



The Impact of Viticulture on Soil Characteristics and Microbial Communities in the Ningxia Region of Northwest China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design and Soil Collection
2.3. Soil Physicochemical Analyses
2.4. Soil Enzyme Activity Analyses
2.5. DNA Extraction, Amplification and Sequencing
2.6. Sequencing Data Processing
2.7. Statistical Analyses
3. Results
3.1. Soil Properties and Enzyme Activities
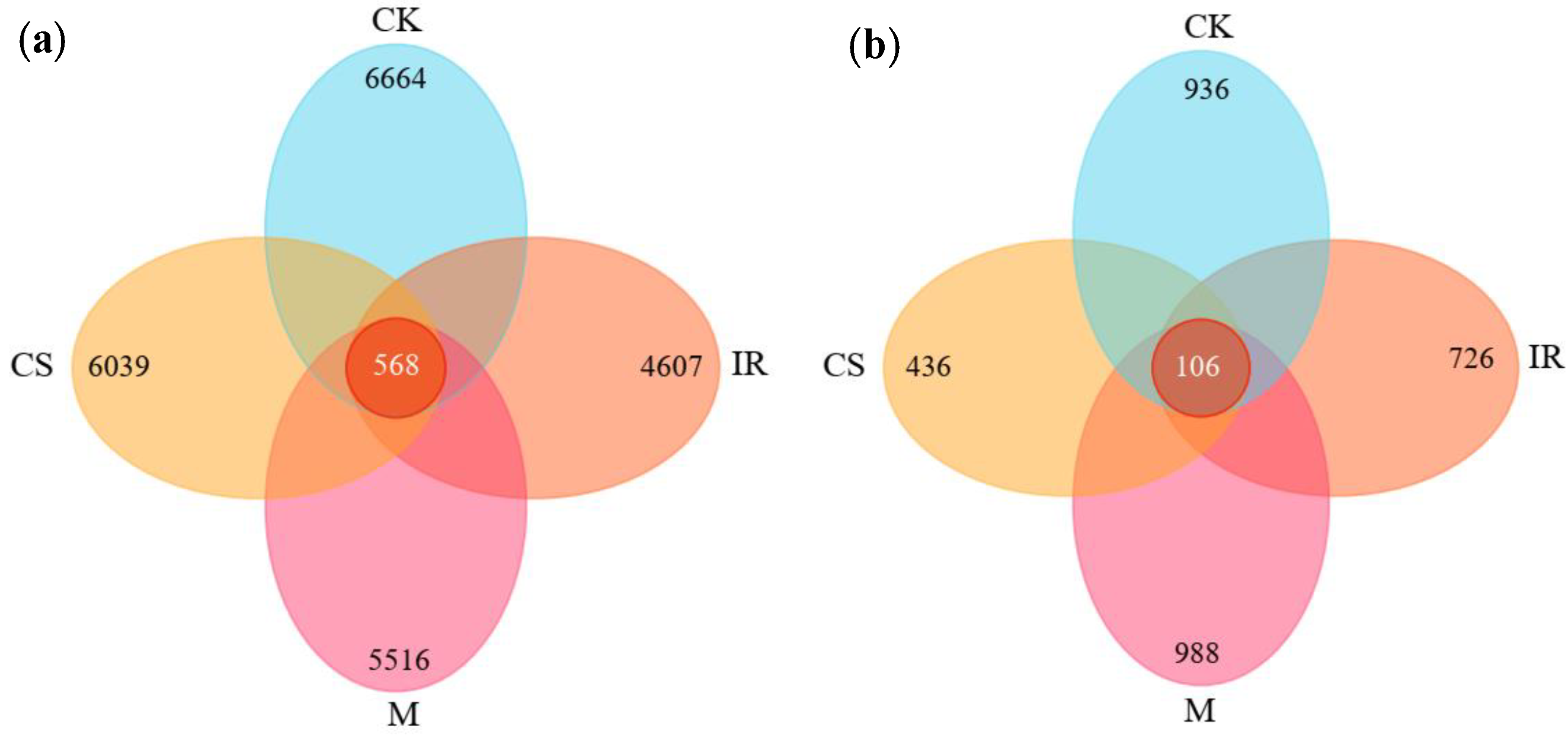
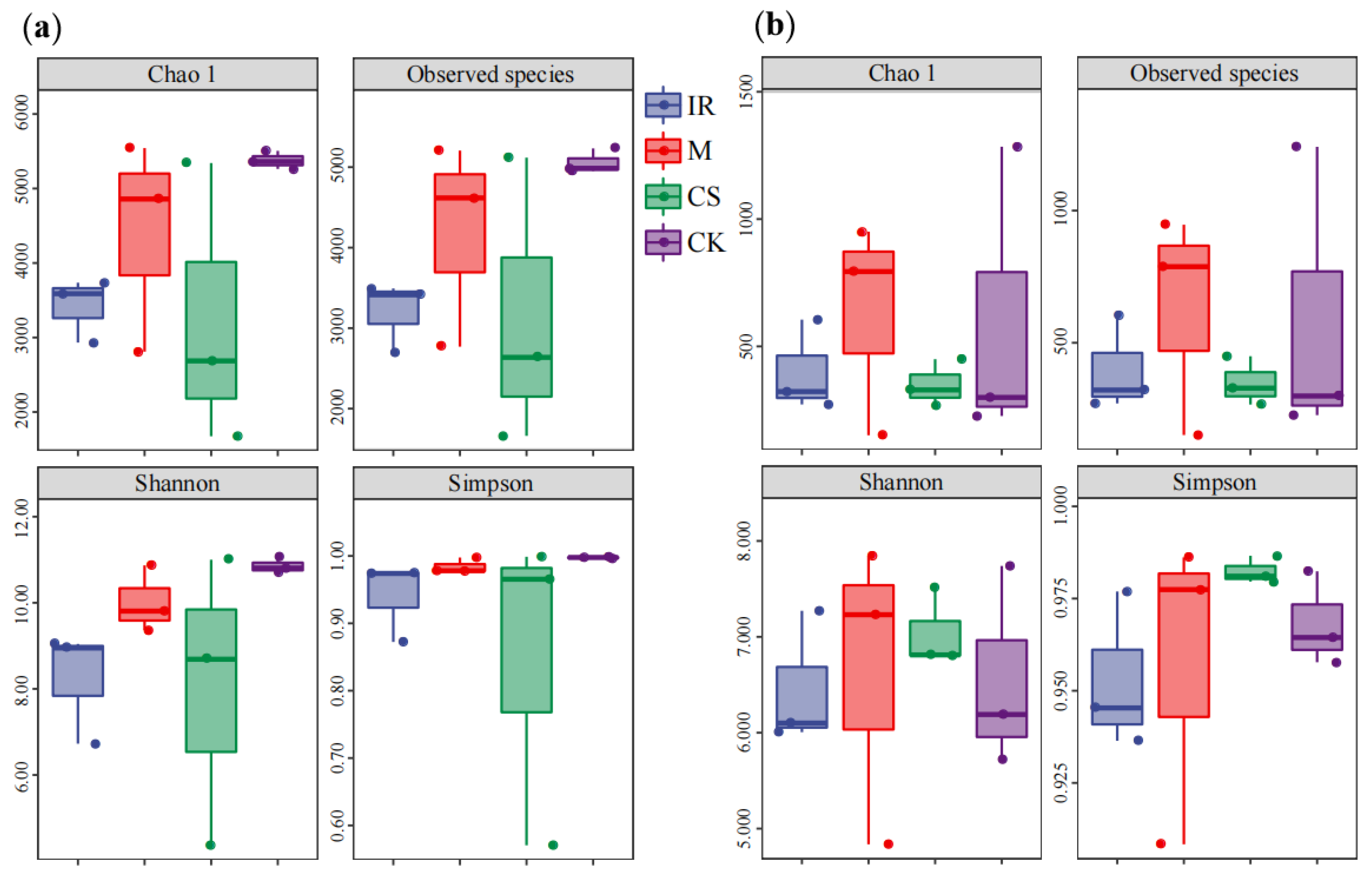
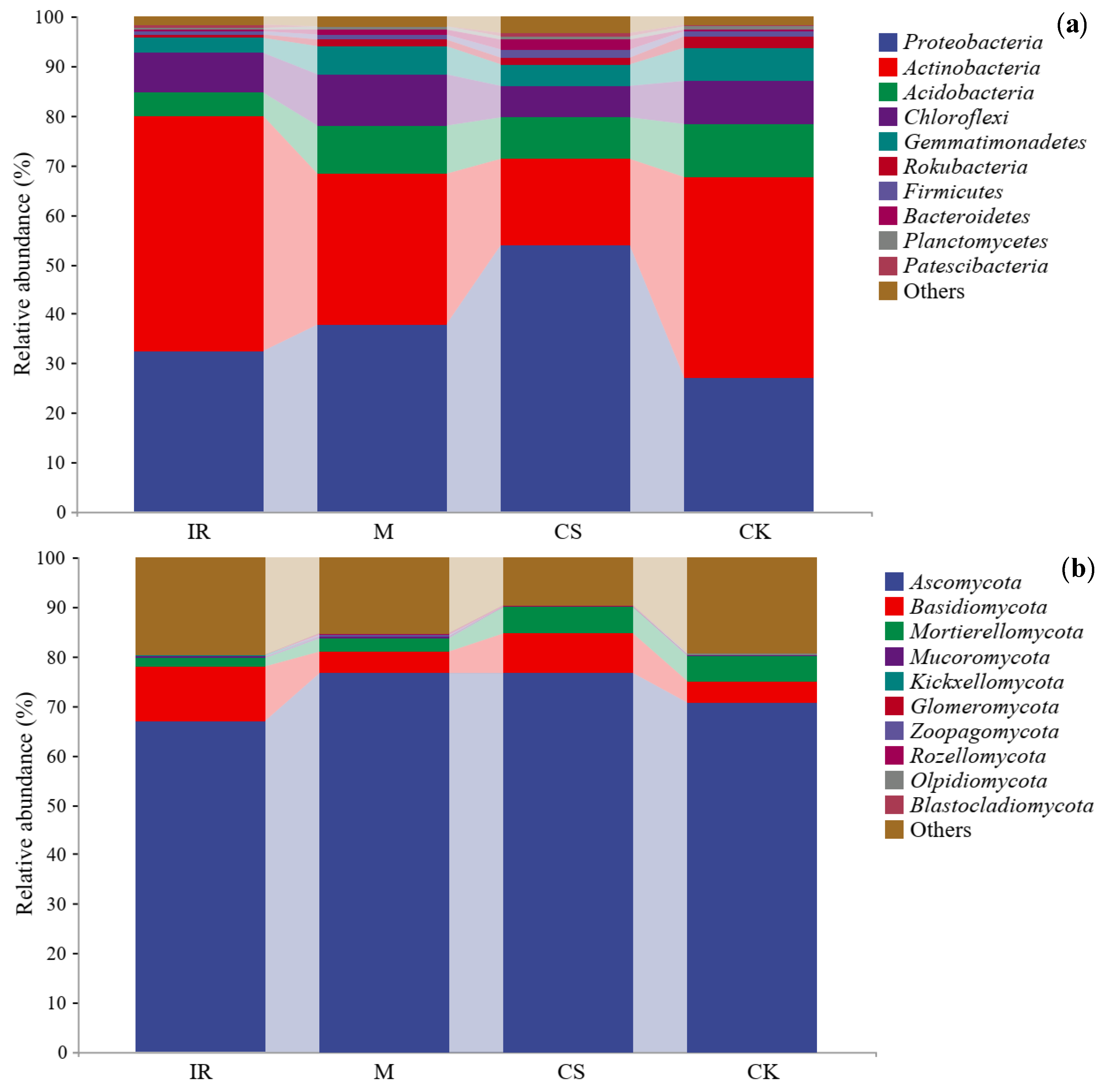
3.2. Characterization of the Soil Bacterial and Fungal Communities in Different Sample Plots
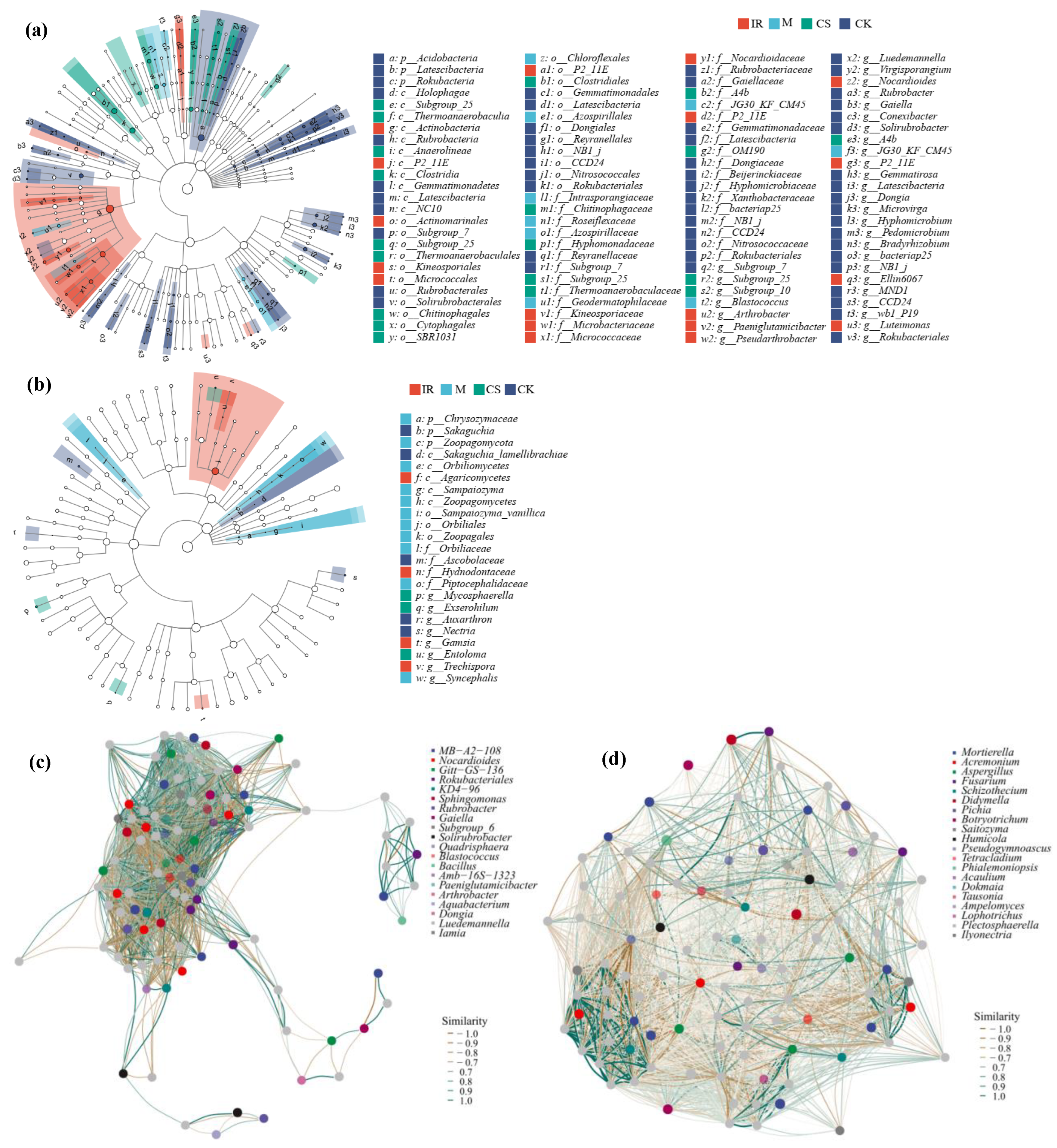
3.3. Differences in Soil Bacterial and Fungal Community Structures in Different Sample Plots
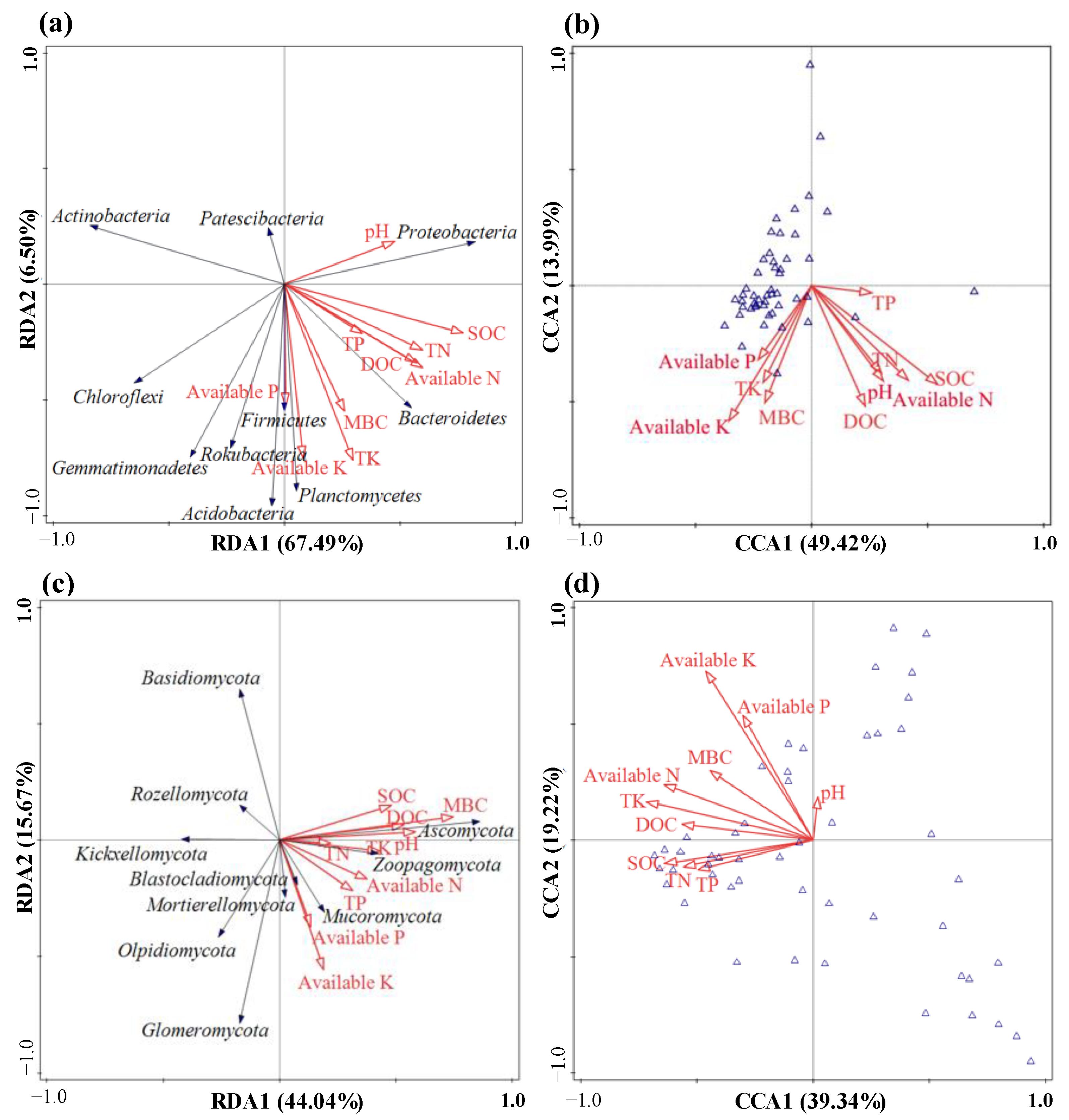
3.4. Soil Microbial Community Responses to Soil Properties and Enzyme Activity Levels
4. Discussion
4.1. Soil Properties and Enzyme Activity Levels
4.2. Soil Bacteria
4.3. Soil Fungi
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, L.; Wang, Z.; Xue, T.; Gao, F.; Wei, R.; Wang, Y.; Han, X.; Li, H.; Wang, H. Combating desertification through the wine industry in Hongsibu, Ningxia. Sustainability 2021, 13, 5654. [Google Scholar] [CrossRef]
- Yang, B.; Yao, H.; Zhang, J.; Li, Y.; Ju, Y.; Zhao, X.; Sun, X.; Fang, Y. Effect of regulated deficit irrigation on the content of soluble sugars, organic acids and endogenous hormones in Cabernet Sauvignon in the Ningxia region of China. Food Chem. 2020, 312, 126020. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, Q.; Xi, B.; Dai, H. Study on the regulation of anthocyanin biosynthesis by exogenous abscisic acid in grapevine. Sci Hortic. 2019, 250, 294–301. [Google Scholar] [CrossRef]
- Li, L.; Sun, B. Grape and wine polymeric polyphenols: Their importance in enology. Crit. Rev. Food Sci. 2019, 59, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Marsal, J.; Mata, M.; Del Campo, J.; Arbones, A.; Vallverdu, X.; Girona, J.; Olivo, N. Evaluation of partial root-zone drying for potential field use as a deficit irrigation technique in commercial vineyards according to two different pipeline layouts. Irrigation Sci. 2008, 26, 347–356. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, S.; Li, L.; Inoue, M.; Xiang, J.; Qiu, G.; Jin, W. Effects of mulching and sub-surface irrigation on vine growth, berry sugar content and water use of grapevines. Agric. Water Manag. 2014, 143, 1–8. [Google Scholar] [CrossRef]
- Jaksic, S.; Ninkov, J.; Milic, S.; Vasin, J.; Banjac, D.; Jaksic, D.; Zivanov, M. The state of soil organic carbon in vineyards as affected by soil types and fertilization strategies (Tri Morave Region, Serbia). Agronomy 2021, 11, 9. [Google Scholar] [CrossRef]
- Rusch, A.; Delbac, L.; Thiery, D. Grape moth density in Bordeaux vineyards depends on local habitat management despite effects of landscape heterogeneity on their biological control. J. Appl. Ecol. 2017, 54, 1794–1803. [Google Scholar] [CrossRef] [Green Version]
