
Zinc Oxide Nanoparticles and Fe-Modified Activated Carbon Affecting the In Vitro Growth of Date Palm Plantlets cv. Medjool

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Preparation of Activated Carbon
2.2.2. Preparation of Fe-Modified Activated Carbon (Fe-AC)
2.2.3. Preparation of Zinc Oxide Nanoparticles (ZnO-NPs)
2.3. Characterization of Fe-Modified Activated Carbon and Zinc Oxide Nanoparticles
2.4. Plant Material and Growth Conditions
2.5. Morphological Characteristics
2.6. Physiological and Biochemical Analyses
2.6.1. Assessment of Indole Acetic Acid
2.6.2. Assessment of Photosynthetic Pigments
2.6.3. Assessment of Total Soluble Phenolic Compounds
2.6.4. Assessment of Free Amino Acids
2.6.5. Assessment of Total Soluble Sugars
2.6.6. Assessment of Soluble Protein
2.6.7. Assay of Peroxidase Activity (POD)
2.6.8. Assay of Phenylalanine Ammonia Lyase Activity (PAL)
2.7. Statistical Analysis
3. Results
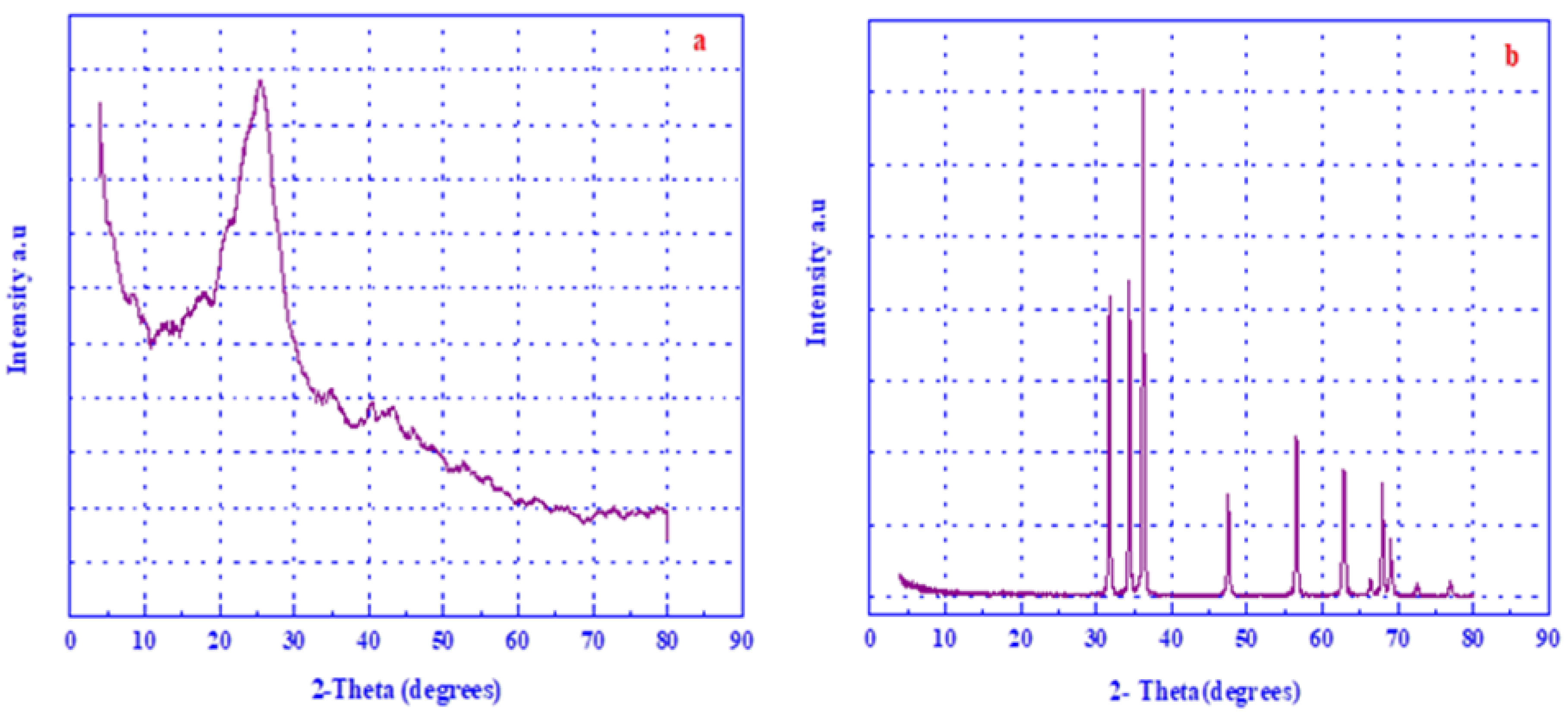
3.1. X-ray Diffraction (XRD)
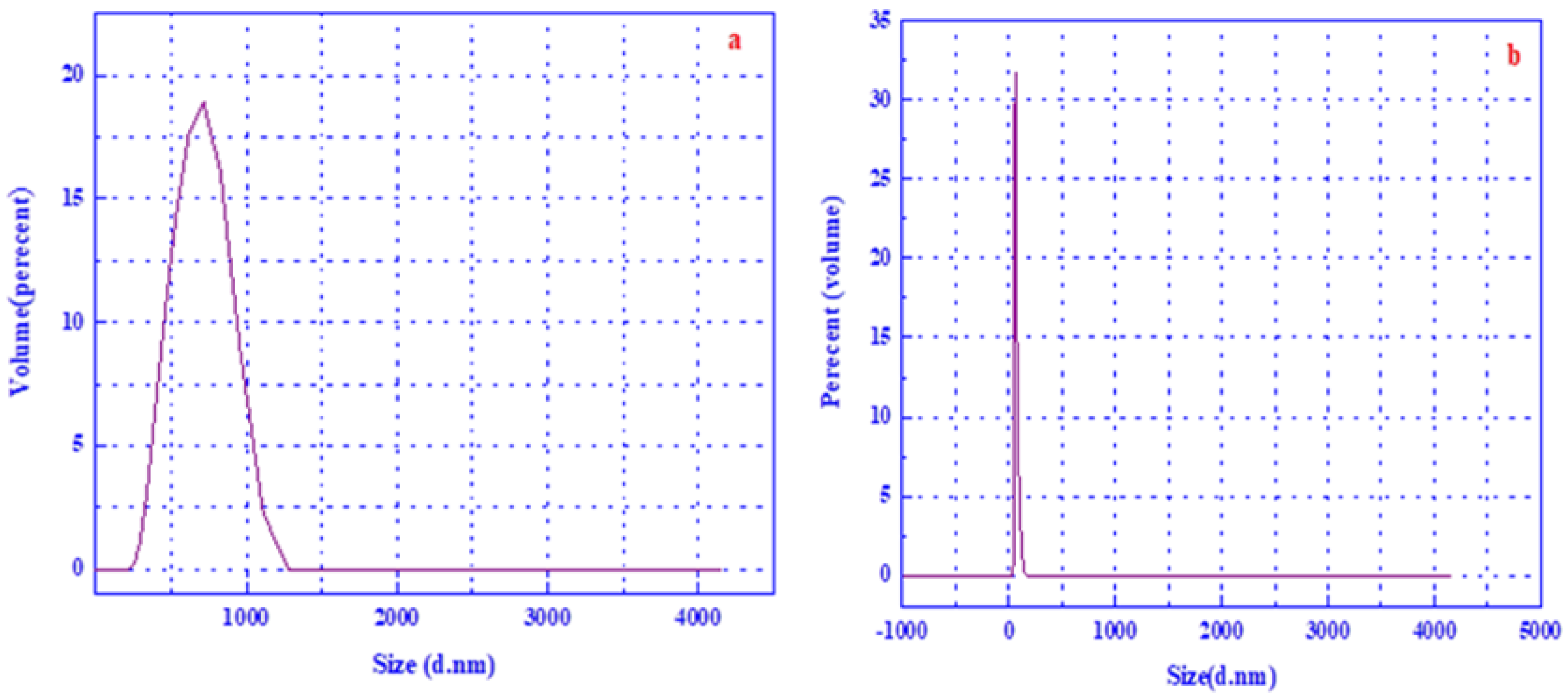
3.2. Dynamic Light Scattering (DLS)
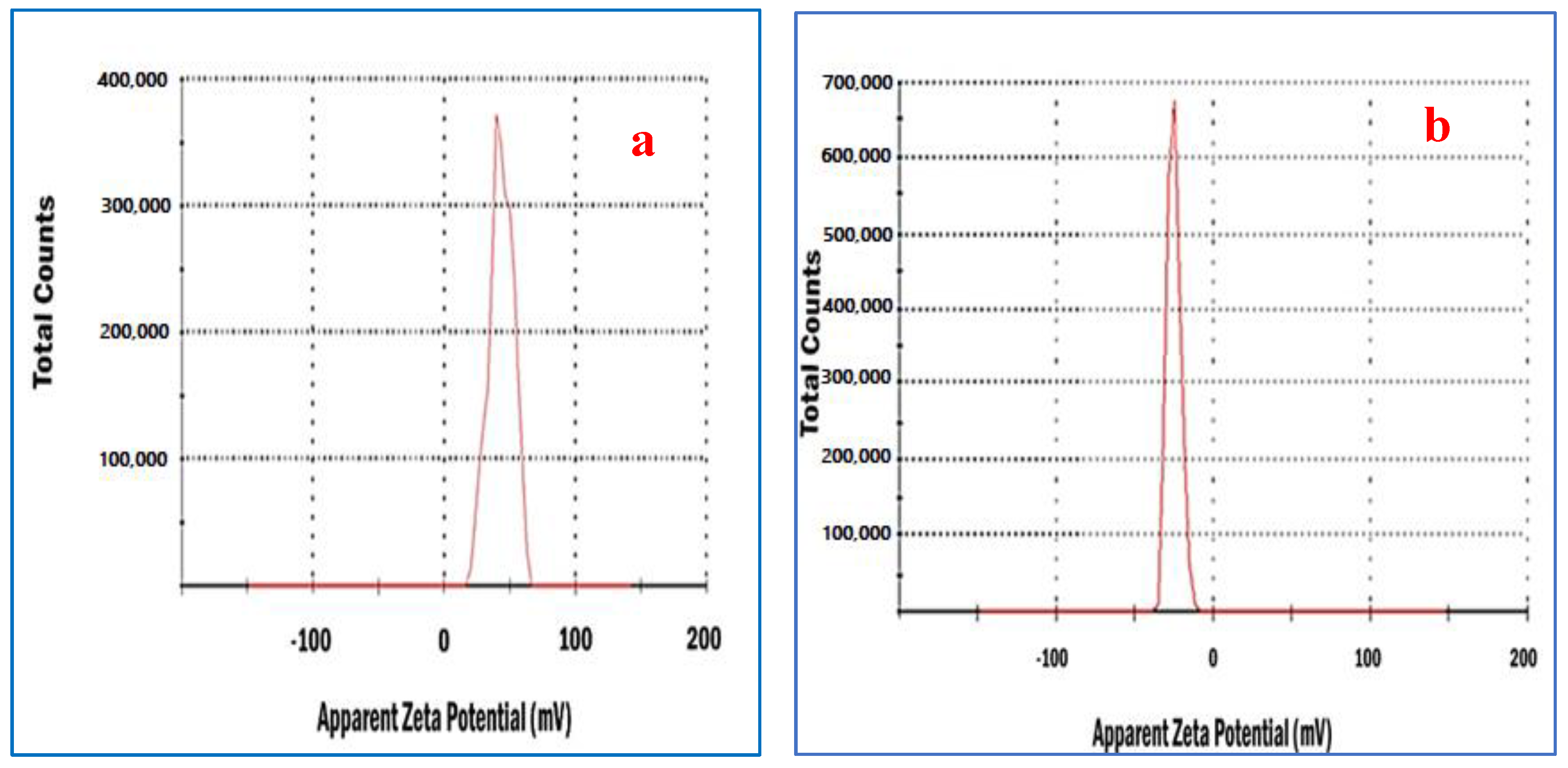
3.3. Zeta Potential Analysis
3.4. Scanning and Transmission Electron Microscopic Images
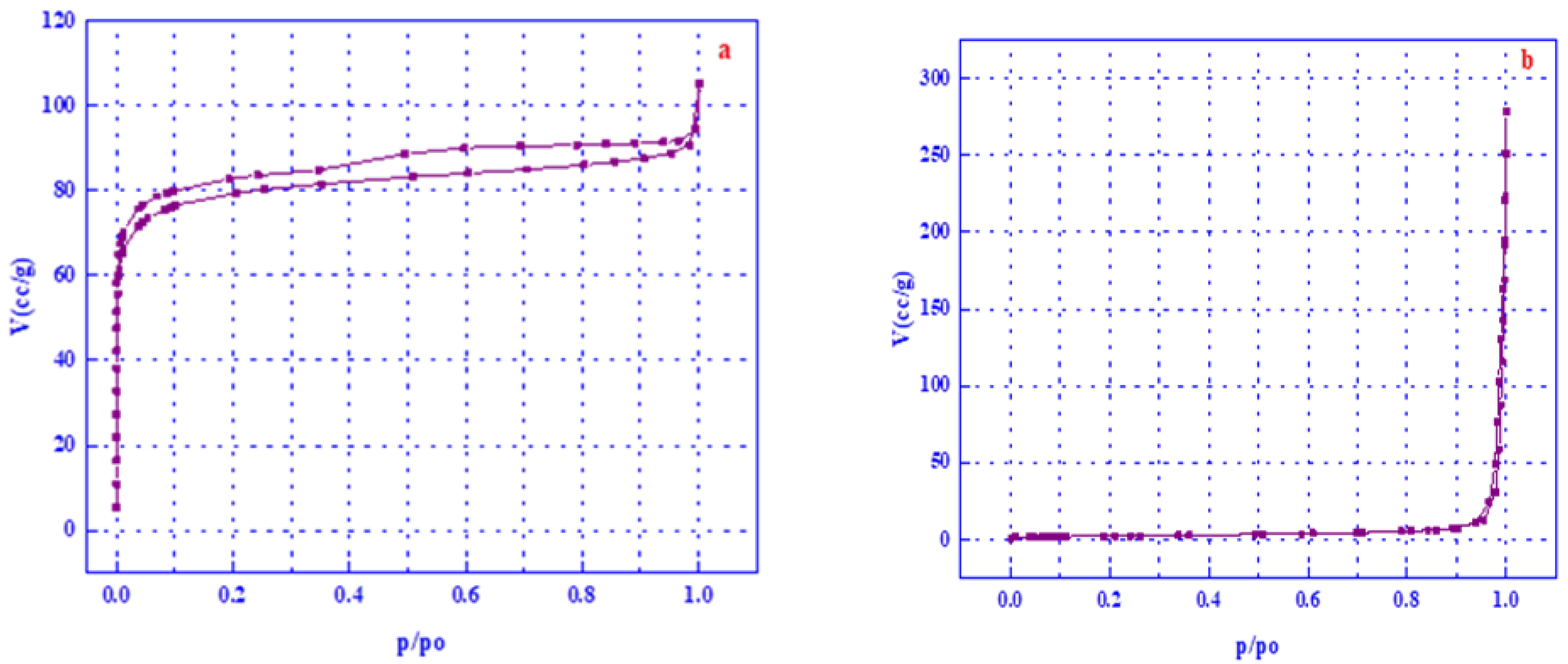
3.5. Surface Area
3.6. Morphological Observations
3.7. Physiological Observations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kholif, A.E.; Gouda, G.A.; Patra, A.K. The sustainable mitigation of in vitro ruminal biogas emissions by ensiling date palm leaves and rice straw with lactic acid bacteria and Pleurotus ostreatus for cleaner livestock production. J. Appl. Microbiol. 2022, 132, 2925–2939. [Google Scholar] [CrossRef] [PubMed]
- Qadir, A.; Shakeel, F.; Ali, A.; Faiyazuddin, M. Phototherapeutic potential and pharmaceutical impact of Phoenix dactylifera (date palm): Current research and future prospects. J. Food Sci. Technol. 2020, 57, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Mirza, M.B.; Elkady, A.; Al-Attar, A.M.; Syed, F.Q.; Mohammed, F.A.; Hakeem, K.R. Induction of apoptosis and cell cycle arrest by ethyl acetate fraction of Phoenix dactylifera L. (Ajwa dates) in prostate cancer cells. J. Ethnopharmacol. 2018, 218, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Kordrostami, M.; Mafakheri, M.; Al-Khayri, J.M. Date palm (Phoenix dactylifera L.) genetic improvement via biotechnological approaches. Tree Genet. Genomes 2022, 18, 1–28. [Google Scholar] [CrossRef]
- Awad, M.A. Promotive effects of a 5-aminolevulinic acid-based fertilizer on growth of tissue culture-derived date palm plants (Phoenix dactylifera L.) during acclimatization. Sci. Hortic. 2008, 118, 48–52. [Google Scholar] [CrossRef]
- Zein El Din, A.F.M.; Ibrahim, M.F.M.; Farag, R.; El-Gawad, H.; El-Banhawy, A.; Alaraidh, I.A.; Rashad, Y.M.; Lashin, I.; El-Yazied, A.A.; Elkelish, A.; et al. Influence of polyethylene glycol on leaf anatomy, stomatal behavior, water loss, and some physiological traits of date palm plantlets grown in vitro and ex vitro. Plants 2020, 9, 1440. [Google Scholar] [CrossRef]
