
Generation of Composite Rosa roxburghii Plants with Transgenic Roots by Agrobacterium-Mediated Transformation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Agrobacterium Strains
2.2. Agrobacterium-Mediated Root Transformation and Generation of Composite R. roxburghii Tratt. Plants
2.3. Fluorescence Observation
2.4. DNA, RNA Extraction and PCR, RT-PCR Analysis
2.5. Statistical Analysis
3. Results
3.1. Establishment of Two Agrobacterium-Mediated Transformation Methods for Generating Composite R. roxburghii Tratt. Plants
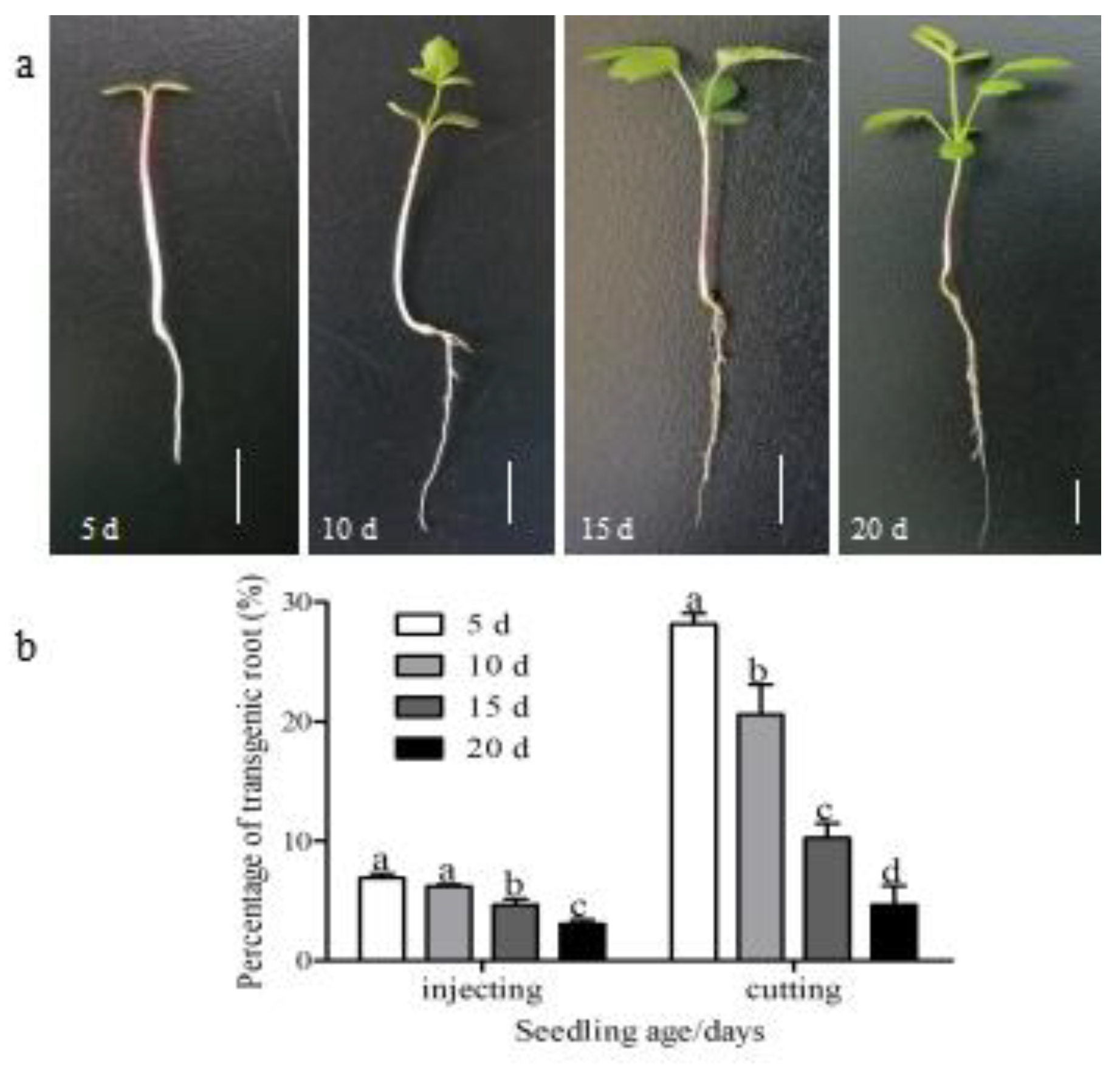
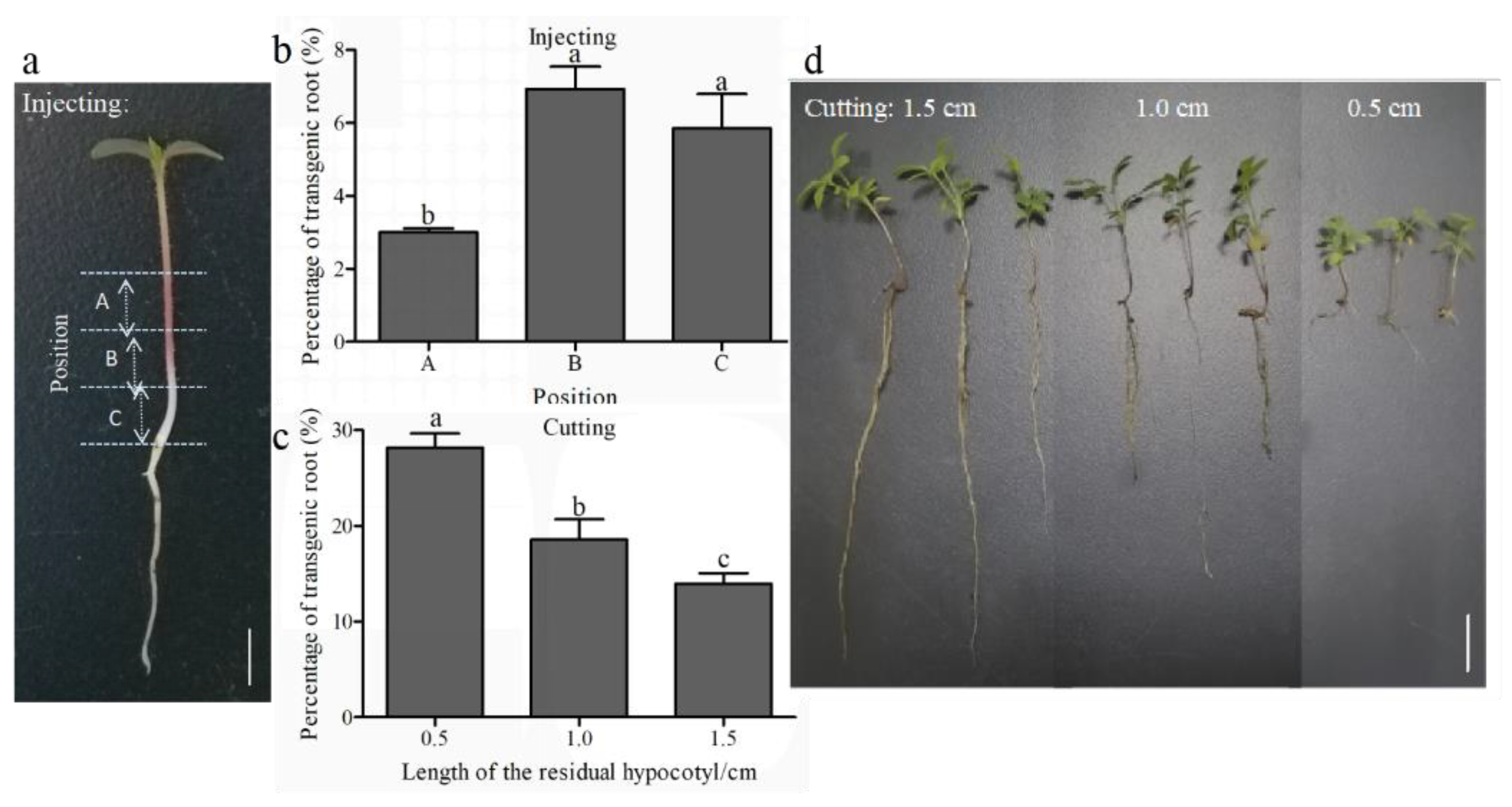
3.2. Optimization of Bacterial Concentration, Seedling Age, Infection Site, and Infection Methods
4. Discussion
4.1. Establishment of Two Agrobacterium-Mediated Transformation Methods for Generating Composite R. roxburghii Plants
4.2. Optimization of Agrobacterium-Mediated Transformation System
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lu, M.; Zhang, H.S.; An, H.M. Chloroplast DNA-based genetic variation of Rosa roxburghii in Southwest China: Phylogeography and conservation Implications. Hortic. Plant J. 2021, 7, 286–294. [Google Scholar] [CrossRef]
- Huang, M.; Xu, Q.; Deng, X.X. l-Ascorbic acid metabolism during fruit development in an ascorbate-rich fruit crop chestnut rose (Rosa roxburghii Tratt.). J. Plant Physiol. 2014, 171, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.C.; Yao, Q.P.; Zhu, D.Q.; Dong, B.Y. Hydrogen-rich water maintains the quality of Rosa roxburghii by regulating AsA biosynthesis and regeneration. Postharvest Biol. Technol. 2022, 195, 112136. [Google Scholar] [CrossRef]
- Huang, X.L.; Yan, H.Q.; Zhai, L.S.; Yang, Z.T.; Yi, Y. Characterization of the Rosa roxburghii Tratt. transcriptome and analysis of MYB genes. PLoS ONE 2019, 14, e0203014. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Ma, W.T.; Liu, Y.Q.; An, H.M.; Ludlow, R.A. Transcriptome analysis reveals candidate lignin-related genes and transcription factors in Rosa roxburghii during fruit ripening. Plant Mol. Biol. Report. 2020, 38, 331–342. [Google Scholar] [CrossRef]
- Yan, H.Q.; Wu, Z.M.; Liu, Y.J.; Weng, Q.B.; Yi, Y.; Huang, X.L. Functional divergence of RrGL3 and RrEGL3 from Rosa roxburghii in mediating trichome development. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 147, 313–324. [Google Scholar] [CrossRef]