- Gougeon, L.; Da Costa, G.; Le Mao, I.; Ma, W.; Teissedre, P.; Guyon, F.; Richard, T. Wine analysis and authenticity using H-1-NMR metabolomics data: Application to Chinese wines. Food Anal. Methods 2018, 11, 3425–3434. [Google Scholar] [CrossRef]
- Parga-Dans, E.; Alonso Gonzalez, P.; Otero-Enriquez, R. The role of expert judgments in wine quality assessment: The mismatch between chemical, sensorial and extrinsic cues. Brit. Food J. 2022, 124, 4286–4303. [Google Scholar] [CrossRef]
- Wang, X.; Xie, X.; Chen, N.; Wang, H.; Li, H. Study on current status and climatic characteristics of wine regions in China. Vitis 2018, 57, 9–16. [Google Scholar]
- Oliver, D.P.; Bramley, R.G.V.; Riches, D.; Porter, I.; Edwards, J. Review: Soil physical and chemical properties as indicators of soil quality in Australian viticulture. Aust. J. Grape Wine Res. 2013, 19, 129–139. [Google Scholar] [CrossRef]
- Mouazen, A.M.; Steffens, M.; Borisover, M. Reflectance and fluorescence spectroscopy in soil science-Current and future research and developments. Soil Till. Res. 2016, 155, 448–449. [Google Scholar] [CrossRef]
- Bunemann, E.K.; Bongiorno, G.; Bai, Z.; Creamer, R.E.; De Deyn, G.; de Goede, R.; Fleskens, L.; Geissen, V.; Kuyper, T.W.; Mader, P.; et al. Soil quality—A critical review. Soil Biol. Biochem. 2018, 120, 105–125. [Google Scholar] [CrossRef]
- Morvan, X.; Saby, N.P.A.; Arrouays, D.; Le Bas, C.; Jones, R.J.A.; Verheijen, F.G.A.; Bellamy, P.H.; Stephens, M.; Kibblewhite, M.G. Soil monitoring in Europe: A review of existing systems and requirements for harmonisation. Sci. Total Environ. 2008, 391, 1–12. [Google Scholar] [CrossRef]
- Sharma, N.K.; Singh, R.J.; Mandal, D.; Kumar, A.; Alam, N.M.; Keesstra, S. Increasing farmer’s income and reducing soil erosion using intercropping in rainfed maize-wheat rotation of Himalaya, India. Agric. Ecosyst. Environ. 2017, 247, 43–53. [Google Scholar] [CrossRef]
- Eivazi, F.; Bayan, M.R.; Schmidt, K. Select soil enzyme activities in the historic Sanborn Field as affected by long-term cropping systems. Commun. Soil Sci. Plan. 2003, 34, 2259–2275. [Google Scholar] [CrossRef]
- Crecchio, C.; Curci, M.; Pellegrino, A.; Ricciuti, P.; Tursi, N.; Ruggiero, P. Soil microbial dynamics and genetic diversity in soil under monoculture wheat grown in different long-term management systems. Soil Biol. Biochem. 2007, 39, 1391–1400. [Google Scholar] [CrossRef]
- Bian, T.; Zheng, S.; Li, X.; Wang, S.; Zhang, X.; Wang, Z.; Li, X.; Fu, H.; Sun, Z. The variation of soil phosphorus fractions and microbial community composition under consecutive cucumber cropping in a greenhouse. Horticulturae 2022, 8, 320. [Google Scholar] [CrossRef]
- Xu, H.W.; Qu, Q.; Lu, B.B.; Li, P.; Xue, S.; Liu, G.B. Response of soil specific enzyme activity to vegetation restoration in the Loess hilly region of China. Catena 2020, 191, 104564. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, X.; Wang, X.; Shao, H.; Yang, J.; Wang, X. Soil enzymes as indicators of saline soil fertility under various soil amendments. Agric. Ecosyst. Environ. 2017, 237, 274–279. [Google Scholar]