- Kasthuri, J.; Veerapandian, S.; Rajendiran, N. Biological synthesis of silver and gold nanoparticles using apiin as reducing agent. Colloids Surf. B 2009, 68, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Andresen, E.; Peiter, E.; Küpper, H. Trace metal metabolism in plants. J. Exp. Bot. 2018, 69, 909–954. [Google Scholar] [CrossRef] [Green Version]
- Sturikova, H.; Krystofova, O.; Huska, D.; Adam, V. Zinc, zinc nanoparticles and plants. J. Hazard. Mater. 2018, 349, 101–110. [Google Scholar] [CrossRef]
- Kawachi, M.; Kobae, Y.; Mori, H.; Tomioka, R.; Lee, Y.; Maeshima, M. A mutant strain Arabidopsis thaliana that lacks vacuolar membrane zinc transporter MTP1 revealed the latent tolerance to excessive zinc. Plant Cell Physiol. 2009, 50, 1156–1170. [Google Scholar] [CrossRef]
- Lee, S.; Kim, S.A.; Lee, J.; Guerinot, M.L.; An, G. Zinc deficiency inducible osZIP8 encodes a plasma membrane-localized zinc transporter in rice. Mol. Cells 2010, 29, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, T.; Kundu, S.; Rao, A.S. Zinc delivery to plants through seed coating with nano-zinc oxide particles. J. Plant Nutr. 2016, 39, 136–146. [Google Scholar] [CrossRef]
- Rajput, V.; Minkina, T.; Sushkova, S.; Behal, A.; Maksimov, A.; Blicharska, E.; Ghazaryan, K.; Movsesyan, H.; Barsova, N. ZnO and CuO nanoparticles: A threat to soil organisms. Environ. Geochem. Health 2020, 42, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Al-Mayahi, A.M.W. The effect of humic acid (HA) and zinc oxide nanoparticles (ZnO-NPS) on in vitro regeneration of date palm (Phoenix dactylifera L.) cv. Quntar. Plant Cell Tissue Organ Cult. 2021, 145, 445–456. [Google Scholar] [CrossRef]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification. Plant Soil 2008, 30, 1–17. [Google Scholar] [CrossRef]
- Hansch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Hausman, J.F. Changes in peroxidase activity, auxin level and ethylene production during root formation by poplar shoots raised in vitro. Plant Growth Regul. 1993, 13, 263–268. [Google Scholar] [CrossRef]
- Moncousin, C. Adventitious rhizogenesis control: New developments. Acta Hortic. 1988, 230, 97–104. [Google Scholar] [CrossRef]
- Gendreau, E.; Traas, J.; Desnos, T.; Grandjean, O.; Caboche, M.; Hofte, H. Cellular basis of hypocotyl growth in Arabidopsis thaliana. Plant Physiol. 1997, 114, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Ripetti, V.; Kevers, C.; Gaspar, T. Two successive media for the rooting of walnut shoots in vitro. Changes in peroxidase activity and in ethylene production. Adv. Hortic. Sci. 1994, 8, 29–32. [Google Scholar]
- Siegel, B. Plant peroxidases—An organismic perspective. Plant Growth Regul. 1993, 12, 303–312. [Google Scholar] [CrossRef]
