X.G.; Hao, W.Y. Mycorrhizal dependency of various kind of plants. Acta Bot. Sin. 1989, 31, 721–725. [Google Scholar]
- Gao, X.B.; Chen, J.; Zhao, J.F.; Li, Z.; Guo, C.; Zhou, F.Y.; Wang, Z.X.; Tu, Y.Y.; Zhou, Y.F. Colonization characteristics of arbuscular mycorrhiza fungi in rhizosphere of local tea trees in Guizhou. Southwest China J. Agric. Sci. 2016, 29, 1328–1335. [Google Scholar]
- Shao, Y.D.; Zhang, D.J.; Hu, X.C.; Wu, Q.S.; Jiang, C.J.; Xia, T.J.; Gao, X.B.; Kuča, K. Mycorrhiza-induced changes in root growth and nutrient absorption of tea plants. Plant Soil Environ. 2018, 64, 283–289. [Google Scholar]
- Lee, C.P.; Shih, P.H.; Hsu, C.L.; Yen, G.C. Hepatoprotection of tea seed oil (camellia oleifera abel.) against CCl4-induced oxidative damage in rats. Food Chem. Toxicol. 2007, 45, 888–895. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Meng, J.X.; Pan, D.F.; Yang, C.; Li, Y. Mating system and progeny genetic diversity of camellia oleifera ‘Ruan Zhi’. J. For. Res. 2019, 30, 267–272. [Google Scholar] [CrossRef]
- Liu, R.C.; Xiao, Z.Y.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.S. Mycorrhizal fungal diversity and its relationship with soil properties in Camellia oleifera. Agriculture 2021, 11, 470. [Google Scholar] [CrossRef]
- Hou, H.Y.; Duan, Y.P. Camellia japonica is in landscape afforestation medium disposition and application. Anhui Agri. Sci. Bull. 2009, 15, 76. [Google Scholar]
- Berruti, A.; Demasi, S.; Lumini, E.; Kobayashi, N.; Bianciotto, V.; Bianciotto, V. Wild Camellia japonica specimens in the Shimane prefecture (Japan) host previously undescribed AMF diversity. Appl. Soil Ecol. 2017, 115, 10–18. [Google Scholar] [CrossRef]
- Tunstall, A.C. Mycorrhiza in tea plants. Quart. J. Indian Tea Assoc. Indian 1926, 159. [Google Scholar]
- Singh, S.; Pandey, A.; Chaurasia, B.; Palni, L.M.S. Diversity of arbuscular mycorrhizal fungi associated with the rhizosphere of tea growing in ‘natural’ and ‘cultivated’ ecosites. Biol. Fert. Soils 2008, 44, 491–500. [Google Scholar] [CrossRef]
- Gupta, R.K.; Sharma, C. Diversity of arbuscular mycorrhizal fungi in Camellia sinensis in Uttarakhand State, India. Afr. J. Biotechnol. 2013, 9, 5313–5319. [Google Scholar]
- Sharma, C.; Gupta, R.K.; Pathak, R.K.; Choudhary, K.K. Seasonal colonization of arbuscular mycorrhiza fungi in the roots of Camellia sinensis (tea) in different tea gardens of India. ISRN Biodivers. 2015, 2013, 593086. [Google Scholar] [CrossRef]
- Lu, D.S.; Wu, X.Q. Species of VAM fungi around tea roots in the southern area of Henan province. J. Nanjing For. Univ. 2005, 29, 33–36. [Google Scholar]
- Wu, L.S.; Wang, Y.; Li, M.; Liu, R.J.; Ding, Z.T. A survey of arbuscular mycorrhizal fungi in the rhizosphere of Camellia sinensis in Laoshan. J. Qingdao Agric. Univ. 2009, 26, 171–173. [Google Scholar]
- Xing, D.; Zhang, A.M.; Li, Z.; Chen, J.; Wang, Z.X.; Tu, Y.Y.; Gao, X.B. Resources and morphological characteristics of arbuscular mycorrhiza fungi around tea rhizosphere in Guizhou. Guizhou Agric. Sci. 2015, 43, 102–106. [Google Scholar]