- Trasar-Cepeda, C.; Leiros, M.C.; Gil-Sotres, F. Hydrolytic enzyme activities in agricultural and forest soils. Some implications for their use as indicators of soil quality. Soil Biol. Biochem. 2008, 40, 2146–2155. [Google Scholar] [CrossRef]
- Wang, B.; Xue, S.; Liu, G.B.; Zhang, G.H.; Li, G.; Ren, Z.P. Changes in soil nutrient and enzyme activities under different vegetations in the Loess Plateau area, Northwest China. Catena 2012, 92, 186–195. [Google Scholar] [CrossRef]
- Cui, J.; Liu, C.; Li, Z.; Wang, L.; Chen, X.; Ye, Z.; Fang, C. Long-term changes in topsoil chemical properties under centuries of cultivation after reclamation of coastal wetlands in the Yangtze Estuary, China. Soil. Till. Res. 2012, 123, 50–60. [Google Scholar] [CrossRef]
- Xie, X.; Pu, L.; Wang, Q.; Zhu, M.; Xu, Y.; Zhang, M. Response of soil physicochemical properties and enzyme activities to long-term reclamation of coastal saline soil, Eastern China. Sci. Total Environ. 2017, 607, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Qi, R.; Li, J.; Lin, Z.; Li, Z.; Li, Y.; Yang, X.; Zhang, J.; Zhao, B. Temperature effects on soil organic carbon, soil labile organic carbon fractions, and soil enzyme activities under long-term fertilization regimes. Appl. Soil Ecol. 2016, 102, 36–45. [Google Scholar] [CrossRef]
- Li, J.; Xie, T.; Zhu, H.; Zhou, J.; Li, C.; Xiong, W.; Xu, L.; Wu, Y.; He, Z.; Li, X. Alkaline phosphatase activity mediates soil organic phosphorus mineralization in a subalpine forest ecosystem. Geoderma 2021, 404, 115376. [Google Scholar] [CrossRef]
- Acosta-Martinez, V.; Cruz, L.; Sotomayor-Ramirez, D.; Perez-Alegria, L. Enzyme activities as affected by soil properties and land use in a tropical watershed. Appl. Soil Ecol. 2007, 35, 35–45. [Google Scholar] [CrossRef]
- Li, J.; Liu, G.; Kwak, J.; Chang, S.X.; Gao, H.; Wu, Q.; Yang, J.; Chen, J. Reclamation of desert land to continuous cotton cropping affects soil properties and microbial communities in the desert-oasis ecotone of Xinjiang, China. J. Soils Sediments 2020, 20, 862–873. [Google Scholar] [CrossRef]
- Franco-Otero, V.G.; Soler-Rovira, P.; Hernandez, D.; Lopez-de-Sa, E.G.; Plaza, C. Short-term effects of organic municipal wastes on wheat yield, microbial biomass, microbial activity, and chemical properties of soil. Biol. Fertil. Soils 2012, 48, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Szoboszlay, M.; Dohrmann, A.B.; Poeplau, C.; Don, A.; Tebbe, C.C. Impact of land-use change and soil organic carbon quality on microbial diversity in soils across Europe. FEMS Microbiol. Ecol. 2017, 93, fix146. [Google Scholar] [CrossRef] [Green Version]
- Tosi, M.; Correa, O.S.; Soria, M.A.; Vogrig, J.A.; Sydorenko, O.; Montecchia, M.S. Land-use change affects the functionality of soil microbial communities: A chronosequence approach in the Argentinian Yungas. Appl. Soil Ecol. 2016, 108, 118–127. [Google Scholar] [CrossRef]
- Wu, W.; Dong, C.; Wu, J.; Liu, X.; Wu, Y.; Chen, X.; Yu, S. Ecological effects of soil properties and metal concentrations on the composition and diversity of microbial communities associated with land use patterns in an electronic waste recycling region. Sci. Total Environ. 2017, 601, 57–65. [Google Scholar] [CrossRef]