- Quoirin, M.; Boxus, P.; Gaspar, T. Root initiation and isoperoxidases of stem tip cuttings from mature Prunus plants. Phys. Veg. 1974, 12, 165–174. [Google Scholar]
- De Klerk, G.J.; Ter Brugge, J.; Smulders, R.; Benschop, M. Basic peroxidases and rooting in microcuttings of Malus domestica. Acta Hortic 1990, 280, 29–36. [Google Scholar] [CrossRef]
- Youssef, A.M.; El-Didamony, H.; El-Sharabasy, S.; Sobhy, M.; Hassan, A.F.; Buláneke, R. Adsorption of 2, 4 dichlorophenoxyacetic acid on different types of activated carbons-based date palm pits: Kinetic and thermodynamic studies. Int. J. Pure Appl. Chem. 2017, 14, 1–15. [Google Scholar] [CrossRef]
- Marrakchi, F.; Ahmed, M.J.; Khanday, W.A.; Asif, M.; Hameed, B.H. Mesoporous activated carbon prepared from chitosan flakes via single-step sodium hydroxide activation for the adsorption of methylene blue. Int. J. Biol. Macromol. 2017, 98, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Dong, F.Q.; Dai, Q.W.; Zhong, G.Q.; Halik, U.; Lee, D.J. Coal tar residues based activated carbon: Preparation and characterization. J. Taiwan Inst. Chem. Eng. 2016, 63, 166–169. [Google Scholar] [CrossRef]
- Liu, Q.S.; Zheng, T.; Li, N.; Wang, P.; Abulikemu, G. Modification of bamboo-based activated carbon using microwave radiation and its effects on the adsorption of methylene blue. Appl. Surf. Sci. 2016, 256, 3309–3315. [Google Scholar] [CrossRef]
- Thomas, T.D. The role of activated charcoal in plant tissue culture. Biotechnol. Adv. 2008, 26, 618–631. [Google Scholar] [CrossRef]
- Bedia, J.; Peñas-Garzón, M.; Gómez-Avilés, A.; Rodriguez, J.J.; Belver, C. Review on activated carbons by chemical activation with FeCl3. J. Carbon Res. 2020, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Pottino, B.G. Methods in Plant Tissue Culture; Deptartment of Horticulture, Agriculture College, Maryland University: College park, MD, USA, 1962; pp. 8–29. [Google Scholar]
- Shindy, W.W.; Smith, O.E. Identification of plant hormones from cotton ovules. Plant Physiol. 1975, 55, 550–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumanta, N.; Haque, C.I.; Nishika, J.; Suprakash, R. Spectrophotometric Analysis of Chlorophylls and Carotenoids from Commonly Grown Fern Species by Using Various Extracting Solvents. Res. J. Chem. Sci. 2014, 4, 63–69. [Google Scholar]
- Shahidi, F.; Naczk, M. Methods of analysis and quantification of phenolic compounds. In Food Phenolic: Sources, Chemistry, Effects and Applications; Technomic Publishind Company, Inc.: Lancaster, PA, USA, 1995; pp. 287–293. [Google Scholar]
- Jayeraman, J. Laboratory Manual in Biochemistry; Wiley Eastern Ltd.: New Delhi, India, 1985; p. 107. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principal of protein—Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Hammer Schmidt, R.; Nuckles, E.M.; Kuc, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant. 1982, 20, 73–82. [Google Scholar]
