
Identification and Comparative Analysis of the Rosaceae RCI2 Gene Family and Characterization of the Cold Stress Response in Prunus mume




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Plants Genome Resources
2.3. Identification of RCI2s Gene Family
2.4. Gene Structure and Protein Conserved Motif Analysis
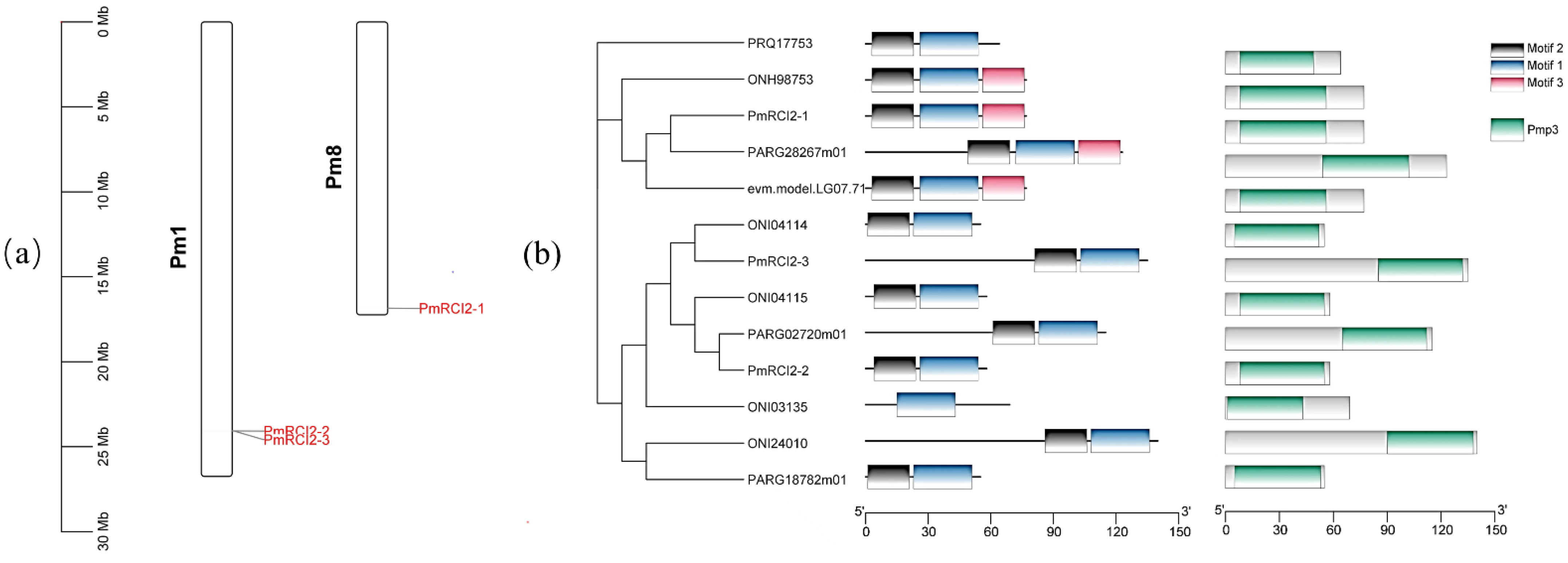
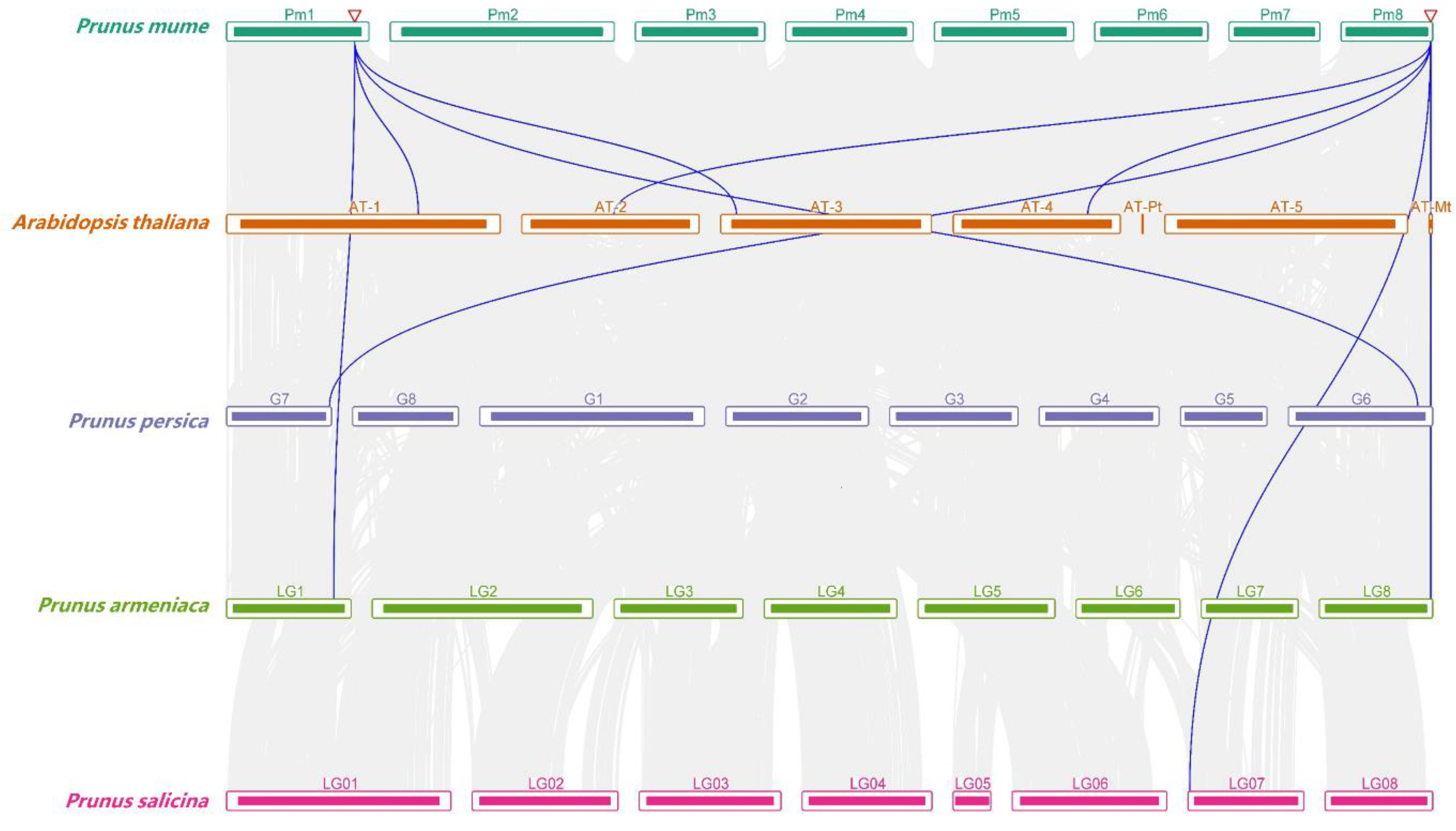
2.5. Chromosome Location and Synteny Analysis
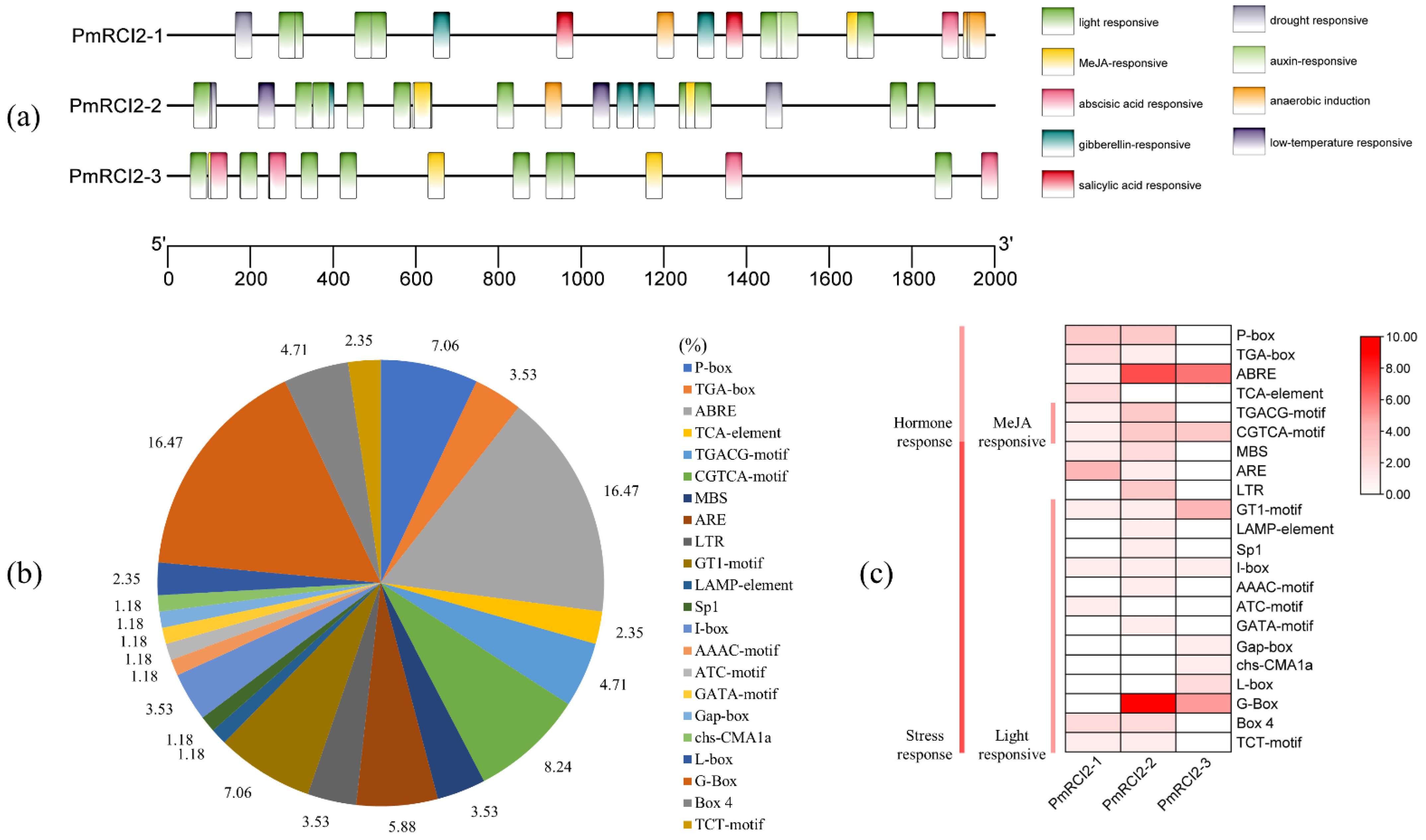
2.6. Cis-Acting Element Analysis of RCI2 Gene Promoters
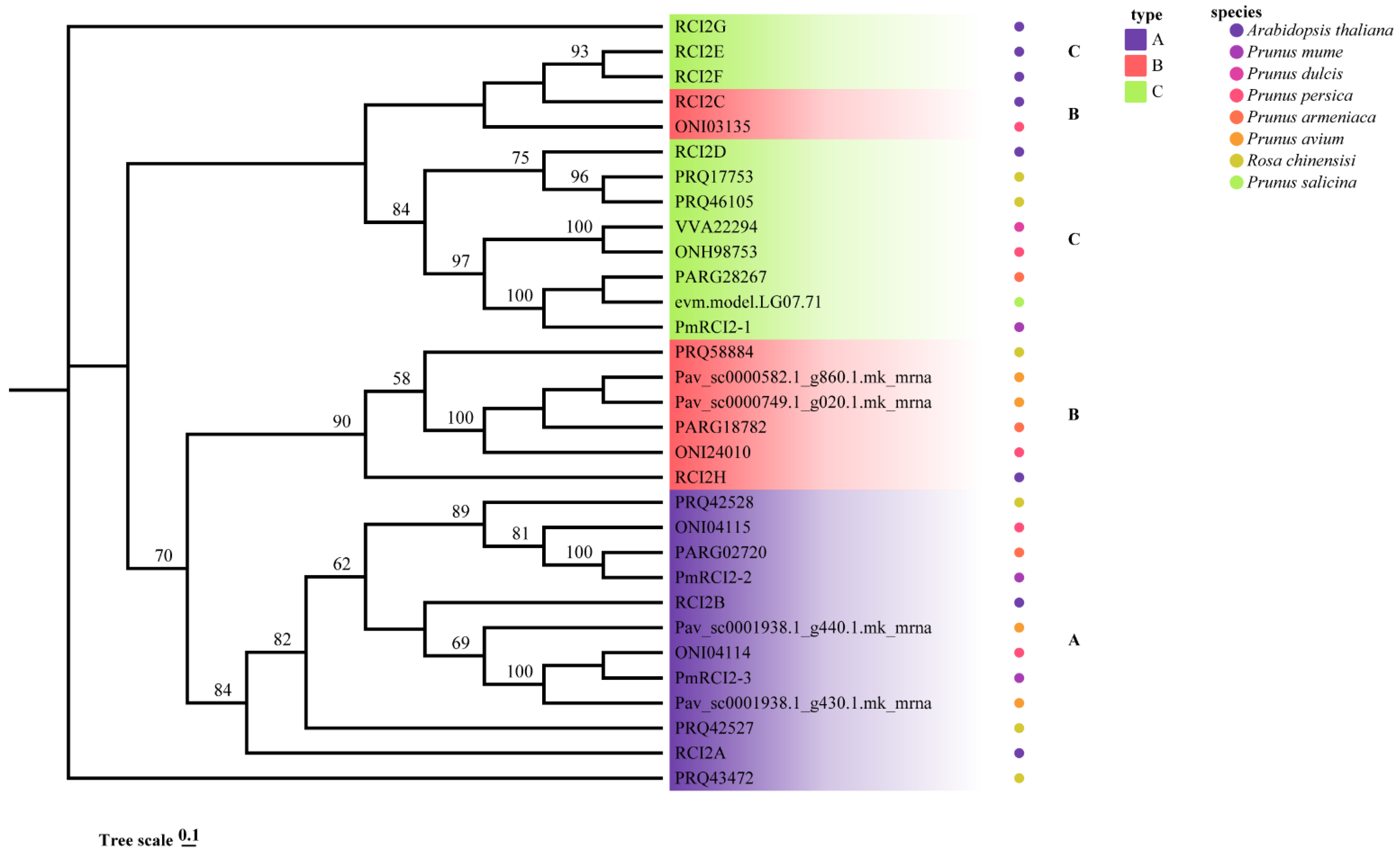
2.7. Phylogenetic Analysis of RCI2s
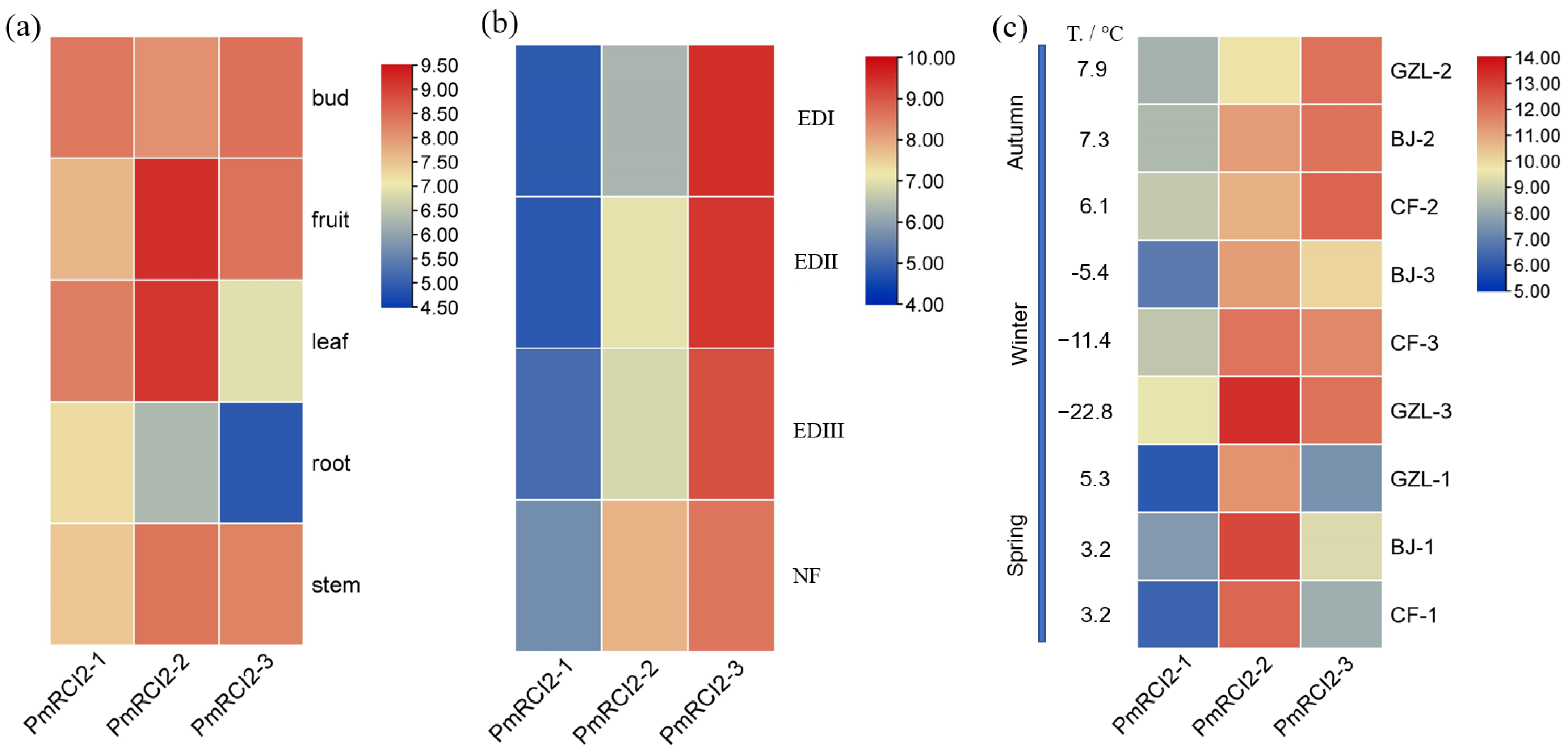
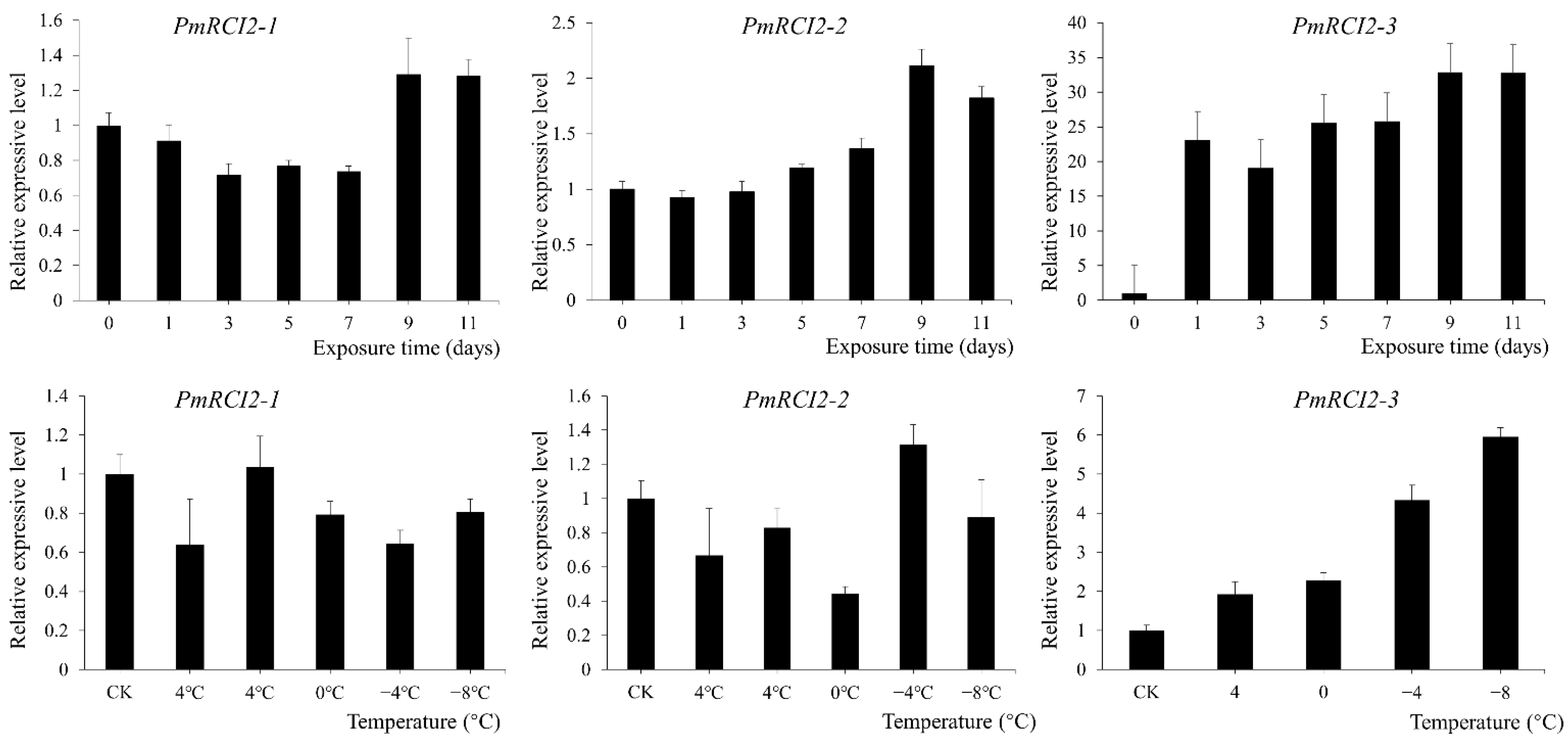
2.8. Expression Pattern of PmRCI2s
3. Results
3.1. Identification of Rosaceae RCI2s Gene Family
3.2. Gene Structure Analysis and Phylogenetic of RCI2 Genes
3.3. Analysis of Cis-Elements in the Promoters of PmRCI2s
3.4. Collinear Analysis of RCI2s in Rosaceae and A. thaliana
3.5. Expression Pattern of PmRCI2s under Cold Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weiser, C. Cold resistance and injury in woody plants: Knowledge of hardy plant adaptations to freezing stress may help us to reduce winter damage. Science 1970, 169, 1269–1278. [Google Scholar] [CrossRef]
