


The Potential Roles of the N and P Supplies on the Internal Browning Incidence in Sweet Cherries in the Southern Chile


 ,
,  and
and
Abstract
:1. Introduction
2. Andisols of Southern Chile: A Particular Scenario for the Sweet Cherry Production
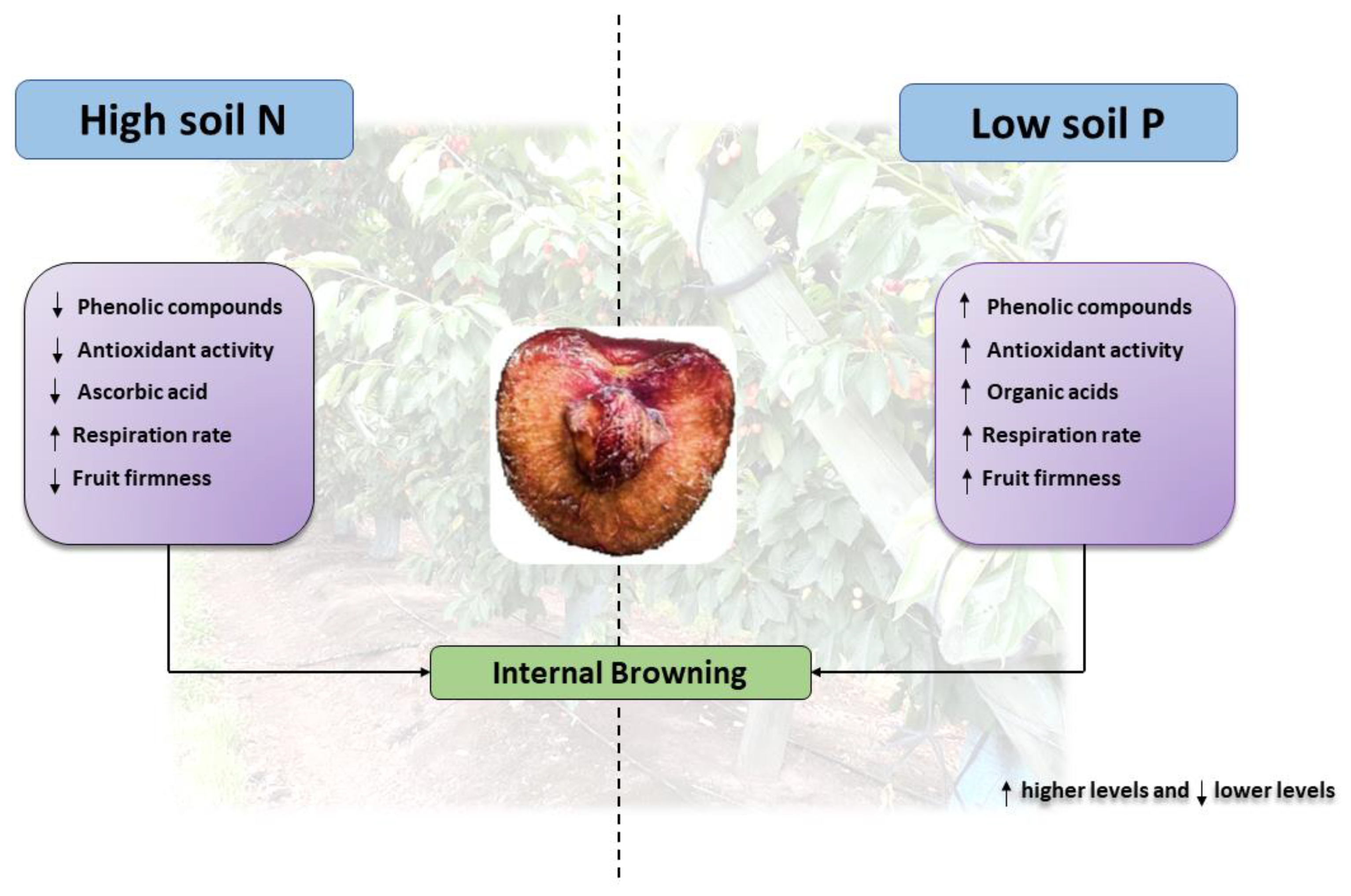
3. Physiological and Biochemical Traits of the Internal Browning in Fresh Fruits
4. Non-Enzymatic Antioxidant Compounds in Sweet Cherry Fruits
5. Nitrogen Excess Could Reduce the Quality, Condition, and Antioxidant Levels in Fruits
6. Soil P Supply: A Key Pre-Harvest Factor to Enhance the Quality, Condition and Antioxidants in Fruits

{kind=link}
{kind=link}
| Species | Treatments | Phosphorus Treatment | Effects | References |
|---|---|---|---|---|
| Prunus cerasus × Prunus canescens | ammonium polyphosphate | 20 g P tree−1 | Increased P concentrations in fruit, but had lower levels of phenolic compounds | [156] |
| Fragaria ananassa | P | 40–60 Kg ha−1 | Higher effect on the fruit length, fruit diameter, and fruits per plants, and fruit weight | [150] |
| Fragaria ananassa | Control-Fe−-P− | P− | Lower °Brix accumulation and higher phenolic compounds synthesis in fruits | |
| Higher malic acid, lower quinic acid, sorbitol-6-phosphate and quinic acid | [153] | |||
| Malus domestica | Macronutrients 1/4-strength of MSmedium (1/4 MS), 1/2-strength (1/2 MS), full-strength (1 MS), two-fold (2 MS) and three-fold (3 MS) | 1/4-x MS and 1/2xMS | Higher concentration of anthocyanins in the shoots | [157] |
| Fragaria ananassa | 0–20–30–40-50% of P | 30% P | Higher anthocyanins concentration in the fruits | [158] |
| Malus domestica | 0–0.5 mg/mL TSP solution | 0.5 mg/TSP | Decreased weight loss, flesh firmness and respiration rate | [160] |
| Prunus persica | Control Full fertilized- Low N- Low P-Low K (hoagland’s solution) | Low P | Decreased yield, fruit weight. Increased SSC, Antioxidant activity, Total Phenols, PPO and browning potential | [42] |
| Cucumis sativus L. | 90–360 mg P plant−1 | 90 mg P plant−1 | Increased fruit respiration rate at postharvest | [149] |
| Bramley’s seedling | Control treatment-KH2PO4 sprays | KH2PO4 Sprays | Decreased core flush in fruits, senescent breakdown and superficial scald | [161] |
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- PRO CHILE Destacado en Prensa: Chile se Convierte en el mayor Proveedor Mundial de 28 Productos Liderados Por Cobre, Cerezas y Salmón. Available online: https://www.prochile.gob.cl/noticias/detalle-noticia/2021/08/12/chile-se-convierte-en-el-mayor-proveedor-mundial-de-28-productos-liderados-por-cobre-cerezas-y-salmón (accessed on 11 October 2022).