- Lister, C.E.; Lancaster, J.E.; Walker, J.R.L. Phenylalanine ammonia-lyase activity and its relationship to anthocyanin and flavonoid levels in New Zealand grown apple cultivars. J. Am. Soc. Hortic. Sci. 1996, 121, 281–285. [Google Scholar] [CrossRef]
- SAS. SAS/STAT User’s Guide, Release 6.03 ed.; SAS Institute Inc.: Cary, NC, USA, 1988. [Google Scholar]
- Xu, Z.; Zhang, T.; Yuan, Z.; Zhang, D.; Sun, Z.; Huang, Y.; Zhou, Y. Fabrication of cotton textile waste-based magnetic activated carbon using FeCl 3 activation by the Box–Behnken design: Optimization and characteristics. RSC Adv. 2018, 8, 38081–38090. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Y.; Xu, W.; Wang, Y.; Zhang, B.; Luo, S.; Hu, C. Porous Fe2O3 nanospheres anchored on activated carbon cloth for high-performance symmetric supercapacitors. Nano Energy 2019, 57, 379–387. [Google Scholar] [CrossRef]
- Aroua, M.K.; Leong, S.P.P.; Teo, L.Y.; Yin, C.Y.; Daud, W.M.A.W. Real-time determination of kinetics of adsorption of lead (II) onto palm shell-based activated carbon using ion selective electrode. Bioresour. Technol. 2008, 99, 5786–5792. [Google Scholar] [CrossRef]
- Cheng, S.; Zhang, L.; Ma, A.; Xia, H.; Peng, J.; Li, C.; Shu, J. Comparison of activated carbon and iron/cerium modified activated carbon to remove methylene blue from wastewater. J. Environ. Sci. 2018, 65, 92–102. [Google Scholar] [CrossRef]
- Bigdeli, F.; Morsali, A.; Retalleau, P. Synthesis and Characterization of Different zinc (II) Oxide Nano-Structures from Direct Thermal Decomposition of ID Coordination Polymers. Polyhedron 2010, 29, 801–806. [Google Scholar] [CrossRef]
- Siddique, A.; Nayak, A.K.; Singh, J. Synthesis of FeCl3-activated carbon derived from waste Citrus limetta peels for removal of fluoride: An eco-friendly approach for the treatment of groundwater and bio-waste collectively. Groundw. Sustain. Dev. 2020, 10, 100339. [Google Scholar] [CrossRef]
- Meléndrez, M.F.; Cardenas, G.; Arbiol, J. Synthesis and Characterization of Gallium Colloidal Nanoparticles. J. Colloid Interf. Sci. 2010, 346, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.; Cuesta, A.; Montes-Moran, M.; Martinez-Alonso, A.; Tascon, J. Zeta Potential as a Tool to Characterize Plasma Oxidation of Carbon Fibers. J. Colloid Interface Sci. 1997, 192, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, M.S.; Ahmed, I.A.; Bader, D.M.; Hassan, A.F. Green Synthesis of Nano Zinc Oxide/Nanohydroxyapatite Composites Using Date Palm Pits Extract and Eggshells: Adsorption and Photocatalytic Degradation of Methylene Blue. Nanomaterials 2021, 12, 49. [Google Scholar] [CrossRef]
- Hassan, A.F. Synthesis of carbon nano-onion embedded metal–organic frameworks as an efficient adsorbent for cadmium ions: Kinetic and thermodynamic studies. Environ. Sci. Pollut. Res. 2019, 26, 24099–24111. [Google Scholar] [CrossRef]