- Yeshvekar, R.K.; Nitnavare, R.B.; Chakradhar, T.; Bhatnagar-Mathur, P.; Reddy, M.K.; Reddy, P.S. Molecular characterization and expression analysis of pearl millet plasma membrane proteolipid 3 (Pmp3) genes in response to abiotic stress conditions. Plant Gene 2017, 10, 37–44. [Google Scholar] [CrossRef]
- Navarre, C.; Goffeau, A. Membrane hyperpolarization and salt sensitivity induced by deletion of PMP3, a highly conserved small protein of yeast plasma membrane. EMBO J. 2000, 19, 2515–2524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, J.; Ballesteros, M.L.; Salinas, J. Phylogenetic and functional analysis of Arabidopsis RCI2 genes. J. Exp. Bot. 2007, 58, 4333–4346. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Ge, L.; Li, G.; He, P.; Yang, Y.; Liu, S. In silico identification and expression analysis of Rare Cold Inducible 2 (RCI2) gene family in cucumber. J. Plant Biochem. Biotechnol. 2020, 29, 56–66. [Google Scholar] [CrossRef]
- Won, S.Y.; Jung, J.A.; Kim, J.S. Genome-wide analysis of the MADS-Box gene family in Chrysanthemum. Comput. Biol. Chem. 2021, 90, 107424. [Google Scholar] [CrossRef]