- Lu, C. Establishment of VIGS System in Rosa roxburghii Tratt. and the Effect of RrGGP Silencing on Fruit AsA Biosynthesis. Master’s Dissertation, Guizhou University, Guiyang, China, 2021. [Google Scholar]
- Liu, S.; Wang, X.Y.; Li, Q.Q.; Peng, W.T.; Zhang, Z.M.; Chu, P.F.; Guo, S.J.; Fan, Y.L.; Lyu, S.H. AtGCS promoter-driven clustered regularly interspaced short palindromic repeats/Cas9 highly efficiently generates homozygous/biallelic mutations in the transformed roots by Agrobacterium rhizogenes–mediated transformation. Front. Plant Sci. 2022, 13, 952428. [Google Scholar] [CrossRef] [PubMed]
- Niazian, M.; Noori, S.A.S.; Galuszka, P.; Mortazavian, S.M.M. Tissue culture-based Agrobacterium-mediated and in planta transformation methods. Soil Water Res. 2017, 53, 133–143. [Google Scholar] [CrossRef] [Green Version]
- Alagarsamy, K.; Shamala, L.F.; Wei, S. Protocol: High efficiency in-planta Agrobacterium-mediated transgenic hairy root induction of Camellia sinensis var. sinensis. Plant Methods 2018, 14, 17. [Google Scholar] [CrossRef]
- Stanišić, M.; Ćosić, T.; Savić, J.; Krstić-Milošević, D.; Mišić, D.; Ninković, S.; Banjac, N. Hairy root culture as a valuable tool for allelopathic studies in apple. Tree Physiol. 2019, 39, 888–905. [Google Scholar] [CrossRef]
- Meng, D.; Yang, Q.; Dong, B.Y.; Song, Z.H.; Niu, L.L.; Wang, L.T.; Cao, H.Y.; Li, H.H.; Fu, Y.J. Development of an efficient root transgenic system for pigeon pea and its application to other important economically plants. Plant Biotechnol. J. 2019, 17, 1804–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.Y.; Song, Z.H.; Du, T.T.; Yang, W.L.; Chen, T.; Han, Y.; Dong, B.Y.; Cao, H.Y.; Niu, L.L.; Amin, R.; et al. CcCIPK14 gene function analysis to illuminate the efficient root transgenic system. J. Vis. Exp. JoVE 2021, 175, e62304. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.L.; Xu, F.L.; Zhou, H.Z.; Liu, X.X.; Yang, X.Y.; Weng, K.X.; Sun, X.L.; Lyu, S.H. A fast, simple, high efficient and one-step generation of composite cucumber plants with transgenic roots by Agrobacterium rhizogenes-mediated transformation. Plant Cell Tissue Organ Cult. (PCTOC) 2020, 141, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.L.; Zhang, X.H.; Zhong, L.J.; Wang, X.Y.; Jin, L.S.; Lyu, S.H. One-step generation of composite soybean plants