- Bulánek, R.; Hrdina, R.; Hassan, A.F. Preparation of polyvinylpyrrolidone modified nano magnetite for degradation of nicotine by heterogeneous Fenton process. J. Environ. Chem. Eng. 2019, 7, 102988–102996. [Google Scholar] [CrossRef]
- Madhusudhanan, K.; Rahiman, B.A. The effect of activated charcoal supplemented media to browning of in vitro cultures of Piper species. Biol. Plant. 2000, 43, 297–299. [Google Scholar] [CrossRef]
- Sparjanbabu, D.S.; Naveen Kumar, P.; Krishna, M.S.R.; Ramajayam, D.; Susanthi, B. Effect Of Activated Charcoal, Culture Media And Plant Growth Regulators on in Vitro Germination and Development of Elite Dura Oil Palm (Elaeis guineensis Jacq.) Zygotic Embryos. Plant Cell Biotechnol. Mol. Biol. 2019, 20, 314–323. [Google Scholar]
- Cattelan, L.V.; Stein, V.C.; Souza, S.A.; Heiden, G.; Buttow, M.V.; Bobrowski, V.L. Estabelecimento in vitro de Matricaria recutita utilizando diferentes condicoes de cultivo. Rev. Bras. Biociências 2007, 5, 201–203. [Google Scholar]
- He Guo, X.; Lu, R.; Niu, B.; Pasapula, V.; Hou, P.; Cai, F.; Xu, Y.; Chen, F. Changes in morphology and biochemical indices in browning callus derived from Jatropha curcus hypocotyls. Plant Cell Tissue Organ Cult. 2009, 98, 11–17. Available online: https://link.springer.com/article/10.1007/s11240-009-9533-y (accessed on 5 July 2022).
- Hassan, M.M.; Taha, R.A.; Abd El-Aziz, M.E.; Shaaban, E.A.; Ibrahim, E.A. Impact of nano-zinc-oxide as an alternative source of zinc in date palm culture media. Plant Cell Tissue Organ Cult. 2022, 150, 73–84. [Google Scholar] [CrossRef]
- Regni, L.; Del Buono, D.; Micheli, M.; Facchin, S.L.; Tolisano, C.; Proietti, P. Effects of Biogenic ZnO Nanoparticles on Growth, Physiological, Biochemical Traits and Antioxidants on Olive Tree In Vitro. Horticulturae 2022, 8, 161. [Google Scholar] [CrossRef]
- El-Mahdy, M.T.; Elazab, D.S. Impact of Zinc Oxide Nanoparticles on Pomegranate Growth under In Vitro Conditions. Russ. J. Plant Physiol. 2020, 67, 162–167. [Google Scholar] [CrossRef]
- Du, W.; Sun, Y.; Ji, R.; Zhu, J.; Wu, J.; Guo, H. TiO2 and ZnO nanoparticles negatively affect wheat growth and soil enzyme activities in agricultural soil. J. Environ. Monitor. 2011, 13, 822–828. [Google Scholar] [CrossRef]
- Pandurangan, M.; Kim, D.H. In vitro toxicity of zinc oxide nanoparticles: A review. J. Nanoparticle Res. 2015, 17, 158. [Google Scholar] [CrossRef]
- Pan, M.; Van Staden, J. Effect of activated charcoal, autoclaving and culture media on sucrose hydrolysis. Plant Growth Regul. 1999, 29, 135–141. [Google Scholar] [CrossRef]
- Liu, J.H.; Mukherjee, I.; Reid, D.M. Stimulation of adventitious rooting in sunflower (Helianthus annuus) by low pH: Possible role of auxin. Can. J. Bot. 1993, 71, 1645–1650. [Google Scholar] [CrossRef]
- Norton, M.E.; Boe, A.A. In vitro propagation of ornamental Rosaceous plant. HortScience 1982, 17, 190–191. [Google Scholar] [CrossRef]
- Spiegel-Roy, P.; Goldschmidt, E. Biology of Citrus; Cambridge University Press: Cambridge, UK; pp. 140–184.