- Mitsuya, S.; Taniguchi, M.; Miyake, H.; Takabe, T. Disruption of RCI2A leads to over-accumulation of Na+ and increased salt sensitivity in Arabidopsis thaliana plants. Planta 2005, 222, 1001–1009. [Google Scholar] [CrossRef]
- Fu, J.; Zhang, D.; Liu, Y.; Ying, S.; Shi, Y.; Song, Y.; Li, Y.; Wang, T. Isolation and characterization of maize PMP3 genes involved in salt stress tolerance. PLoS ONE 2012, 7, e31101. [Google Scholar] [CrossRef] [Green Version]
- Capel, J.; Jarillo, J.A.; Salinas, J.; Martinez-Zapater, J.M. Two homologous low-temperature-inducible genes from Arabidopsis encode highly hydrophobic proteins. Plant Physiol. 1997, 115, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Ben-Romdhane, W.; Ben-Saad, R.; Meynard, D.; Zouari, N.; Mahjoub, A.; Fki, L.; Guiderdoni, E.; Al-Doss, A.; Hassairi, A. Overexpression of AlTMP2 gene from the halophyte grass Aeluropus littoralis in transgenic tobacco enhances tolerance to different abiotic stresses by improving membrane stability and deregulating some stress-related genes. Protoplasma 2018, 255, 1161–1177. [Google Scholar] [CrossRef]
- Ben Romdhane, W.; Ben-Saad, R.; Meynard, D.; Verdeil, J.L.; Azaza, J.; Zouari, N.; Fki, L.; Guiderdoni, E.; Al-Doss, A.; Hassairi, A. Ectopic expression of Aeluropus littoralis plasma membrane protein gene AlTMP1 confers abiotic stress tolerance in transgenic