- ASOEX Estadísticas de Exportación. Available online: https://www.asoex.cl/estadisticas-de-exportacion.html (accessed on 20 October 2022).
- ODEPA-CIREN Catastro Frutícola Región de O’Higgins. 2021. Available online: https://www.ciren.cl/proyectos/catastros/catastro-fruticola/ (accessed on 25 October 2022).
- Murakami, Y.; Hernández, R. The Impacts of China on Economic Growth: Evidence for Brazil, Chile, and Peru. J. Post Keynes. Econ. 2018, 41, 1–25. [Google Scholar] [CrossRef]
- Blanke, M.; Yuri, A. Chile: Record Exports of Fruit Grown by the Andes. Erwerbsobstbau 2020, 62, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Rojas, G.; Fernandez, E.; Whitney, C.; Luedeling, E.; Cuneo, I.F. Adapting Sweet Cherry Orchards to Extreme Weather Events—Decision Analysis in Support of Farmers’ Investments in Central Chile. Agric. Syst. 2021, 187, 103031. [Google Scholar] [CrossRef]
- Mora, M.L.; Garcia, J.; Santander, J.; Demanet, R. Rol de Los Fertilizantes Nitrogenados y Fosfatados En Los Procesos de Acidificación de Los Suelos. Front. Agrícola 1999, 51, 59–81. [Google Scholar]
- Bañados, M.P.; Strik, B.C.; Bryla, D.R.; Righetti, T.L. Response of Highbush Blueberry to Nitrogen Fertilizer During Field Establishment, I: Accumulation and Allocation of Fertilizer Nitrogen and Biomass. HortScience 2012, 47, 648–655. [Google Scholar] [CrossRef]
- Ribera, A.E.; Reyes-Diaz, M.; Alberdi, M.; Zuñiga, G.E.; Mora, M.L. Antioxidant Compounds in Skin and Pulp of Fruits Change among Genotypes and Maturity Stages in Highbush Blueberry (Vaccinium corymbosum L.) Grown in Southern Chile. J. Soil Sci. Plant Nutr. 2010, 10, 509–536. [Google Scholar] [CrossRef]
- González-Villagra, J.; Escobar, A.L.; Ribera-Fonseca, A.; Cárcamo, M.P.; Omena-Garcia, R.P.; Nunes-Nesi, A.; Inostroza-Blancheteau, C.; Alberdi, M.; Reyes-Díaz, M. Differential Mechanisms between Traditionally Established and New Highbush Blueberry (Vaccinium corymbosum L.) Cultivars Reveal New Insights into Manganese Toxicity Resistance. Plant Physiol. Biochem. 2021, 158, 454–465. [Google Scholar] [CrossRef]
- Mora, M.L.; Canales, J. Humin-clay Interactions on Surface Reactivity in Chilean Andisols. Commun. Soil Sci. Plant Anal. 1995, 26, 2819–2828. [Google Scholar] [CrossRef]
- Huygens, D.; Roobroeck, D.; Cosyn, L.; Salazar, F.; Godoy, R.; Boeckx, P. Microbial Nitrogen Dynamics in South Central Chilean Agricultural and Forest Ecosystems Located on an Andisol. Nutr. Cycl. Agroecosyst. 2011, 89, 175–187. [Google Scholar] [CrossRef]
- Mora, M.L.; Baeza, G.; Pizarro, C.; Demanet, R. Effect of Calcitic and Dolomitic Lime on Physicochemical Properties of a Chilean Andisol. Commun. Soil Sci. Plant Anal. 1999, 30, 427–439. [Google Scholar] [CrossRef]
- Barrow, N.J.; Cartes, P.; Mora, M.L. Modifications to the Freundlich Equation to Describe Anion Sorption over a Large Range and to Describe Competition between Pairs of Ions. Eur. J. Soil. Sci. 2005, 56, 601–606. [Google Scholar] [CrossRef]
- Cartes, P.; Cea, M.; Jara, A.; Violante, A.; de la Luz Mora, M. Description of Mutual Interactions between Silicon and Phosphorus in Andisols by Mathematical and Mechanistic Models. Chemosphere 2015, 131, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Santibáñez, Q.F. Atlas Agroclimático de Chile. Estado Actual y Tendencias Del Clima. Tomo IV: Regiones Del Biobío y de La Araucanía, 1st ed.; Universidad de Chile. Facultad de Ciencias agronómicas, FIA: Santiago, Chile, 2017; p. 74. [Google Scholar]