- Regni, L.; Micheli, M.; Del Pino, A.M.; Palmerini, C.A.; D’Amato, R.; Facchin, S.L.; Famiani, F.; Peruzzi, A.; Mairech, H.; Proietti, P. The First Evidence of the Beneficial Effects of Se-Supplementation on in Vitro Cultivated Olive Tree Explants. Plants 2021, 10, 1630. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Desoky, E.-S.M.; Saad, A.M.; Eid, R.S.M.; Selem, E.; Elrys, A.S. Biological Silicon Nanoparticles Improve Phaseolus vulgaris L. Yield and Minimize Its Contaminant Contents on a Heavy Metals-Contaminated Saline Soil. J. Environ. Sci. China 2021, 106, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, N.; Ahmed, T.; Noman, M.; Shahid, M.; Nazir, M.M.; Ali, L.; Alnusaire, T.S.; Li, B.; Schulin, R.; Wang, G. Iron Oxide Nanoparticles Ameliorated the Cadmium and Salinity Stresses in Wheat Plants, Facilitating Photosynthetic Pigments and Restricting Cadmium Uptake. Sci. Total Environ. 2021, 769, 145221. [Google Scholar] [CrossRef] [PubMed]
- Del Buono, D.; Di Michele, A.; Costantino, F.; Trevisan, M.; Lucini, L. Biogenic Zno Nanoparticles Synthesized Using a Novel Plant Extract: Application to Enhance Physiological and Biochemical Traits in Maize. Nanomaterials 2021, 11, 1270. [Google Scholar] [CrossRef] [PubMed]
- Salama, D.M.; Osman, S.A.; Abd El-Aziz, M.E.; Abd Elwahed, M.S.A.; Shaaban, E.A. Effect of Zinc Oxide Nanoparticles on the Growth, Genomic DNA, Production and the Quality of Common Dry Bean (Phaseolus vulgaris). Biocatal. Agric. Biotechnol. 2019, 18, 101083. [Google Scholar] [CrossRef]
- Tedeschini, E.; Proietti, P.; Timorato, V.; D’Amato, R.; Nasini, L.; Dei Buono, D.; Businelli, D.; Frenguelli, G. Selenium as Stressor and Antioxidant Affects Pollen Performance in Olea Europaea. Flora-Morphol. Distrib. Funct. Ecol. Plants 2015, 215, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Dobrikova, A.G.; Apostolova, E.L.; Han´c, A.; Yotsova, E.; Borisova, P.; Sperdouli, I.; Adamakis, I.-D.S.; Moustakas, M. Cadmium Toxicity in Salvia sclarea, L.: An Integrative Response of Element Uptake, Oxidative Stress Markers, Leaf Structure and Photosynthesis. Ecotoxicol. Environ. Saf. 2021, 209, 111851. [Google Scholar] [CrossRef]
- Salih, A.M.; Al-Qurainy, F.; Khan, S.; Tarroum, M.; Nadeem, M.; Shaikhaldein, H.O.; Gaafar, A.-R.Z.; Alfarraj, N.S. Biosynthesis of Zinc Oxide Nanoparticles Using Phoenix dactylifera and Their Effect on Biomass and Phytochemical Compounds in Juniperus Procera. Sci. Rep. 2021, 11, 19136. [Google Scholar] [CrossRef]
- Zafar, H.; Abbasi, B.H.; Zia, M. Physiological and Antioxidative Response of Brassica nigra (L.) to ZnO Nanoparticles Grown in Culture Media and Soil. Toxicol. Environ. Chem. 2019, 101, 281–299. [Google Scholar] [CrossRef]
- Javed, R.; Usman, M.; Yücesan, B.; Zia, M.; Gürel, E. Effect of Zinc Oxide (ZnO) Nanoparticles on Physiology and Steviol Glycosides Production in Micropropagated Shoots of Stevia Rebaudiana Bertoni. Plant Physiol. Biochem. 2017, 110, 94–99. [Google Scholar] [CrossRef]
- Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases have more functions than a Swiss army knife. Cell Rep. 2005, 24, 255–265. [Google Scholar] [CrossRef]
- Deng, F.; Aoki, M.; Yogo, Y. Effect of naringenin on the growth and lignin biosynthesis of gramineous plants. Weed Biol. Manag. 2004, 4, 49–55. [Google Scholar] [CrossRef]
- Sohail; Kamran, K.; Kemmerling, B.; Shutaywi, M.; Mashwani, Z.U.R. Nano zinc elicited biochemical characterization, nutrtitional assessment, antioxidant enzymes and fatty acids profiling of rapeseed. PLoS ONE 2020, 15, e0241568. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahab, D.A.; Othman, N.A.R.M.; Hamada, A.M. Zinc Oxide Nanoparticles Induce Changes in the Antioxidant Systems and Macromolecules in the Solanum Nigrum Callus. Egypt. J. Bot. 2020, 60, 503–517. [Google Scholar] [CrossRef]
- El-Badri, A.M.A.; Batool, M.; Mohamed, I.A.A.; Khatab, A.; Sherif, A.; Wang, Z.; Salah, A.; Nishawy, E.; Ayaad, M.; Kuai, J.; et al. Modulation of Salinity Impact on Early Seedling Stage via Nano-Priming Application of Zinc Oxide on Rapeseed (Brassica napus, L.). Plant Physiol. Biochem. 2021, 166, 376–392. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed; Academic Press Pub.: New York, NY, USA, 1995; p. 559. [Google Scholar]