- Deng, X.J.; Zhou, G.Y.; Liu, J.A.; Li, L.; Bu, T.T. Diversity and community structure of arbuscular mycorrhizal fungi in Camellia oleifera stands in Hunan. J. Cent. South Univ. For. Tech. 2011, 31, 38–42. [Google Scholar]
- Lin, Y.L.; Li, Z.Y.; Wu, F.; Pei, Y.; Zhang, Y.; Zhang, L.P.; Yang, T.; Tan, M.X. Community structure characteristics of arbuscular mycorrhizal fungi among Camellia oleifera cultivars. For. Res. 2020, 33, 163–169. [Google Scholar]
- Zhou, G.R.; Shang, K.; Jiang, L. Diversity survey of AM fungi in rhizosphere soil of wild Camellia oleifera. J. Guizhou Univ. 2019, 36, 26–31. [Google Scholar]
- Yuan, T.; Tao, G.Y.; Jiang, L. Arbuscular mycorrhizal fungi in the rhizospheric soil of four forest types in Fanjingshan national nature reserve. J. Northeast For. Univ. 2018, 46, 83–86. [Google Scholar]
- He, W. AM Fungi Diversity in the Main Ornamental Gardens of Chongqing Nanshan Botanical Park. Master’s Thesis, Southwest University, Chongqing, China, 2009. [Google Scholar]
- Lee, E.H.; Ka, K.H.; Eom, A.H. Diversity of arbuscular mycorrhizal fungi in rhizospheres of Camellia japonica and neighboring plants inhabiting Wando of Korea. Korean J. Mycol. 2014, 42, 34–39. [Google Scholar] [CrossRef]
- Wu, Q.S.; Srivastava, A.K. AMF diversity in citrus rhizosphere. Ind. J. Agric. Sci. 2017, 87, 653–659. [Google Scholar]
- Gai, J.P.; Feng, G.; Li, X.L. Review of researches on biodiversity of arbusculay mycorrhizal fungi. Soils 2005, 37, 236–242. [Google Scholar]
- Yang, C.X.; Li, L.L. Research progress in arbuscular mycorrhizal fungi identification method application. Guizhou Agric. Sci. 2014, 42, 93–97. [Google Scholar]
- Guo, X.H.; Gong, J. Differential effects of abiotic factors and host plant traits on diversity and community composition of root-colonizing arbuscular mycorrhizal fungi in a salt-stressed ecosystem. Mycorrhiza 2014, 24, 79–94. [Google Scholar] [CrossRef]
- Yang, F.; Cao, J.M.; Chen, Y.Y.; Wang, J.L. Research progress on structure and identification method of arbusular mycorrhizal fungi. Mod. Agric. Sci. Tech. 2019, 17, 152–154+157. [Google Scholar]
- Smith, S.E.; Smith, F.A. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, A.; Konishi, S. Relationship between vesicular-arbuscular mycorrhizal infection and soil phosphorus concentration in tea fields. Soil Sci. Plant Nutr. 1989, 35, 139–143. [Google Scholar] [CrossRef] [Green Version]
- Ren, M.X.; Luo, Y.P. Advances in the study of VA mycorrhizae of tea trees. J. Tea 2005, 31, 28–31. [Google Scholar]
- Mejstrik, V. The frequency of vesicular-arbuscular mycorrhizae in the roots of Camellia japonica L. from different sites in New Zealand. Pac. Sci. 1974, 28, 73–77. [Google Scholar]
- Borriello, R.; Berruti, A.; Lumini, E.; Beffa, M.T.D.; Scariot, V.; Bianciotto, V. Edaphic factors trigger diverse AM fungal communities associated to exotic camellias in closely located Lake Maggiore (Italy) sites. Mycorrhiza 2015, 25, 253–265. [Google Scholar] [CrossRef]