with transgenic roots by Agrobacterium rhizogenes-mediated transformation. BMC Plant Biol. 2020, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Z.; Li, W.; Xian, W.Y.; Huang, W.; Yang, R.L. Free and bound phenolic profiles of Rosa roxburghii Tratt. leaves and their antioxidant and inhibitory effects on α-Glucosidase. Front. Nutr. 2022, 9, 1379. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.T.; Huang, Q.Y.; Wen, G.C.; Yuan, H.W.; He, Y.; Yan, D.L.; Huang, J.Q.; Wang, X.F.; Zheng, B.S. Construction of Agrobacterium rhizogenes-mediated transformation system of Carya illinoinensis without dependence on tissue culture. J. Fruit Sci. 2022, 39, 131–140. [Google Scholar]
- Bahman, B.; Mohammad, N.; Rahul, B.; Sumita, J. A critical review on use of Agrobacterium rhizogenes and their associated binary vectors for plant transformation. Biotechnol. Adv. 2019, 37, 107405. [Google Scholar]
- Beigmohammadi, M.; Sharafi, A.; Jafari, S. An optimized protocol for Agrobacterium rhizogenes-mediated genetic transformation of Citrullus colocynthis. J. Appl. Biotechnol. Rep. 2019, 6, 113–117. [Google Scholar] [CrossRef] [Green Version]
- Desmet, S.; De, K.E.; Van, V.J.; Baeyen, S.; Van, H.J.; Geelen, D.; Dhooghe, E. Differential efficiency of wild type rhizogenic strains for rol gene transformation of plants. Appl. Microbiol. Biotechnol. 2019, 103, 6657–6672. [Google Scholar] [CrossRef]
- Sathasivam, R.; Choi, M.; Radhakrishnan, R.; Kwon, H.; Yoon, J.; Yang, S.H.; Kim, J.K.; Chung, Y.S.; Park, S.U. Effects of various Agrobacterium rhizogenes strains on hairy root induction and analyses of primary and secondary metabolites in Ocimum basilicum. Front. Plant Sci. 2022, 13, 983776. [Google Scholar] [CrossRef]
- Xiang, T.H.; Wang, S.S.; Wu, P.; Li, Y.F.; Zhang, T.; Wu, D.Z.; Zhou, S.Y. Cucumopine type Agrobacterium rhizogenes K599 (NCPPB2659) T-DNA mediated plant transformation and its application. Bangladesh J. Bot. 2016, 45, 935–945. [Google Scholar]
- Franco, J.A.V.; Collier, R.; Wang, Y.; Huo, N.X.; Gu, Y.; Thilmony, R.; Thomson, J.G. Draft genome sequence of Agrobacterium rhizogenes strain NCPPB2659. Genome Announc. 2016, 4, e00746-16. [Google Scholar]