- Gasic, K.; Korban, S.S. Expression of Arabidopsis phytochelatin synthase in Indian mustard (Brassica juncea) plants enhances tolerance for Cd and Zn. Planta 2007, 225, 1277–1285. [Google Scholar] [CrossRef]
- Empadinhas, N.; Costa, M.S.d. Osmoadaptation mechanisms in prokaryotes: Distribution of compatible solutes. Int. Microbiol. 2008, 11, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Magnaval, C.; Noirot, M.; Verdeil, J.; Blattes, A.; Huet, C.; Grosdemange, F.; Buffard-Morel, J. Free amino acid composition of coconut (Cocos nucifera L.) calli under somatic embryogenesis induction conditions. J. Plant Physiol. 1995, 146, 155–161. [Google Scholar] [CrossRef]
- George, E.F.; Hall, M.A.; De Klerk, G.J. Plant Propagation by Tissue Culture, 3rd ed.; Springer: Dordrecht, The Netherlands, 2008; p. 107. [Google Scholar]
- Bhaduri, A.M.; Fulekar, M.H. Antioxidant enzyme responses of plants to heavy metal stress. Rev. Environ. Sci. Biotechnol. 2012, 11, 55–69. [Google Scholar] [CrossRef]
- Koç, E.; İşlek, C.; Büyükkartal, H.N. Comparison of phenylalanine ammonia lyase response to lead and zinc stress in different wheat genotypes. Commun. Fac. Sci. Univ. Ank. Ser. C 2018, 27, 37–44. [Google Scholar]
- Farghaly, F.A.; Radi, A.A.; Al-Kahtany, F.A.; Hamada, A.M. Impacts of zinc oxide nano and bulk particles on redox-enzymes of the Punica granatum callus. Sci. Rep. 2020, 10, 19722. [Google Scholar] [CrossRef]
- Asada, K. Production and action of active oxygen species in photosynthetic tissues. In Causes of Photo-Oxidative Stress and Amelioration of Defense Systems in Plants; Foyer, C., Mullineaux, P., Eds.; CRC Press: Boca Raton, FL, USA, 1994; pp. 77–104. [Google Scholar]
- Rai, V.; Vaypayee, P.; Singh, S.N.; Mehrotra, S. Effect of chromium accumulation on photosynthetic pigments oxidative stress defence system nitrate reduction proline level and eugenol content of Ocimum tenuiflorum L. Plant Sci. 2004, 167, 1159. [Google Scholar] [CrossRef]
- Radi, A.A.; Farghaly, F.A.; Al-Kahtany, F.A.; Hamada, A.M. Zinc oxide nanoparticles-mediated changes in ultrastructure andmacromolecules of pomegranate callus cells. Plant Cell Tissue Organ Cult. 2018, 135, 247–261. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | SBET (m2/g) | CBET | VP (cm3/g) | r (nm) |
|---|---|---|---|---|
| Fe-AC ZnO-NPs | 369.75 8.99 | 2483 102 | 0.1470 0.1467 | 1.9355 64.23 |
| Treatments | Control | ZnO-NPs | ZnO-BPs | Mean | HSD | Control | ZnO-NPs | ZnO-BPs | Mean | HSD |
|---|---|---|---|---|---|---|---|---|---|---|
| Growth vigor | Plantlet fresh weight (g) | |||||||||
| AC | 2.33 b ± 0.58 | 5.00 a ± 0.00 | 5.00 a ± 0.00 | 4.11 A | 0.40 | 0.72 c ± 0.01 | 2.54 b ± 0.02 | 2.50 b ± 0.05 | 1.92 A | 0.14 |
| Fe-AC | 1.33 c ± 0.58 | 5.00 a ± 0.00 | 4.50 a ± 0.50 | 3.61 B | 0.55 c ± 0.02 | 2.41 b ± 0.31 | 2.86 a ± 0.11 | 1.94 A | ||
| Mean | 1.83 B | 5.00 A | 4.75 A | 0.63 C | 2.48 B | 2.68 A | ||||
| HSD | 0.49 | 0.17 | ||||||||
| Plantlet height (cm) | Number of leaves/plantlet | |||||||||
| AC | 27.67 cd ± 1.53 | 55.33 a ± 2.52 | 30.67 c ± 2.08 | 37.89 A | 1.88 | 1.00 b ± 0.00 | 4.00 a ± 0.82 | 4.00 a ± 0.82 | 3.00 A | 0.73 |
| Fe-AC | 25.33 d ± 0.58 | 36.67 b ± 2.08 | 29.67 cd ± 1.53 | 30.56 B | 1.67 b ± 0.47 | 1.67 b ± 0.47 | 3.33 a ± 0.47 | 2.22 B | ||
| Mean | 26.50 C | 46.00 A | 30.17 B | 1.33 B | 2.83 A | 3.67 A | ||||
| HSD | 2.30 | 0.89 | ||||||||
| Leaf length (cm) | Leaf width (cm) | |||||||||
| AC | 20.33 d ± 1.53 | 37.67 a ± 1.53 | 20.67 d ± 3.06 | 26.22 A | 2.57 | 0.49 c ± 0.02 | 1.02 a ± 0.04 | 0.80 b ± 0.01 | 0.77 A | 0.08 |
| Fe-AC | 15.50 e ± 0.50 | 31.00 b ± 4.58 | 25.67 c ± 1.53 | 24.06 A | 0.30 d ± 0.02 | 1.08 a ± 0.16 | 0.82 b ± 0.07 | 0.73 A | ||
| Mean | 17.92 C | 34.33 A | 23.17 B | 0.34 C | 1.05 A | 0.81 B | ||||
| HSD | 3.15 | 0.09 | ||||||||
| Root length (cm) | Number of roots/plantlet | |||||||||
| AC | 5.47 c ± 0.50 | 15.33 a ± 1.53 | 4.63 cd ± 0.65 | 8.48 A | 0.88 | 3.33 d ± 0.58 | 14.67 b ± 1.53 | 18.33 a ± 1.53 | 12.11 A | 1.33 |