tobacco by improving water status and cation homeostasis. Int. J. Mol. Sci. 2017, 18, 692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.O.; Kim, H.S.; Lim, H.G.; Jang, H.Y.; Kim, E.; Ahn, S.J. Functional characterization of salt-stress induced rare cold inducible gene from Camelina sativa (CsRCI2D). J. Plant Biol. 2021, 65, 279–289. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, J.Y.; Kim, S.J.; An, K.S.; An, G.; Kim, S.R. Isolation of cold stress-responsive genes in the reproductive organs, and characterization of the OsLti6b gene from rice (Oryza sativa L.). Plant Cell Rep. 2007, 26, 1097–1110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, H.; Sun, L.; Fan, G.; Ye, M.; Jiang, L.; Liu, X.; Ma, K.; Shi, C.; Bao, F. The genetic architecture of floral traits in the woody plant Prunus mume. Nat. Commun. 2018, 9, 1702. [Google Scholar] [CrossRef] [Green Version]
- Bolser, D.; Staines, D.M.; Pritchard, E.; Kersey, P. Ensembl plants: Integrating tools for visualizing, mining, and analyzing plant genomics data. In Plant Bioinformatics; Springer: Berlin/Heidelberg, Germany, 2016; pp. 115–140. [Google Scholar]
- Jung, S.; Lee, T.; Cheng, C.-H.; Buble, K.; Zheng, P.; Yu, J.; Humann, J.; Ficklin, S.P.; Gasic, K.; Scott, K. 15 years of GDR: New data and functionality in the Genome Database for Rosaceae. Nucleic Acids Res. 2019, 47, D1137–D1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Walsh, B. Population-genetic models of the fates of duplicate genes. In Origin and Evolution of New Gene Functions; Springer: Berlin/Heidelberg, Germany, 2003; pp. 279–294. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2003. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, W.; Sun, L.; Zhao, F.; Huang, B.; Yang, W.; Tao, Y.; Wang, J.; Yuan, Z.; Fan, G. The genome of Prunus mume. Nat. Commun. 2012, 3, 1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Zhuo, X.; Zhao, K.; Zheng, T.; Han, Y.; Yuan, C.; Zhang, Q. Transcriptome profiles reveal the crucial roles of hormone and sugar in the bud dormancy of Prunus mume. Sci. Rep. 2018, 8, 5090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Lu, J.; Xu, Z.; Yang, W.; Wang, J.; Cheng, T.; Zhang, Q. Selection of suitable reference genes for miRNA expression normalization by qRT-PCR during flower development and different genotypes of Prunus mume. Sci. Hortic. 2014, 169, 130–137. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, J.; Pan, Y.; Li, J.; Shi, H.; Zeng, Y.; Guo, H.; Yang, S.; Zheng, W.; Yu, J. Natural variation in CTB4a enhances rice adaptation to cold habitats. Nat. Commun. 