- Correia, S.; Schouten, R.; Silva, A.P.; Gonçalves, B. Factors Affecting Quality and Health Promoting Compounds during Growth and Postharvest Life of Sweet Cherry (Prunus avium L.). Front. Plant Sci. 2017, 8, 2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balbontín, C.; Ayala, H.; Rubilar, J.; Cote, J.; Figueroa, C.R. Transcriptional Analysis of Cell Wall and Cuticle Related Genes during Fruit Development of Two Sweet Cherry Cultivars with Contrasting Levels of Cracking Tolerance. Chil. J. Agric. Res. 2014, 74, 162–169. [Google Scholar] [CrossRef] [Green Version]
- Roco, L.; Engler, A.; Bravo-Ureta, B.; Jara-Rojas, R. Farm Level Adaptation Decisions to Face Climatic Change and Variability: Evidence from Central Chile. Environ. Sci. Policy 2014, 44, 86–96. [Google Scholar] [CrossRef]
- Blanke, M.M.; Lang, G.A.; Meland, M. Orchard Microclimate Modification. Cherries Bot. Prod. Uses 2017, 244–268. [Google Scholar] [CrossRef]
- Rios, J.C.; Robledo, F.; Schreiber, L.; Zeisler, V.; Lang, E.; Carrasco, B.; Silva, H. Association between the Concentration of N-Alkanes and Tolerance to Cracking in Commercial Varieties of Sweet Cherry Fruits. Sci. Hortic. 2015, 197, 57–65. [Google Scholar] [CrossRef]
- Cline, J.A.; Meland, M.; Sekse, L.; Webster, A.D. Rain Cracking of Sweet Cherries: II. Influence of Rain Covers and Rootstocks on Cracking and Fruit Quality. Acta Agric. Scand. 1995, 45, 224–230. [Google Scholar] [CrossRef]
- Blanco, V.; Zoffoli, J.P.; Ayala, M. High Tunnel Cultivation of Sweet Cherry (Prunus avium L.): Physiological and Production Variables. Sci. Hortic. 2019, 251, 108–117. [Google Scholar] [CrossRef]
- Bustamante, M.; Muñoz, A.; Romero, I.; Osorio, P.; Mánquez, S.; Arriola, R.; Reyes-Díaz, M.; Ribera-Fonseca, A. Impact of Potassium Pre-Harvest Applications on Fruit Quality and Condition of Sweet Cherry (Prunus avium L.) Cultivated under Plastic Covers in Southern Chile Orchards. Plants 2021, 10, 2778. [Google Scholar] [CrossRef]
- Dever, M.; MacDonald, R.A.; Cliff, M.A.; Lane, W.D. Sensory Evaluation of Sweet Cherry Cultivars. HortScience 1996, 31, 150–153. [Google Scholar] [CrossRef]
- Crisosto, C.H.; Crisosto, G.M.; Metheney, P. Consumer Acceptance of ‘Brooks’ and ‘Bing’ Cherries Is Mainly Dependent on Fruit SSC and Visual Skin Color. Postharvest Biol. Technol. 2003, 28, 159–167. [Google Scholar] [CrossRef]
- McCune, L.M.; Kubota, C.; Stendell-Hollis, N.R.; Thomson, C.A. Cherries and Health: A Review. Crit. Rev. Food Sci. Nutr. 2010, 51, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.S.; Adkins, Y.; Laugero, K.D. A Review of the Health Benefits of Cherries. Nutrients 2018, 10, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Cao, G.; Prior, R.L. Total Antioxidant Capacity of Fruits. J. Agric. Food Chem. 1996, 44, 701–705. [Google Scholar] [CrossRef]
- Giné-Bordonaba, J.; Echeverria, G.; Ubach, D.; Aguiló-Aguayo, I.; López, M.L.; Larrigaudière, C. Biochemical and Physiological Changes during Fruit Development and Ripening of Two Sweet Cherry Varieties with Different Levels of Cracking Tolerance. Plant Physiol. Biochem. 2017, 111, 216–225. [Google Scholar] [CrossRef]
- Alique, R.; Martínez, M.A.; Alonso, J. Influence of the Modified Atmosphere Packaging on Shelf Life and Quality of Navalinda Sweet Cherry. Eur. Food Res. Technol. 2003, 217, 416–420. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Zhang, X.; Wang, F.; Zhao, Q. Effect of Pressurized Argon Combined with Controlled Atmosphere on the Postharvest Quality and Browning of Sweet Cherries. Postharvest Biol. Technol. 2019, 147, 59–67. [Google Scholar] [CrossRef]
- Zoffoli, J.P.; Paulina, N.; Jessica, R.; Álvaro, J.; Carolina, C.; Natalia, P. Características de Poscosecha de Cerezas Cultivar “Regina”. Red Agrícola 2020, 42, 37–45. [Google Scholar]
- Lepe, J.P. Boletín de Fruta, Junio 2022; ODEPA|Oficina de Estudios y Políticas Agrarias: Santiago, Chile, 2022.