| Fe-AC | 7.40 b ± 0.53 | 4.17 cd ± 1.04 | 3.13 d ± 0.15 | 4.90 B | 2.33 d ± 0.58 | 11.33 c ± 1.53 | 16.33 ab ± 1.53 | 10.00 B | ||
| Mean | 6.43 B | 9.75 A | 3.88 C | 2.83 c | 13.00 B | 17.33 A | ||||
| HSD | 1.07 | 1.62 | ||||||||
| Treatments | Control | ZnO-NPs | ZnO-BPs | Mean | HSD | Control | ZnO-NPs | ZnO-BPs | Mean | HSD |
|---|---|---|---|---|---|---|---|---|---|---|
| IAA (mg/g FW) | Soluble Protein (mg/g FW) | |||||||||
| AC | 0.058 c ± 0.012 | 0.079 a ± 0.006 | 0.075 ab ± 0.006 | 0.073 A | 0.008 | 0.04 b ± 0.006 | 0.07 a ± 0.024 | 0.05 ab ± 0.003 | 0.05 A | 0.01 |
| Fe-AC | 0.057 c ± 0.004 | 0.077 a ± 0.003 | 0.064 bc ± 0.011 | 0.064 B | 0.05 ab ± 0.013 | 0.06 ab ± 0.002 | 0.05 ab ± 0.015 | 0.05 A | ||
| Mean | 0.061 B | 0.079 A | 0.067 B | 0.04 A | 0.06 A | 0.05 A | ||||
| HSD | 0.009 | 0.02 | ||||||||
| Free amino acids (mg/g FW) | Total sugars (mg/g FW) | |||||||||
| AC | 5.44 cd ± 1.11 | 6.29 bc ± 0.28 | 6.75 ab ± 0.70 | 6.16 A | 0.65 | 1.50 b ± 0.16 | 1.87 b ± 0.25 | 1.72 b ± 0.11 | 1.69 B | 0.32 |
| Fe-AC | 4.93 d ± 0.45 | 6.19 bc ± 0.28 | 7.70 a ± 0.55 | 6.28 A | 1.42 b ± 0.08 | 1.65 b ± 0.08 | 3.09 a ± 0.69 | 2.05 A | ||
| Mean | 5.19 C | 6.24 B | 7.22 A | 1.46 B | 1.76 B | 2.40 A | ||||
| HSD | 0.79 | 0.3911 | ||||||||
| Chlorophyll a (mg/g FW) | Chlorophyll b (mg/g FW) | |||||||||
| AC | 0.15 d ± 0.003 | 0.32 a ± 0.047 | 0.30 ab ± 0.002 | 0.26 A | 0.02 | 0.12 d ± 0.004 | 0.19 a ± 0.020 | 0.18 ab ± 0.18 | 0.16 A | 0.01 |
| Fe-AC | 0.17c d ± 0.007 | 0.27 b ± 0.008 | 0.20 c ± 0.019 | 0.21 B | 0.13 cd ± 0.012 | 0.16 ab ± 0.008 | 0.15 bc ± 0.15 | 0.15 B | ||
| Mean | 0.16 C | 0.30 A | 0.249 B | 0.13 B | 0.17 A | 0.17 A | ||||
| HSD | 0.03 | 0.02 | ||||||||
| Carotenoids (mg/g FW) | Total soluble phenols (mg/g FW) | |||||||||
| AC | 0.03 b ± 0.009 | 0.04 ab ± 0.006 | 0.04 ab ± 0.004 | 0.04 A | 0.0056 | 1.03 c ± 0.31 | 1.50 ac ± 0.22 | 1.43 bc ± 0.006 | 1.32 B | 0.40 |
| Fe-AC | 0.03 ab ± 0.001 | 0.04 a ± 0.004 | 0.03 ab ± 0.006 | 0.04 A | 1.22 bc ± 0.08 | 1.79 ab ± 0.61 | 2.15 a ± 0.62 | 1.72 A | ||
| Mean | 0.03 B | 0.04 A | 0.04 AB | 1.13 B | 1.65 A | 1.80 A | ||||
| HSD | 0.007 | 0.49 | ||||||||
| POD (unit/mg protein) | PAL (unit/mg protein) | |||||||||
| AC | 767.29 d ± 42.76 | 1172.38 bc ± 75.01 | 1274.93 ac ± 104.9 | 1071.53 B | 115.64 | 353.49 d ± 82.2 | 806.37 b ± 53.51 | 676.87 c ± 38.74 | 612.24 B | 65.09 |
| Fe-AC | 1075.92 c ± 41.5 | 1294.20 ab ± 198.65 | 1465.51 a ± 128.11 | 1278.54 A | 574.88 c ± 12.52 | 833.67 b ± 103.70 | 971.66 a ± 45.60 | 793.39 A | ||
| Mean | 921.60 B | 1233.29 A | 1370.22 A | 464.18 B | 820.02 A | 824.24 A | ||||
| HSD | 141.63 | 79.73 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdellatif, Y.M.R.; Elsayed, M.S.; Hassan, M.M.; Ahmed, I.A.; Ragab, A.H.; Shams El-Din, I.M.; Abdelaal, W.B.; El-Aal, M.S.A.; Zein El Din, A.F.M. Zinc Oxide Nanoparticles and Fe-Modified Activated Carbon Affecting the In Vitro Growth of Date Palm Plantlets cv. Medjool. Horticulturae 2022, 8, 1179. https://doi.org/10.3390/horticulturae8121179
Abdellatif YMR, Elsayed MS, Hassan MM, Ahmed IA, Ragab AH, Shams El-Din IM, Abdelaal WB, El-Aal MSA, Zein El Din AFM. Zinc Oxide Nanoparticles and Fe-Modified Activated Carbon Affecting the In Vitro Growth of Date Palm Plantlets cv. Medjool. Horticulturae. 2022; 8(12):1179. https://doi.org/10.3390/horticulturae8121179
Chicago/Turabian StyleAbdellatif, Yasmin M. R., Maha S. Elsayed, Mona M. Hassan, Inas A. Ahmed, Ahmed H. Ragab, Ibrahim M. Shams El-Din, Walid B. Abdelaal, Mona S. Abd El-Aal, and Amal F. M. Zein El Din. 2022. "Zinc Oxide Nanoparticles and Fe-Modified Activated Carbon Affecting the In Vitro Growth of Date Palm Plantlets cv. Medjool" Horticulturae 8, no. 12: 1179. https://doi.org/10.3390/horticulturae8121179
APA StyleAbdellatif, Y. M. R., Elsayed, M. S., Hassan, M. M., Ahmed, I. A., Ragab, A. H., Shams El-Din, I. M., Abdelaal, W. B., El-Aal, M. S. A., & Zein El Din, A. F. M. (2022). Zinc Oxide Nanoparticles and Fe-Modified Activated Carbon Affecting the In Vitro Growth of Date Palm Plantlets cv. Medjool. Horticulturae, 8(12), 1179. https://doi.org/10.3390/horticulturae8121179