- Bencherif, K.; Boutekrabt, A.; Dalpé, Y.; Sahraoui, A.L.H. Soil and seasons affect arbuscular mycorrhizal fungi associated with Tamarix rhizosphere in arid and semi-arid steppes. Appl. Soil Ecol. 2016, 107, 182–190. [Google Scholar] [CrossRef]
- Varela-Cervero, S.; López-García, A.; Barea, J.M.; Azcón-Aguilar, C. Spring to autumn changes in the arbuscular mycorrhizal fungal community composition in the different propagule types associated to a Mediterranean shrubland. Plant Soil 2016, 408, 1–14. [Google Scholar] [CrossRef]
- Chandra, K.K.; Jamaluddin, A. Seasonal variation of VAM fungi in tree species planted in coalmine overbunden of Kusmunda (MP). J. Trop. For. 1998, 14, 118–123. [Google Scholar]
- Gould, A.B.; Hendrix, J.W.; Ferriss, R.S. Relationship of mycorrhizal activity to time following reclamation of surface mine land in western Kentucky. I. Propagule and spore population densities. Can. J. Bot. 1996, 74, 247–261. [Google Scholar] [CrossRef]
- Liang, S.M.; Zheng, F.L.; Abd_Allah, E.F.; Muthuramalingam, P.; Wu, Q.S.; Hashem, A. Spatial changes of arbuscular mycorrhizal fungi in peach and their correlation with soil properties. Soudi J. Biol. Sci. 2021. [Google Scholar] [CrossRef]
- Lin, Z. Effect of VA mycorrhizal species on tea tree growth and absorption of mineral elements. J. Tea Sci. 1993, 13, 15–20. [Google Scholar]
- Pandey, A.; Palni, L.M.S. Bacillus species: The dominant bacteria of the rhizosphere of established tea bushes. Microbiol. Res. 1997, 152, 359–365. [Google Scholar] [CrossRef]
- Avio, L.; Castaldini, M.; Fabiani, A.; Bedini, S.; Sbrana, C.; Turrini, A.; Giovannetti, M. Impact of nitrogen fertilization and soil tillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem. Soil Biol. Biochem. 2013, 67, 285–294. [Google Scholar] [CrossRef]
- He, S.L.; Liu, B.Y. Effect of VA mycorrhizal bacteria on tea tree mineral nutrition and its mechanism. J. Southwest Agric. Univ. 1994, 16, 492–496. [Google Scholar]
- Shao, Y.D.; Hu, X.C.; Wu, Q.S.; Yang, T.Y.; Srivastava, A.K.; Zhang, D.J.; Gao, X.B.; Kuča, K. Mycorrhizas promote P acquisition of tea plants through changes in root morphology and P transporter gene expression. S. Afr. J. Bot. 2021, 137, 455–462. [Google Scholar] [CrossRef]
- Chen, W.L.; Zhao, X.G.; Wang, H.; Yuan, Z.L. Effects of arbuscular mycorrhizal fungi (AMF) inoculation on detoxification capacity of tea tree under Pb stress. Henan Sci. 2014, 32, 511–515. [Google Scholar]
- Liu, J.; Xiao, B.; Wang, L.X.; Li, J.; Pu, G.T.; Gao, T.; Liu, W. Influence of AM on the growth of tea plant and tea quality under salt stress. J. Tea Sci. 2013, 33, 140–146. [Google Scholar]
- Liu, C.Y.; Wang, Y.J.; Wu, Q.S.; Yang, T.Y.; Kuča, K. Arbuscular mycorrhizal fungi improve the antioxidant capacity of tea (Camellia sinensis) seedlings under drought stress. Not. Bot. Horti Agrobot. 2020, 48, 1993–2005. [Google Scholar] [CrossRef]
- Xu, P.H.; Wang, F.Q.; Qi, Y.G.; Zhang, F.; Yang, Q.; Xiao, B. Effect of arbuscular mycorrhizal fungi on drought resistance in tea plant (Camellia sinensis). Acta Agric. Bor. Occid. Sin. 2017, 26, 1033–1040. [Google Scholar]