- Zhang, L.; Xue, T.; Yuan, L.; Gao, F.; Hao, X.; Yang, C.; Wang, L.; Han, Y.; Li, H.; Wang, H. The effect of vineyard reclamation on soil properties and microbial communities in desertified land in Hongsibu, Ningxia. Catena 2022, 211, 106002. [Google Scholar] [CrossRef]
- Li, J.; Pokharel, P.; Liu, G.; Chen, J. Reclamation of desert land to different land-use types changes soil bacterial community composition in a desert-oasis ecotone. Land Degrad. Dev. 2021, 32, 1389–1399. [Google Scholar] [CrossRef]
- Li, C.Y.; Zheng, L. Improvement of simultaneous and rapid determination of total nitrogen and total phosphorus in plants by flow analyzer. Soil Fertil. Sci. China 2021, 2, 336–342. [Google Scholar]
- Bao, S.D. Agriculture and Chemistry Analysis of Soil; China Agriculture Press: Beijing, China, 2000; pp. 39–80. [Google Scholar]
- Peng, Q.; Li, X.H.; Wang, K.A.; Lv, X.H.; Yang, N.; Liu, X.F. Effect of reduced controlled release nitrogen fertilizer on the sweet pepper yield and distribution of soil nitrate, ammonium nitrogen. J. Soil Water Conserv. 2012, 26, 106–110. [Google Scholar]
- Lin, Q.M.; Wu, Y.G.; Liu, H.L. Modification of fumigation extraction method for measuring soil microbial biomass carbon. Chin. J. Ecol. 1999, 18, 63–66. [Google Scholar]
- Zhang, L.; Xue, T.; Gao, F.; Wei, R.; Wang, Z.; Li, H.; Wang, H. Carbon storage distribution characteristics of vineyard ecosystems in Hongsibu, Ningxia. Plants 2021, 10, 1199. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Ca, L.Q.; Wu, J.; Qi, P.; Luo, Z.Z.; Zhang, R.Z. The relationship between soil labile organic carbon fractions and the enzyme activities under different tillage measures in the Loess Plateau of central Gansu province. Agric. Res. Arid. Areas 2017, 35, 1–7. [Google Scholar]
- White, T.; Bruns, T.D.; Lee, S.; Taylor, J. Amplification direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Ed.; Sigma Aldrich: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemp, P.F.; Aller, J.Y. Bacterial diversity in aquatic and other environments: What 16S rDNA libraries can tell us. Fems. Microbiol. Ecol. 2004, 47, 161–177. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Gloeckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Koljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Djukic, I.; Zehetner, F.; Tatzber, M.; Gerzabek, M.H. Soil organic-matter stocks and characteristics along an Alpine elevation gradient. J. Plant Nutr. Soil Sci. 2010, 173, 30–38. [Google Scholar] [CrossRef]
- Lal, R. Restoring soil quality to mitigate soil degradation. Sustainability 2015, 7, 5875–5895. [Google Scholar] [CrossRef] [Green Version]
- Domínguez, M.T.; Panettieri, M.; Madejón, E.; Madejón, P. Thistle crops in marginal lands after compost addition: Plant biomass and effect on soil physical, chemical and biological properties. Land Degrad. Dev. 2020, 31, 1167–1175. [Google Scholar] [CrossRef]