2017, 8, 14788. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Hakim, K.; Nahar, K.; Alharby, H.F. Plant Abiotic Stress Tolerance; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, J.E.; Jang, H.Y.; Kwak, K.J.; Ahn, S.J. CsRCI2A and CsRCI2E genes show opposite salt sensitivity reaction due to membrane potential control. Acta Physiol. Plant. 2016, 38, 50. [Google Scholar] [CrossRef]
- Rocha, P.S. Plant abiotic stress-related RCI2/PMP3s: Multigenes for multiple roles. Planta 2016, 243, 1–12. [Google Scholar] [CrossRef]
- Sun, W.; Li, M.; Wang, J. Genome-wide identification and characterization of the RCI2 gene family in allotetraploid Brassica napus compared with its diploid progenitors. Int. J. Mol. Sci. 2022, 23, 614. [Google Scholar] [CrossRef]
- Vishwakarma, K.; Upadhyay, N.; Kumar, N.; Yadav, G.; Singh, J.; Mishra, R.K.; Kumar, V.; Verma, R.; Upadhyay, R.; Pandey, M. Abscisic acid signaling and abiotic stress tolerance in plants: A review on current knowledge and future prospects. Front. Plant Sci. 2017, 8, 161. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene ID | Basic Features | |||
|---|---|---|---|---|---|
| pI | MW/kDa | Length/aa | Prediction of Subcellular Localization | ||