- Lin, Y.L.; Li, Z.Y.; Zhang, L.P.; Wu, F.; Yang, Y.; Tan, M.X.; Hu, D. Effects of organic phosphorus and AM fungi on growth, root morphology and photosynthetic characteristics of Camellia oleifera. Non-Wood For. Res. 2021, 39, 121–128. [Google Scholar]
- Wang, D.X.; Chen, G.C.; Jiang, Z.P. Effects of arbuscular mycorrhiza on the growth and absorption of nitrogen and phosphorus in Camellia oleifera seedlings. Chin. For. Sci. Technol. 2010, 24, 23–26. [Google Scholar]
- Wang, D.X.; Zhang, N.Y.; Chen, G.C. Effects of AM fungi on the growth and drought-resistance of Camellia oleifera. Guangxi For. Sci. 2011, 40, 259–261. [Google Scholar]
- Berruti, A.; Borriello, R.; Beffa, M.; Scariot, V.; Bianciotto, V. Application of nonspecific commercial AMF inocula results in poor mycorrhization in Camellia japonica L. Symbiosis 2013, 61, 63–76. [Google Scholar] [CrossRef]
- Selvakumar, G.; Shagol, C.C.; Kim, K.; Han, S.; Tongmin, S. Spore associated bacteria regulates maize root K+/Na+ ion homeostasis to promote salinity tolerance during arbuscular mycorrhizal symbiosis. BMC Plant Biol. 2018, 18, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.N.; Wu, Q.S.; Li, Y.; Huang, Y.M. Effects of arbuscular mycorrhizal fungi on root system morphology and sucrose and glucose contents of Poncirus trifoliata. Chin. J. Appl. Ecol. 2014, 25, 1125–1129. [Google Scholar]
- Shao, Y.D.; Zhang, D.J.; Hu, X.C.; Wu, Q.S.; Jiang, C.J.; Gao, X.B.; Kuča, K. Arbuscular mycorrhiza improves leaf food quality of tea plants. Not. Bot. Horti Agrobo. 2019, 47, 608–614. [Google Scholar] [CrossRef] [Green Version]
- Shu, J.L.; Li, M.J. Physiological effect of VA mycorrhizal study in tea tree. J. Tea Sci. 1987, 7, 7–14. [Google Scholar]
- Wu, F.; Li, Z.; Lin, Y.; Zhang, L. Effects of Funneliformis mosseae on the utilization of organic phosphorus in Camellia oleifera Abel. Can. J. Microbiol. 2021, 67, 349–357. [Google Scholar] [CrossRef]
- Huang, Y.F.; Wu, Q.L.; Wan, Q.; Shu, B. Research progress of arbuscular mycorrhizal fungi. Mod. Agric. 2019, 12, 9–12. [Google Scholar]
- Zhang, F.; Wang, P.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Effects of mycorrhizal fungi on root-hair growth and hormone levels of taproot and lateral roots in trifoliate orange under drought stress. Arch. Agron. Soil Sci. 2019, 65, 1316–1330. [Google Scholar] [CrossRef]
- Cheng, H.Q.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Arbuscular mycorrhizal fungi alleviate drought stress in trifoliate orange by regulating H+-ATPase activity and gene expression. Front. Plant Sci. 2021, 12, 659694. [Google Scholar] [CrossRef]
- Cheng, X.F.; Wu, H.H.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Mycorrhizal response strategies of trifoliate orange under well-watered, salt stress, and waterlogging stress by regulating leaf aquaporin expression. Plant Physiol. Biochem. 2021, 162, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.N.; Wu, Q.S.; Kuča, K. Unravelling the role of arbuscular mycorrhizal fungi in mitigating the oxidative burst of plants under drought stress. Plant Biol. 2021, 23, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Z. Effects of arbuscular mycorrhizal fungi on growth and quality of tea under adversity stress. J. Green Sci. Technol. 2016, 18, 127–128. [Google Scholar]