- Zhang, W.; Long, X.; Huo, X.; Chen, Y.; Lou, K. 16S rRNA-Based PCR-DGGE analysis of actinomycete communities in fields with continuous cotton cropping in Xinjiang, China. Microb. Ecol. 2013, 66, 385–393. [Google Scholar] [CrossRef]
- Cenini, V.L.; Fornara, D.A.; McMullan, G.; Ternan, N.; Carolan, R.; Crawley, M.J.; Clement, J.; Lavorel, S. Linkages between extracellular enzyme activities and the carbon and nitrogen content of grassland soils. Soil Biol. Biochem. 2016, 96, 198–206. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson, J.; Ahnstrom, J.; Weibull, A.C. The effects of organic agriculture on biodiversity and abundance: A meta-analysis. J. Appl. Ecol. 2005, 42, 261–269. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Maeder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [Green Version]
- Nacke, H.; Thuermer, A.; Wollherr, A.; Will, C.; Hodac, L.; Herold, N.; Schoening, I.; Schrumpf, M.; Daniel, R. Pyrosequencing-based assessment of bacterial community structure along different management types in German forest and grassland soils. PLoS ONE 2011, 6, e17000. [Google Scholar] [CrossRef] [Green Version]
- Xiong, W.; Li, Z.; Liu, H.; Xue, C.; Zhang, R.; Wu, H.; Li, R.; Shen, Q. The effect of long-term continuous cropping of black pepper on soil bacterial communities as determined by 454 pyrosequencing. PLoS ONE 2015, 10, e0136946. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Zhang, F.; Gale, W.J.; Wang, W.; Sang, W.; Yang, H. Effects of reclamation years on composition and diversity of soil bacterial communities in Northwest China. Can. J. Microbiol. 2018, 64, 28–40. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yang, Z.; Dang, P.; Zhu, H.; Gao, Y.; Vu, N.H.; Zhao, Z. Response of soil microbial community dynamics to Robinia pseudoacacia L. afforestation in the loess plateau: A chronosequence approach. Plant Soil 2018, 423, 327–338. [Google Scholar] [CrossRef]
- Kerfahi, D.; Tripathi, B.M.; Dong, K.; Go, R.; Adams, J.M. Rainforest conversion to rubber plantation may not result in lower soil diversity of bacteria, fungi, and nematodes. Microb. Ecol. 2016, 72, 359–371. [Google Scholar] [CrossRef]
- Tian, Q.; Taniguchi, T.; Shi, W.; Li, G.; Yamanaka, N.; Du, S. Land-use types and soil chemical properties influence soil microbial communities in the semiarid Loess Plateau region in China. Sci. Rep. 2017, 7, 45289. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Chi, Z.; Li, J.; Wu, H.; Yan, B. Bacterial community structure and function in soils from tidal freshwater wetlands in a Chinese delta: Potential impacts of salinity and nutrient. Sci. Total Environ. 2019, 696, 134029. [Google Scholar] [CrossRef]
- Cai, W.; Li, Y.; Wang, P.; Niu, L.; Zhang, W.; Wang, C. Revealing the relationship between microbial community structure in natural biofilms and the pollution level in urban rivers: A case study in the Qinhuai River basin, Yangtze River Delta. Water Sci. Technol. 2016, 74, 1163–1176. [Google Scholar] [CrossRef] [Green Version]