- Harb, J.; Streif, J.; Saquet, A. Impact of Controlled Atmosphere Storage Conditions on Storability and Consumer Acceptability of Sweet Cherries ‘Regina’. J. Hort. Sci. Biotech. 2003, 78, 574–579. [Google Scholar] [CrossRef]
- Karagiannis, E.; Michailidis, M.; Karamanoli, K.; Lazaridou, A.; Minas, I.S.; Molassiotis, A. Postharvest Responses of Sweet Cherry Fruit and Stem Tissues Revealed by Metabolomic Profiling. Plant Physiol. Biochem. 2018, 127, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Escalona, V.; Battistoni, B.; Valdebenito, W.; Sagredo, K. Efecto del almacenamiento prolongado bajo atmósfera controlada sobre la calidad de cerezas Regina. Red Agrícola 2019, 4, 50–53. [Google Scholar]
- Groth, S.; Budke, C.; Neugart, S.; Ackermann, S.; Kappenstein, F.-S.; Daum, D.; Rohn, S. Influence of a Selenium Biofortification on Antioxidant Properties and Phenolic Compounds of Apples (Malus domestica). Antoxidants 2020, 9, 187. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, R.; Proietti, P.; Onofri, A.; Regni, L.; Esposto, S.; Servili, M.; Businelli, D.; Selvaggini, R. Biofortification (Se): Does It Increase the Content of Phenolic Compounds in Virgin Olive Oil (VOO)? PLoS ONE 2017, 12, e0176580. [Google Scholar] [CrossRef] [Green Version]
- Pezzarossa, B.; Remorini, D.; Gentile, M.L.; Massai, R. Effects of Foliar and Fruit Addition of Sodium Selenate on Selenium Accumulation and Fruit Quality. J. Agric. Food Chem. 2012, 92, 781–786. [Google Scholar] [CrossRef]
- Babalar, M.; Mohebbi, S.; Zamani, Z.; Askari, M.A. Effect of Foliar Application with Sodium Selenate on Selenium Biofortification and Fruit Quality Maintenance of ‘Starking Delicious’ Apple during Storage. J. Sci. Food Agric. 2019, 99, 5149–5156. [Google Scholar] [CrossRef]
- Olivos, A.; Johnson, S.; Xiaoqiong, Q.; Crisosto, C.H. Fruit Phosphorous and Nitrogen Deficiencies Affect ‘Grand Pearl’ Nectarine Flesh Browning. HorstScience 2012, 47, 391–394. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Fuentes, A.D.; Colinas, M.T.L.; Pinedo-Espinoza, J.M. Effect of Fertilization on the Concentration of N, P, K, Ca, Mg, Fe, Mn, Cu, Zn, and Phenylalanine Ammonio-Lyase Activity in Fruit of ‘Zacatecas’- Type Peach (Prunus persica (L.) Batsch). Acta Hortic. 2004, 521–525. [Google Scholar] [CrossRef]
- Mora, M.L.; Cartes, P.; Demanet, R.; Cornforth, I.S. Effects of Lime and Gypsum on Pasture Growth and Composition on an Acid Andisol in Chile, South America. Commun. Soil Sci. Plant Anal. 2002, 33, 2069–2081. [Google Scholar] [CrossRef]
- Mora, M.L.; Alfaro, M.; Williams, P.H.; Stehr, W.; Rolando, D. Effect of Fertilizer Input on Soil Acidification in Relation to Growth and Chemical Composition of a Pasture and Animal Production. J. Plant Nutr. Soil Sci. 2004, 4, 29–40. [Google Scholar]
- Fernández, P.; García, F.; Fernandez, J. Estimación de las extracciones de nutrientes en el cultivo de cerezo en la Región de Murcia. Implicaciones del nitrógeno. Actas de Horticultura 2014, 66, 167–175. [Google Scholar]
- Sepúlveda, G.L. El Cultivo del Cerezo; Instituto de Investigacioes Agropecuarias, INIA La Platina: La Pintana, Chile, 2005.
- Yang, W.Q.; Goulart, B.L. Aluminum and Phosphorus Interactions in Mycorrhizal and Nonmycorrhizal Highbush Blueberry Plantlets. J. Am. Soc. Hortic. Sci. 1997, 122, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Quero-Garcia, J.; Lezzoni, A.; Paulawska, J.; Lang, G. Cherries: Botany, Production and Uses; Emma McCann, CABI: Wallingford, UK, 2017; Volume 4. [Google Scholar]
- Hirzel, J. (Ed.) Diagnóstico Nutricional y Principios de Fertilización en Frutales y Vides, 2nd ed.; Trama Impresores: Chillan, Chile, 2014. [Google Scholar]
- Tomás-Barberán, F.A.; Espín, J.C. Phenolic Compounds and Related Enzymes as Determinants of Quality in Fruits and Vegetables. J. Sci. Food Agric. 2001, 81, 853–876. [Google Scholar] [CrossRef]
- Kader, A.; Chordas, A. Evaluating the Browning Potential of Peaches. J.Calif. Agri. 1984, 38, 14–15. [Google Scholar]