- Zhao, Q.H.; Sun, L.T.; Wang, Y.; Ding, Z.T.; Li, M. Effects of arbuscular mycorrhizal fungi and nitrogen regimes on plant growth, nutrient uptake and tea quality in Camellia sinensis (L.) O. Kuntze. Plant Physiol. J. 2014, 50, 164–170. [Google Scholar]
- Wang, Y.J.; Gao, X.B.; Wu, Q.S.; Ji, D.B.; Cai, F.; Liu, C.Y. Influences of arbuscular mycorrhizal fungi on plant growth and tea quality of Fuding Dabaicha seedlings under different water conditions. J. Tea Sci. 2020, 40, 588–596. [Google Scholar]
- Cao, J.L.; Shao, Y.D.; Zou, Y.N.; Wu, Q.S.; Yang, T.Y.; Kuča, K. Inoculation with Clariodeoglomus etunicatum improves leaf food quality of tea exposed to P stress. Not. Bot. Horti Agrobo. 2021, 49, 12166. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Camellia Plants | Sampling Regions | Identification Method | Identified Genena of AMF | Dominant Genus of AMF | Reference |
|---|---|---|---|---|---|
| C. sinensis | Uttaranchal Himalaya (India) | Morphology | Acaulospora; Gigaspora; Glomus; Scutellospora | Glomus | [25] |
| Dehradun District (India) | Morphology | Glomus | Glomus | [26] | |
| Dehradun Himalaya (India) | Morphology | Acaulospora; Glomus; Gigaspora | Acaulospora and Glomus | [27] | |
| Henan (China) | Morphology | Acaulospora; Gigaspora; Glomus; Scutellospora | Acaulospora and Glomus | [28] | |
| Qingdao (China) | Morphology | Acaulospra; Gigaspora; Glomus | Acaulospra and Glomus | [29] | |
| Guizhou (China) | Morphology | Acaulospora; Entrophospora; Gigaspora; Glomus | Acaulospra and Glomus | [30] | |
| C. oleifera | Hunan (China) | Morphology | Acaulospora; Glomus; Scutellospora | Glomus | [31] |
| Wuhan (China) | High-throughput sequencing of 18S rRNA gene | Acaulospora; Ambispora; Archaeospora; Claroideoglomus; Diversispora; Gigaspora; Glomus; Paraglomus; Redeckera; Scutellospora | Glomus | [21] | |
| Jiangxi (China) | High-throughput sequencing of 18S rRNA gene | Acaulospora; Ambispora; Archaeospora; Claroideoglomus; Diversispora; Geosiphon; Gigaspora; Glomus; Pacispora; Paraglomus; Scutellospora; Septoglomus | Glomus | [32] | |
| Guiyang (China) | High-throughput sequencing of 18S rRNA gene | Acaulospora; Archaeospora; Claroideoglomus; Diversispora; Glomus; Paraglomus | Glomus | [33] | |
| C. japonica | Fanjing Mountain (China) | Morphology | Acaulospora; Funneliformis; Glomus; Pacispora; Scutellospora; | Glomus | [34] |
| Chongqing (China) | Morphology | Acaulospora; Gigaspora; Glomus; Scutellospora | Acaulospora and Glomus | [35] | |
| Shimane prefecture (Japan) | High-throughput sequencing of 18S rRNA gene | Acaulospora; Ambispora; Archaeospora; Claroideoglomus; Diversispora; Funneliformis; Geosiphon; Gigaspora; Glomus; Paraglomus; Redeckera; Rhizophagus; Scutellospora | Glomus and Rhizophagus | [23] | |
| Diankwan Island (Korea) | High-throughput sequencing of 18S rRNA gene | Acaulospora; Ambispora; Claroideoglomus; Glomus; Rhizophagus; Scutellospora | Acaulospora and Rhizophagus | [36] |