- Castellanos-Arévalo, A.P.; Estrada-Luna, A.A.; Cabrera-Ponce, J.L.; Valencia-Lozano, E.; Herrera-Ubaldo, H.; Folter, S.; Blanco-Labra, A.; Délano-Frier, J.P. Agrobacterium rhizogenes-mediated transformation of grain (Amaranthus hypochondriacus) and leafy (A. hybridus) amaranths. Plant Cell Rep. 2020, 39, 1143–1160. [Google Scholar] [CrossRef] [PubMed]
- Faizal, A.; Manik, V.T.; Iriawati, I.; Esyanti, R.R. Establishment of hairy root cultures and analysis of saponin production in Javanese ginseng (Talinum paniculatum Gaertn). Acta Hortic. 2020, 1285, 193–202. [Google Scholar] [CrossRef]
- Gao, X.Q. Establishment and Application of Agrobacterium-Mediated Walnut Genetic Transformation System. Master’s Thesis, Northwest A&F University, Xianyang, China, 2021. [Google Scholar]
- Bandaranayake, P.C.G.; Yoder, J.I. Factors affecting the efficiency of Rhizobium rhizogenes root transformation of the root parasitic plant Triphysaria versicolor and its host Arabidopsis thaliana. BioMed Cent. 2018, 14, 61. [Google Scholar] [CrossRef] [Green Version]
- Rahman, L.U.; Kouno, H.; Hashiguchi, Y.; Yamamoto, H.; Narbad, A.; Parr, A.; Walton, N.; Ikenaga, T.; Kitamura, Y. HCHL expression in hairy roots of Beta vulgaris yields a high accumulation of p-hydroxybenzoic acid (pHBA) glucose ester, and linkage of pHBA into cell walls. Bioresour. Technol. 2009, 100, 4836–4842. [Google Scholar] [CrossRef]
- Mei, C. Establishment and Verification of Hairy Root Transformation System of Common Vetch (Vicia sativa L.). Master’s Thesis, Lanzhou University, Lanzhou, China, 2020. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, L.; Lu, M.; An, H. Generation of Composite Rosa roxburghii Plants with Transgenic Roots by Agrobacterium-Mediated Transformation. Horticulturae 2022, 8, 1079. https://doi.org/10.3390/horticulturae8111079
Gong L, Lu M, An H. Generation of Composite Rosa roxburghii Plants with Transgenic Roots by Agrobacterium-Mediated Transformation. Horticulturae. 2022; 8(11):1079. https://doi.org/10.3390/horticulturae8111079
Chicago/Turabian StyleGong, Lisha, Min Lu, and Huaming An. 2022. "Generation of Composite Rosa roxburghii Plants with Transgenic Roots by Agrobacterium-Mediated Transformation" Horticulturae 8, no. 11: 1079. https://doi.org/10.3390/horticulturae8111079
APA StyleGong, L., Lu, M., & An, H. (2022). Generation of Composite Rosa roxburghii Plants with Transgenic Roots by Agrobacterium-Mediated Transformation. Horticulturae, 8(11), 1079. https://doi.org/10.3390/horticulturae8111079