- Zumsteg, A.; Luster, J.; Goeransson, H.; Smittenberg, R.H.; Brunner, I.; Bernasconi, S.M.; Zeyer, J.; Frey, B. Bacterial, archaeal and fungal succession in the forefield of a receding glacier. Microb. Ecol. 2012, 63, 552–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, K.S.; Leff, J.W.; Barberán, A.; Bates, S.T.; Betley, J.; Crowther, T.W.; Kelly, E.F.; Oldfield, E.E.; Shaw, E.A.; Steenbock, C.; et al. Biogeographic patterns in below-ground diversity in New York City’s Central Park are similar to those observed globally. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141988. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlova, Y.E.; Kostina, N.V.; Gorlenko, M.V.; Umarov, M.M. Structural and functional features of microbial communities in a soddy-podzolic soil under self-remediation conditions. Mosc. Univ. Soil Sci. Bull. 2008, 63, 184–189. [Google Scholar] [CrossRef]
- Maestre, F.T.; Delgado-Baquerizo, M.; Jeffries, T.C.; Eldridge, D.J.; Ochoa, V.; Gozalo, B.; Quero, J.L.; García-Gómez, M.; Gallardo, A.; Ulrich, W.; et al. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc. Natl. Acad. Sci. USA 2015, 112, 15684–15689. [Google Scholar] [CrossRef] [Green Version]
- Porras-Alfaro, A.; Herrera, J.; Natvig, D.O.; Lipinski, K.; Sinsabaugh, R.L. Diversity and distribution of soil fungal communities in a semiarid grassland. Mycologia 2011, 103, 10–21. [Google Scholar] [CrossRef]
- Prober, S.M.; Leff, J.W.; Bates, S.T.; Borer, E.T.; Firn, J.; Harpole, W.S.; Lind, E.M.; Seabloom, E.W.; Adler, P.B.; Bakker, J.D.; et al. Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecol. Lett. 2015, 18, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Ng, E.L.; Patti, A.F.; Rose, M.T.; Schefe, C.R.; Wilkinson, K.; Smernik, R.J.; Cavagnaro, T.R. Does the chemical nature of soil carbon drive the structure and functioning of soil microbial communities? Soil Biol. Biochem. 2014, 70, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Janusz, G.; Pawlik, A.; Sulej, J.; Swiderska-Burek, U.; Jarosz-Wilkolazka, A.; Paszczynski, A. Lignin degradation: Microorganisms, enzymes involved, genomes analysis and evolution. Fems. Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, Y.; Chang, S.X.; Liang, X.; Qin, H.; Chen, J.; Xu, Q. Linking soil fungal community structure and function to soil organic carbon chemical composition in intensively managed subtropical bamboo forests. Soil Biol. Biochem. 2017, 107, 19–31. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, B.; Zheng, S.; Zhang, X.; Wang, X.; Dong, W.; Xie, Q.; Wang, G.; Xiao, Y.; Chen, F.; et al. A phosphate starvation response-centered network regulates mycorrhizal symbiosis. Cell 2021, 184, 5527. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Si, H.; He, J.; Fan, L.; Li, L. The shifts of maize soil microbial community and networks are related to soil properties under different organic fertilizers. Rhizosphere 2021, 19, 100388. [Google Scholar] [CrossRef]
- Talbot, J.M.; Allison, S.D.; Treseder, K.K. Decomposers in disguise: Mycorrhizal fungi as regulators of soil C dynamics in ecosystems under global change. Funct. Ecol. 2008, 22, 955–963. [Google Scholar] [CrossRef] [Green Version]