| Camellia Plants | Mycorrhizal Fungi | Mycorrhizal Effects on Camellia Plants | Reference |
|---|---|---|---|
| Camellia sinensis | Acauospora scrobiculata, Glomus aggregatum, G. fasciculatum, G. geosporum, G. intraradices, and Scutellospora calospora | biomass ↑ | [6] |
| Ac. spinosa, Ac. sp. 1, G. aggregatum, G. ambisporum, G. clavisporum, G. geosporum, G. mosseae, G. pustulatum, and Glomus sp. | leaf sugar ↑; amino acids ↑; proteins ↑ | [15] | |
| Claroideoglomus etunicatum, Diversispora spurca, D. versiformis, and mixed-AMF | stem diameter ↑; plant height ↑; leaf area ↑; bud number ↑; root morphogenesis ↑; root-hair growth ↑ | [18] | |
| G. epigaeumg | biomass ↑; mineral elements ↑; leaf P, Mg, Fe, Zn, and Cu ↑; chlorophyll ↑; soil phosphatase activity ↑ | [55] | |
| G. versiforme under salt stress | growth ↑; leaf and root N, P, K, Mg, Fe, and Zn ↑; water saturation deficit↓ | [56] | |
| Clariodeoglomus etunicatum under drought stress | leaf water content ↑; antioxidant enzyme activity ↑ | [57] | |
| G. intraradices, G. mosseae, and G. versiforme under drought stress | plant growth performance ↑; soluble protein ↑; proline ↑; malondialdehyde↓; superoxidase ↑; peroxidase ↑; catalase ↑; glutathione ↑ | [58] | |
| C. etunicatum under P stress | root system architecture ↑; root P ↑; root acid phosphatase in P50 ↑; soil neutral and total phosphatase ↑; CsPT1 ↑; CsPT4↓ | [59] | |
| G. intraradices under Pb stress | Biomass ↑; glomalin ↑; Pb in glomalin ↑ | [60] | |
| Camellia oleifera | Funneliformis mosseae | total leaf area ↑; root length ↑; root average diameter ↑ | [61] |
| G. versiforme and G. mosseae | root biomass ↑; root P ↑; leaf N↓ | [62] | |
| G.intraradices, G. mosseae, and G. versiforme | leaf water content ↑; stability of cell membrane ↑; soluble sugar ↑ | [63] | |
| Camellia japonica | F. mosseae | number of flowers ↑; flower depth ↑; leaf size ↑; chlorophyll ↑; root Ca, Mg, K, Cu, Mn, Fe, and Zn ↑; leaf Cu and Mn ↑ | [64] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, R.-C.; Xiao, Z.-Y.; Hashem, A.; Abd_Allah, E.F.; Xu, Y.-J.; Wu, Q.-S. Unraveling the Interaction between Arbuscular Mycorrhizal Fungi and Camellia Plants. Horticulturae 2021, 7, 322. https://doi.org/10.3390/horticulturae7090322
Liu R-C, Xiao Z-Y, Hashem A, Abd_Allah EF, Xu Y-J, Wu Q-S. Unraveling the Interaction between Arbuscular Mycorrhizal Fungi and Camellia Plants. Horticulturae. 2021; 7(9):322. https://doi.org/10.3390/horticulturae7090322
Chicago/Turabian StyleLiu, Rui-Cheng, Zhi-Yan Xiao, Abeer Hashem, Elsayed Fathi Abd_Allah, Yong-Jie Xu, and Qiang-Sheng Wu. 2021. "Unraveling the Interaction between Arbuscular Mycorrhizal Fungi and Camellia Plants" Horticulturae 7, no. 9: 322. https://doi.org/10.3390/horticulturae7090322
APA StyleLiu, R. -C., Xiao, Z. -Y., Hashem, A., Abd_Allah, E. F., Xu, Y. -J., & Wu, Q. -S. (2021). Unraveling the Interaction between Arbuscular Mycorrhizal Fungi and Camellia Plants. Horticulturae, 7(9), 322. https://doi.org/10.3390/horticulturae7090322