- Robards, K.; Prenzler, P.D.; Tucker, G.; Swatsitang, P.; Glover, W. Phenolic Compounds and Their Role in Oxidative Processes in Fruits. Food Chem. 1999, 66, 401–436. [Google Scholar] [CrossRef]
- Murata, M.; Noda, I.; Homma, S. Enzymatic Browning of Apples on the Market: Relationship between Browning, Polyphenol Content, and Polyphenol Oxidase. Jpn. Soc. food Sci. Technol. 1995, 42, 820–826. [Google Scholar] [CrossRef]
- Abdallah, A.Y.; Gil, M.I.; Biasi, W.; Mitcham, E.J. Inhibition of Superficial Scald in Apples by Wounding: Changes in Lipids and Phenolics. Postharvest Biol. Technol. 1997, 12, 203–212. [Google Scholar] [CrossRef]
- Mayer, A.M.; Harel, E. Polyphenol Oxidases in Plants. Phytochemistry 1979, 18, 193–215. [Google Scholar] [CrossRef]
- Moon, K.M.; Kwon, E.-B.; Lee, B.; Kim, C.Y. Recent Trends in Controlling the Enzymatic Browning of Fruit and Vegetable Products. Molecules 2020, 25, 2754. [Google Scholar] [CrossRef]
- Tian, S.-P.; Jiang, A.-L.; Xu, Y.; Wang, Y.-S. Responses of Physiology and Quality of Sweet Cherry Fruit to Different Atmospheres in Storage. Food Chem. 2004, 87, 43–49. [Google Scholar] [CrossRef]
- Barrelt, D.M.; Gonzalez, C. Activity of Softening Enzymes during Cherry Maturation. J. Food. Sci. 1994, 59, 574–577. [Google Scholar] [CrossRef]
- Gatto, M.A.; Sergio, L.; Ippolito, A.; Di Venere, D. Phenolic Extracts from Wild Edible Plants to Control Postharvest Diseases of Sweet Cherry Fruit. Postharvest Biol. Technol. 2016, 120, 180–187. [Google Scholar] [CrossRef]
- Cheng, G.W.; Crisosto, C.H. Browning Potential, Phenolic Composition, and Polyphenoloxidase Activity of Buffer Extracts of Peach and Nectarine Skin Tissue. J. Am. Soc. Hortic. Sci. 1995, 120, 835–838. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.M.; Liu, S.X.; Zhang, D.L.; Chang, F.; Li, Y.B. Studies on Anti-Browning of Coconut (Cocos Nucifera L.) Fruit. J. Trop. Agric. 1995, 72, 254–256. [Google Scholar]
- Amiot, M.J.; Fleuriet, A.; Cheynier, V.; Nicolas, J. Phenolic Compounds and Oxidative Mechanisms in Fruit and Vegetables; Clarendon Press: Oxford, UK, 1997. [Google Scholar]
- Lea, A. Flavor, Color, and Stability in Fruit Products: The Effect of Polyphenols. Plant Polyphen. 1992, 59, 827–847. [Google Scholar]
- Herrmann, K. Flavonols and Flavones in Food Plants: A Review. Int. J. Food Sci. Technol. 1976, 11, 433–448. [Google Scholar] [CrossRef]
- Ju, Z.; Yuan, Y.; Liu, C.; Zhan, S.; Wang, M. Relationships among Simple Phenol, Flavonoid and Anthocyanin in Apple Fruit Peel at Harvest and Scald Susceptibility. J. Post. Biol. Tech. 1996, 8, 83–93. [Google Scholar] [CrossRef]
- Teissonn, C. Internal Bruising of Pineapple. Fruits 1972, 27, 329–339. [Google Scholar]
- Hodges, D.M.; Lester, G.E.; Munro, K.; Toivonen, P. Oxidative Stress: Importance for Postharvest Quality. HortScience 2004, 39, 924–929. [Google Scholar] [CrossRef]
- Szilard, L. On the Nature of the Aging Process|PNAS. Available online: https://www.pnas.org/doi/abs/10.1073/pnas.45.1.30 (accessed on 20 October 2022).
- Orgel, L.E. The Maintenance of the Accuracy of Protein Synthesis and Its Relevance to Aging. Sci. Aging. Knowledge. Env. 2001, 2001, cp8. [Google Scholar] [CrossRef]
- Chance, B.; Schoener, B.; Oshino, R.; Itshak, F.; Nakase, Y. Oxidation-Reduction Ratio Studies of Mitochondria in Freeze-Trapped Samples. NADH and Flavoprotein Fluorescence Signals. J. Biol. Chem. 1979, 254, 4764–4771. [Google Scholar] [CrossRef] [PubMed]
- Sacher, J.A. Senescence and Postharvest Physiology. Annu. Rev. Plant Physiol. 1973, 24, 197–224. [Google Scholar] [CrossRef]
- Paulin, A.; Droillard, M.J.; Bureau, J.M. Effect of a Free Radical Scavenger, 3,4,5-Trichlorophenol, on Ethylene Production and on Changes in Lipids and Membrane Integrity during Senescence of Petals of Cut Carnations (Dianthus caryophyllus). Physiol. Plant. 1986, 67, 465–471. [Google Scholar] [CrossRef]
- Crisosto, C.H.; Crisosto, G.; Neri, F. Understandig Tree Fruit Quality Based on Consumer Acceptance. Acta Hortic. 2006, 183–190. [Google Scholar] [CrossRef]
- USDA—U.S. Agricultural Research Service. National Nutrient Database for Standar Reference; USDA: Washington, DC, USA, 2017.