- Thiet, R.K.; Frey, S.D.; Six, J. Do growth yield efficiencies differ between soil microbial communities differing in fungal:bacterial ratios? Reality check and methodological issues. Soil Biol. Biochem. 2006, 38, 837–844. [Google Scholar] [CrossRef]
- Huhe; Chen, X.; Hou, F.; Wu, Y.; Cheng, Y. Bacterial and fungal community structures in Loess Plateau Grasslands with different grazing intensities. Front. Microbiol. 2017, 8, 606. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, S.K.; Nemergut, D.R.; Darcy, J.L.; Lynch, R. Do bacterial and fungal communities assemble differently during primary succession? Mol. Ecol. 2014, 23, 254–258. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | IR | M | CS | CK | F Value | p Value |
|---|---|---|---|---|---|---|
| TN (g/kg) | 0.54 ± 0.02 ab | 0.64 ± 0.11 a | 0.65 ± 0.03 a | 0.49 ± 0.04 b | 4.69 | 0.36 |
| TP (g/kg) | 0.40 ± 0.10 a | 0.44 ± 0.04 a | 0.46 ± 0.01 a | 0.37 ± 0.01 a | 1.86 | 0.21 |
| TK (g/kg) | 15.96 ± 0.29 b | 16.55 ± 0.17 ab | 17.30 ± 0.83 a | 16.64 ± 0.03 ab | 4.51 | 0.04 |
| Available nitrogen (mg/kg) | 8.67 ± 1.43 c | 13.48 ± 1.96 c | 35.88 ± 4.63 a | 24.88 ± 1.62 b | 59.37 | <0.01 |
| Available phosphorus(mg/kg) | 1.57 ± 0.28 d | 5.75 ± 0.79 c | 18.71 ± 1.72 b | 31.13 ± 1.65 a | 336.98 | <0.01 |
| Available potassium(mg/kg) | 68.59 ± 0.09 c | 89.14 ± 4.08 b | 88.49 ± 0.09 b | 104.29 ± 0.15 a | 154.12 | <0.01 |
| Ammonium nitrogen (mg/kg) | 2.96 ± 0.29 a | 4.68 ± 1.86 a | 3.23 ± 0.36 a | 3.88 ± 0.23 a | 1.89 | 0.21 |
| nitrate Nitrogen (mg/kg) | 1.77 ± 0.41 b | 6.99 ± 1.06 a | 5.73 ± 0.95 a | 6.73 ± 0.82 a | 24.55 | <0.01 |
| pH | 8.45 ± 0.07 ab | 8.69 ± 0.27 a | 8.51 ± 0.16 ab | 8.22 ± 0.03 b | 4.33 | 0.04 |
| SOC (g/kg) | 2.29 ± 0.13 c | 3.20 ± 0.12 b | 3.86 ± 0.24 a | 1.83 ± 0.12 d | 99.97 | <0.01 |
| MBC (mg/kg) | 86.82 ± 10.50 a | 113.30 ± 17.91 a | 114.23 ± 15.37 a | 94.55 ± 8.96 a | 3.01 | 0.09 |
| DOC (mg/kg) | 25.30 ± 1.50 c | 55.56 ± 2.92 a | 48.02 ± 2.64 b | 18.28 ± 1.00 d | 203.56 | <0.01 |
| Bulk density (g/cm3) | 1.07 ± 0.02 b | 1.11 ± 0.02 b | 1.07 ± 0.04 b | 1.27 ± 0.01 a | 39.55 | <0.01 |
| Enzyme Activities | IR | M | CS | CK | F Value | p Value |
|---|---|---|---|---|---|---|
| Urease (NH4+-N mg/g·24 h) | 0.22 ± 0.17 b | 0.62 ± 0.26 a | 0.40 ± 0.03 ab | 0.57 ± 0.13 a | 3.42 | 0.07 |
| Alkaline phosphatase (U/g) | 763.13 ± 51.63 bc | 920.85 ± 80.71 b | 1555.63 ± 365.29 a | 522.57 ± 101.85 c | 15.30 | <0.01 |
| Sucrase (glucose mg/g·24 h) | 5.48 ± 1.51 ab | 9.98 ± 7.00 ab | 12.57 ± 5.30 a | 2.82 ± 0.10 b | 2.90 | 0.10 |
| Amylase (U/g) | 1.73 ± 0.11 b | 1.68 ± 0.05 b | 1.96 ± 0.13 a | 1.41 ± 0.05 c | 18.38 | <0.01 |
| Ligninase (U/g) | 8.33 ± 4.22 a | 10.30 ± 3.82 a | 5.52 ± 2.72 a | 3.61 ± 2.52 a | 2.28 | 0.16 |
| Cellulase (U/g) | 14.25 ± 3.33 a | 11.82 ± 0.82 a | 16.42 ± 5.41 a | 14.99 ± 3.72 a | 0.81 | 0.52 |
| β-Glucosidase (U/g) | 2.26 ± 1.55 ab | 3.20 ± 0.67 a | 1.56 ± 0.88 ab | 0.96 ± 0.62 b | 2.77 | 0.11 |
| Soil Properties | Bacteria | Fungi | ||||||
|---|---|---|---|---|---|---|---|---|
| Chao 1 | Observed Species | Shannon | Simpson | Chao 1 | Observed Species | Shannon | Simpson | |