| Prunus dulcis | VVA22294 | 7.87 | 12.37 | 118 | Plasma membrane |
| Prunus persica | ONH98753 | 5.09 | 8.53 | 77 | Vacuole |
| ONI04115 | 5.43 | 6.33 | 58 | Plasma membrane | |
| ONI04114 | 4.68 | 6.01 | 55 | Vacuole | |
| ONI03135 | 9.69 | 7.33 | 69 | Plasma membrane | |
| ONI24010 | 7.81 | 13.94 | 140 | Plasma membrane | |
| Prunus armeniaca | PARG28267m01 | 6.15 | 14.13 | 123 | Plasma membrane |
| PARG02720m01 | 9.95 | 12.73 | 115 | Mitochondrion | |
| PARG18782m01 | 4.25 | 5.93 | 55 | Vacuole | |
| Prunus avium | Pav_sc0001938.1_g430.1.mk:mrna | 4.68 | 6.01 | 55 | Vacuole |
| Pav_sc0001938.1_g440.1.mk:mrna | 6.5 | 6.03 | 55 | Plasma membrane | |
| Pav_sc0000749.1_g020.1.mk:mrna | 8.79 | 15.74 | 149 | Plasma membrane | |
| Pav_sc0000582.1_g860.1.mk:mrna | 8.86 | 15.12 | 136 | Plasma membrane | |
| Rosa chinensis | PRQ17753 | 4.75 | 6.6 | 64 | Vacuole |
| PRQ42527 | 5.82 | 6.02 | 55 | Vacuole | |
| PRQ42528 | 5.93 | 6.36 | 58 | Vacuole | |
| PRQ58884 | 4.25 | 5.85 | 55 | Vacuole | |
| PRQ43472 | 4.09 | 7.53 | 71 | Vacuole | |
| PRQ46105 | 5.01 | 8.59 | 79 | Vacuole | |
| Prunus mume | Pm027750 | 4.79 | 8.65 | 77 | Vacuole |
| Pm003262 | 5.43 | 6.35 | 58 | Plasma membrane | |
| Pm003263 | 9.58 | 15.27 | 135 | Vacuole | |