- Faienza, M.F.; Corbo, F.; Carocci, A.; Catalano, A.; Clodoveo, M.L.; Grano, M.; Wang, D.Q.-H.; D’Amato, G.; Muraglia, M.; Franchini, C.; et al. Novel Insights in Health-Promoting Properties of Sweet Cherries. J. Funct. Foods 2020, 69, 103945. [Google Scholar] [CrossRef]
- Gao, L.; Mazza, G. Characterization, Quantitation, and Distribution of Anthocyanins and Colorless Phenolics in Sweet Cherries. J. Agric. Food Chem. 1995, 43, 343–346. [Google Scholar] [CrossRef]
- Seeram, N.P.; Bourquin, L.D.; Nair, M.G. Degradation Products of Cyanidin Glycosides from Tart Cherries and Their Bioactivities. J. Agric. Food Chem. 2001, 49, 4924–4929. [Google Scholar] [CrossRef]
- Serrano, M.; Guillén, F.; Martínez-Romero, D.; Castillo, S.; Valero, D. Chemical Constituents and Antioxidant Activity of Sweet Cherry at Different Ripening Stages. J. Agric. Food Chem. 2005, 53, 2741–2745. [Google Scholar] [CrossRef]
- Kelebek, H.; Selli, S. Evaluation of Chemical Constituents and Antioxidant Activity of Sweet Cherry (Prunus avium L.) Cultivars. Int. J. Food Sci. Technol. 2011, 46, 2530–2537. [Google Scholar] [CrossRef]
- Hu, T.; Subbiah, V.; Wu, H.; BK, A.; Rauf, A.; Alhumaydhi, F.A.; Suleria, H.A.R. Determination and Characterization of Phenolic Compounds from Australia-Grown Sweet Cherries (Prunus avium L.) and Their Potential Antioxidant Properties. ACS Omega 2021, 6, 34687–34699. [Google Scholar] [CrossRef]
- Prusky, D. Pathogen Quiescene in Postharvest Disease. Annu. Rev. Phytopathol. 1996, 34, 413–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaovanalikit, A.; Wrolstad, R.E. Total Anthocyanins and Total Phenolics of Fresh and Processed Cherries and Their Antioxidant Properties. J. Food Sci. 2004, 69, FCT67–FCT72. [Google Scholar] [CrossRef]
- Gonçalves, B.; Landbo, A.-K.; Knudsen, D.; Silva, A.P.; Moutinho-Pereira, J.; Rosa, E.; Meyer, A.S. Effect of Ripeness and Postharvest Storage on the Phenolic Profiles of Cherries (Prunus avium L.). J. Agric. Food Chem. 2004, 52, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Commisso, M.; Bianconi, M.; Di Carlo, F.; Poletti, S.; Bulgarini, A.; Minari, F.; Negri, S.; Stocchero, M.; Ceoldo, S.; Avesani, L.; et al. Multi-Approach Metabolomics Analysis and Artificial Simplified Phytocomplexes Reveal Cultivar-Dependent Synergy between Polyphenols and Ascorbic Acid in Fruits of the Sweet Cherry (Prunus avium L.). PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Redox Sensing and Signalling Associated with Reactive Oxygen in Chloroplasts, Peroxisomes and Mitochondria. Acta Physiol. Plant. 2003, 119, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Sanmartin, M.; Drogoudi, P.D.; Lyons, T.; Pateraki, I.; Barnes, J.; Kanellis, A.K. Over-Expression of Ascorbate Oxidase in the Apoplast of Transgenic Tobacco Results in Altered Ascorbate and Glutathione Redox States and Increased Sensitivity to Ozone. Planta 2003, 216, 918–928. [Google Scholar] [CrossRef]
- Wang, P.; Yin, L.; Liang, D.; Li, C.; Ma, F.; Yue, Z. Delayed Senescence of Apple Leaves by Exogenous Melatonin Treatment: Toward Regulating the Ascorbate–Glutathione Cycle. J. Pineal Res. 2012, 53, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Vanderslice, J.T.; Higgs, D.J.; Hayes, J.M.; Block, G. Ascorbic Acid and Dehydroascorbic Acid Content of Foods-as-Eaten. J. Food Composit. Anal. 1990, 3, 105–118. [Google Scholar] [CrossRef]
- Ferretti, G.; Bacchetti, T.; Belleggia, A.; Neri, D. Cherry Antioxidants: From Farm to Table. Molecules 2010, 15, 6993–7005. [Google Scholar] [CrossRef]
- Anttonen, M.J.; Karjalainen, R.O. Evaluation of Means to Increase the Content of Bioactive Phenolic Compounds in Soft Fruits. Acta Hortic. 2009, 309–314. [Google Scholar] [CrossRef]
- Ali, L.; Alsanius, B.W.; Rosberg, A.K.; Svensson, B.; Nielsen, T.; Olsson, M.E. Effects of Nutrition Strategy on the Levels of Nutrients and Bioactive Compounds in Blackberries. Eur. Food Res. Technol. 2012, 234, 33–44. [Google Scholar] [CrossRef]
- Hanson, E.J.; Hancock, J.F. Managing the nutrient of highbush blueberries. In Bulletin E-2011; Michigan State University Extension: Paw Paw, MI, USA, 1996. [Google Scholar]
- Silva, E. Influence of Preharvest Factors on Postharvest Quality|EOrganic. Available online: http://eorganic.org/node/2670 (accessed on 20 October 2022).
- Daane, K.; Johnson, R.; Michailides, T.; Crisosto, C.; Dlott, J.; Ramirez, H.; Yokota, G.; Morgan, D. Excess Nitrogen Raises Nectarine Susceptibility to Disease and Insects. Calid. Agric. 1995, 49, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Bar-Akiva, A. Effect of Foliar Application of Nutrients on Creasing of ‘Valencia’ Oranges1. HortScience 1975, 10, 69–70. [Google Scholar] [CrossRef]
- Peck, N.H.; Dickson, M.H.; MacDonald, G.E. Tipburn Susceptibility in Semi-Isogenic Inbred Lines of Cabbage as Influenced by Nitrogen. HorstScience 1983, 18, 726–728. [Google Scholar] [CrossRef]
- Artacho, P.; Bonomelli, C. Net Primary Productivity and Allocation to Fine-Root Production in Field-Grown Sweet Cherry Trees under Different Soil Nitrogen Regimes. Sci. Hortic. 2017, 219, 207–215. [Google Scholar] [CrossRef]
- Blanpied, G.; Bramlage, W.; Dewey, D.; LaBelle, R.; Massey, L.J.; Mattus, G.; Stiles, W.; Watada, A. A Standardized Method for Collecting Apple Pressure Test Data; New York State Agricultural Experiment Station: New York, NY, USA, 1978; Available online: https://ecommons.cornell.edu/handle/1813/5080 (accessed on 25 October 2022).