| Bulk density | −0.71 * | 0.70 * | 0.65 * | 0.47 | 0.29 | 0.28 | −0.05 | 0.10 |
| TN | −0.49 | −0.46 | −0.19 | −0.12 | 0.00 | 0.02 | 0.25 | 0.20 |
| TK | 0.08 | 0.11 | 0.28 | 0.20 | −0.08 | −0.08 | 0.14 | 0.39 |
| Available nitrogen | −0.07 | −0.05 | −0.17 | −0.44 | −0.15 | −0.16 | 0.17 | 0.43 |
| Available phosphorus | 0.43 | 0.44 | 0.35 | 0.06 | 0.05 | 0.04 | 0.03 | 0.30 |
| Available potassium | 0.48 | 0.49 | 0.47 | 0.16 | 0.29 | 0.28 | 0.15 | 0.34 |
| pH | −0.39 | −0.38 | −0.37 | −0.26 | −0.23 | −0.23 | −0.27 | −0.44 |
| SOC | −0.44 | −0.42 | −0.37 | −0.40 | −0.22 | −0.21 | 0.15 | 0.19 |
| Urease | 0.49 | 0.49 | 0.33 | 0.16 | −0.06 | −0.07 | −0.35 | −0.24 |
| Alkaline phosphatase | −0.61 * | −0.59 * | −0.54 | −0.53 | −0.34 | −0.33 | 0.13 | 0.21 |
| Sucrase | −0.45 | −0.41 | −0.08 | 0.04 | −0.02 | −0.01 | 0.27 | 0.29 |
| Amylase | −0.46 | −0.45 | −0.47 | −0.43 | −0.33 | −0.33 | 0.08 | 0.08 |
| Soil Properties | Bacteria | Fungi | ||||||
|---|---|---|---|---|---|---|---|---|
| Proteobacteria | Actinobacteria | Acidobacteria | Chloroflexi | Ascomycota | Basidiomycota | Mortierellomycota | Mucoromycota | |
| Bulk density | −0.48 | 0.30 | 0.61 * | 0.31 | −0.08 | −0.20 | 0.30 | −0.27 |
| TN | 0.44 | −0.58 * | 0.15 | −0.17 | 0.19 | −0.05 | −0.05 | 0.03 |
| TK | 0.10 | −0.45 | 0.65 * | 0.02 | 0.37 | −0.09 | 0.33 | −0.15 |
| Available nitrogen | 0.43 | −0.60 * | 0.33 | −0.49 | 0.31 | −0.15 | 0.47 | −0.09 |
| Available phosphorus | −0.09 | −0.13 | 0.61 * | −0.13 | 0.09 | −0.24 | 0.49 | −0.30 |
| Available potassium | −0.06 | −0.25 | 0.85 ** | 0.10 | 0.12 | −0.37 | 0.48 | −0.35 |
| pH | 0.43 | −0.36 | −0.29 | −0.07 | 0.51 | −0.09 | −0.24 | 0.28 |
| SOC | 0.60 * | −0.71 ** | 0.03 | −0.24 | 0.43 | 0.02 | 0.05 | 0.00 |
| Urease | 0.02 | −0.17 | 0.38 | 0.23 | 0.58 * | −0.16 | 0.04 | −0.30 |
| Alkaline phosphatase | 0.70 * | −0.75 ** | −0.09 | −0.49 | 0.28 | 0.10 | 0.11 | 0.07 |
| Sucrase | 0.35 | −0.55 | 0.25 | −0.14 | 0.12 | 0.05 | 0.02 | −0.04 |
| Amylase | 0.44 | −0.38 | −0.40 | −0.35 | 0.31 | 0.20 | −0.15 | −0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, T.; Yang, F.; Li, R.; Li, Y.; Xu, G.; Zhang, L. The Impact of Viticulture on Soil Characteristics and Microbial Communities in the Ningxia Region of Northwest China. Horticulturae 2022, 8, 1097. https://doi.org/10.3390/horticulturae8121097
Xue T, Yang F, Li R, Li Y, Xu G, Zhang L. The Impact of Viticulture on Soil Characteristics and Microbial Communities in the Ningxia Region of Northwest China. Horticulturae. 2022; 8(12):1097. https://doi.org/10.3390/horticulturae8121097
Chicago/Turabian StyleXue, Tingting, Fan Yang, Ruyi Li, Yue Li, Guoqian Xu, and Liang Zhang. 2022. "The Impact of Viticulture on Soil Characteristics and Microbial Communities in the Ningxia Region of Northwest China" Horticulturae 8, no. 12: 1097. https://doi.org/10.3390/horticulturae8121097
APA StyleXue, T., Yang, F., Li, R., Li, Y., Xu, G., & Zhang, L. (2022). The Impact of Viticulture on Soil Characteristics and Microbial Communities in the Ningxia Region of Northwest China. Horticulturae, 8(12), 1097. https://doi.org/10.3390/horticulturae8121097