| Prunus salicina | evm.model.LG07.71 | 4.79 | 8.65 | 77 | Vacuole |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Li, P.; Qiu, L.; Ahmad, S.; Wang, J.; Zheng, T. Identification and Comparative Analysis of the Rosaceae RCI2 Gene Family and Characterization of the Cold Stress Response in Prunus mume. Horticulturae 2022, 8, 997. https://doi.org/10.3390/horticulturae8110997
Yang L, Li P, Qiu L, Ahmad S, Wang J, Zheng T. Identification and Comparative Analysis of the Rosaceae RCI2 Gene Family and Characterization of the Cold Stress Response in Prunus mume. Horticulturae. 2022; 8(11):997. https://doi.org/10.3390/horticulturae8110997
Chicago/Turabian StyleYang, Lichen, Ping Li, Like Qiu, Sagheer Ahmad, Jia Wang, and Tangchun Zheng. 2022. "Identification and Comparative Analysis of the Rosaceae RCI2 Gene Family and Characterization of the Cold Stress Response in Prunus mume" Horticulturae 8, no. 11: 997. https://doi.org/10.3390/horticulturae8110997
APA StyleYang, L., Li, P., Qiu, L., Ahmad, S., Wang, J., & Zheng, T. (2022). Identification and Comparative Analysis of the Rosaceae RCI2 Gene Family and Characterization of the Cold Stress Response in Prunus mume. Horticulturae, 8(11), 997. https://doi.org/10.3390/horticulturae8110997