- Prasad, M.; Spiers, T.M.; Fietje, G. Nitrogen Dressing Increase Fruit Size. Kiwi Fruit 1986, 21. [Google Scholar]
- Hernández-Fuentes, A.; Colinas-Leon, M.; Cortes, J.F.; Veloz, C.; Sánchez, P.G.; Alcázar, J.R. Effect of Fertilization and Storage Conditions on Postharvest Quality of Zacatecas-Type Peach (Prunus persica (L.) Batsch). Act. Hort. 2002, 594, 507–515. [Google Scholar] [CrossRef]
- Ferrante, A.; Spinardi, A.; Maggiore, T.; Testoni, A.; Gallina, P.M. Effect of Nitrogen Fertilisation Levels on Melon Fruit Quality at the Harvest Time and during Storage. J. Food. Sci. 2008, 88, 707–713. [Google Scholar] [CrossRef]
- Cheng, L.; Ma, F.; Ranwala, D. Nitrogen Storage and Its Interaction with Carbohydrates of Young Apple Trees in Response to Nitrogen Supply. Tree Physiol. 2004, 24, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claussen, W.; Lenz, F. Effect of Ammonium or Nitrate Nutrition on Net Photosynthesis, Growth, and Activity of the Enzymes Nitrate Reductase and Glutamine Synthetase in Blueberry, Raspberry and Strawberry. Plant Soil 1999, 208, 95–102. [Google Scholar] [CrossRef]
- Forde, B.G.; Clarkson, D.T. Nitrate and Ammonium Nutrition of Plants: Physiological and Molecular Perspectives. In Advances in Botanical Research; Callow, J.A., Ed.; Academic Press: Cambridge, MA, USA, 1999; Volume 30, pp. 1–90. [Google Scholar]
- Preciado-Rangel, P.; Salas-Pérez, L.; Gallegos-Robles, M.Á.; Ruiz-Espinoza, F.H.; Ayala-Garay, A.V.; Fortis-Hernández, M.; Murillo-Amador, B. Increasing Doses of Potassium Increases Yield and Quality of Muskmelon Fruits under Greenhouse. Hortic. Bras. 2018, 36, 184–188. [Google Scholar] [CrossRef]
- Mengel, K.; Kirkby, E.A. Principles of Plant Nutrition, 5th ed.; Springer Science & Business Media: Cham, Switzerland, 2012; ISBN 978-94-010-1009-2. Available online: https://books.google.cl/books?hl=es&lr=&id=sWhNDJdbLgEC&oi=fnd&pg=PA2&dq=Principles+of+Plant+Nutrition&ots=z4JU065zfI&sig=INFfeYrcVpjB_1Fv2cSyzBa0KHU#v=onepage&q=Principles%20of%20Plant%20Nutrition&f=false (accessed on 24 October 2022).
- Kader, A.A. Postharvest Technology of Horticultural Crops; University of California Agriculture and Natural Resources: Davis, CA, USA, 2002; ISBN 978-1-879906-51-8. [Google Scholar]
- Tabatabaei, S.J.; Yusefi, M.; Hajiloo, J. Effects of Shading and NO3:NH4 Ratio on the Yield, Quality and N Metabolism in Strawberry. Sci. Hortic. 2008, 116, 264–272. [Google Scholar] [CrossRef]
- Neuweiler, R. Nitrogen Fertilization in Integrated Outdoor Strawberry Production. Proc. Int. Soc. Hortic. Sci. 1997, 439, 747–752. Available online: https://www.actahort.org/books/439/439_125.htm (accessed on 24 October 2022). [CrossRef]
- Mukkun, L.; Singh, Z.; Phillips, D. Nitrogen Nutrition Affects Fruit Firmness, Quality and Shelf Life of Strawberry. Acta Hortic. 2001, 553, 69–71. [Google Scholar] [CrossRef]
- Koumanov, K.S.; Staneva, I.N.; Kornov, G.D.; Germanova, D.R. Sweet Cherry Fruit Quality under Fertigation. Acta Hortic. 2016, 551–558. [Google Scholar] [CrossRef]
- Martin, D. Calcium Movement in Apple Trees. Experiments in Tasmania. Div. Plant Ind. 1967, 6, 49–54. [Google Scholar]
- Smock, R.M.; Gross, C.R. Studies on respiration of apples. Mem. Cornell Agric. Exp. Stn. 1950, 297, 45. [Google Scholar]
- Gourley, J.H.; Hopkins, E.F. Some Relations of Nitrogen to Keeping Quality of Fruit. J. Am. Soc. Hortic. Sci. 1929, 26, 167–173. [Google Scholar]
- Huang, Z.-A.; Jiang, D.-A.; Yang, Y.; Sun, J.-W.; Jin, S.-H. Effects of Nitrogen Deficiency on Gas Exchange, Chlorophyll Fluorescence, and Antioxidant Enzymes in Leaves of Rice Plants. Photosynthetica 2004, 42, 357–364. [Google Scholar] [CrossRef]
- Pompelli, M.F.; Martins, S.C.V.; Antunes, W.C.; Chaves, A.R.M.; DaMatta, F.M. Photosynthesis and Photoprotection in Coffee Leaves Is Affected by Nitrogen and Light Availabilities in Winter Conditions. J. Plant. Physiol. 2010, 167, 1052–1060. [Google Scholar] [CrossRef]
- Logan, B.A.; Demmig-Adams, B.; Rosenstiel, T.N.; Adams, W.W., III. Effect of Nitrogen Limitation on Foliar Antioxidants in Relationship to Other Metabolic Characteristics. Planta 1999, 209, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Sánchez, F.; Santos, E.; Kerstupp, S.; Villagómez, R.; Scheinvar, L. Colorant Extract from Red Prickly Pear (Opuntia lasiacantha) for Food Application. J. Agric. Food Chem. 2006, 5, 1330–1337. [Google Scholar]
- Jezek, M.; Zörb, C.; Merkt, N.; Geilfus, C.-M. Anthocyanin Management in Fruits by Fertilization. J. Agric. Food Chem. 2018, 66, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Strissel, T.; Halbwirth, H.; Hoyer, U.; Zistler, C.; Stich, K.; Treutter, D. Growth-Promoting Nitrogen Nutrition Affects Flavonoid Biosynthesis in Young Apple (Malus domestica Borkh.) Leaves. Plant Biol. 2005, 7, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Li, C.; Fu, X.; Li, D.; Li, L.; Chen, X.; Wu, H.; Cui, X.; Zhang, X.; Shen, H.; et al. Effects of Nitrate Deficiency on Nitrate Assimilation and Chlorophyll Synthesis of Detached Apple Leaves. Plant Physiol. Biochem. 2019, 142, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Rubio Ames, Z.; Brecht, J.K.; Olmstead, M.A. Nitrogen Fertilization Rates in a Subtropical Peach Orchard: Effects on Tree Vigor and Fruit Quality. J. Agric. Food Chem. 2020, 100, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Sha, J.; Chen, Q.; Xu, X.; Zhu, Z.; Ge, S.; Jiang, Y. Exogenous Abscisic Acid Regulates Distribution of 13C and 15N and Anthocyanin Synthesis in ‘Red Fuji’ Apple Fruit Under High Nitrogen Supply. Front. Plant Sci. 2020, 10, 1738. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Xu, X.; Jia, Z.; Hou, X.; Chen, Q.; Sha, J.; Liu, Z.; Zhu, Z.; Jiang, Y.; Ge, S. Nitrification Inhibitor 3,4-Dimethylpyrazole Phosphate Application During the Later Stage of Apple Fruit Expansion Regulates Soil Mineral Nitrogen and Tree Carbon–Nitrogen Nutrition, and Improves Fruit Quality. Front. Plant Sci. 2020, 11, 764. [Google Scholar] [CrossRef]
- Stefanelli, D.; Goodwin, I.; Jones, R. Minimal Nitrogen and Water Use in Horticulture: Effects on Quality and Content of Selected Nutrients. Food Res. Int. 2010, 43, 1833–1843. [Google Scholar] [CrossRef]
- Li, J.; Zhu, Z.; Gerendás, J. Effects of Nitrogen and Sulfur on Total Phenolics and Antioxidant Activity in Two Genotypes of Leaf Mustard. J. Plant Nut. 2008, 31, 1642–1655. [Google Scholar] [CrossRef]
- Sharafzadeh, S.; Esmaeili, M.; Mohammadi, A.H. Interaction effects of nitrogen, phosphorus and potassium on growth, essential oil and total phenolic content of sweet basil. Adv. Environ. Biol. 2011, 5, 1285–1290. [Google Scholar]
- Bryant, J.P.; Chapin, F.S.; Klein, D.R. Carbon/Nutrient Balance of Boreal Plants in Relation to Vertebrate Herbivory. Oikos 1983, 40, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Winter, C.K.; Davis, S.F. Organic Foods. J. Food Sci. 2006, 71, R117–R124. [Google Scholar] [CrossRef]
- Mitchell, A.E.; Hong, Y.-J.; Koh, E.; Barrett, D.M.; Bryant, D.E.; Denison, R.F.; Kaffka, S. Ten-Year Comparison of the Influence of Organic and Conventional Crop Management Practices on the Content of Flavonoids in Tomatoes. J. Agric. Food Chem. 2007, 55, 6154–6159. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.G.; Hartley, S.E. A Protein Competition Model of Phenolic Allocation. Oikos 1999, 86, 27–44. [Google Scholar] [CrossRef]
- Hachiya, T.; Watanabe, C.K.; Fujimoto, M.; Ishikawa, T.; Takahara, K.; Kawai-Yamada, M.; Uchimiya, H.; Uesono, Y.; Terashima, I.; Noguchi, K. Nitrate Addition Alleviates Ammonium Toxicity Without Lessening Ammonium Accumulation, Organic Acid Depletion and Inorganic Cation Depletion in Arabidopsis thaliana Shoots. Plant Cell Physiol. 2012, 53, 577–591. [Google Scholar] [CrossRef]
- Stewart, A.J.; Chapman, W.; Jenkins, G.I.; Graham, I.; Martin, T.; Crozier, A. The Effect of Nitrogen and Phosphorus Deficiency on Flavonol Accumulation in Plant Tissues. Plant Cell Environ. 2001, 24, 1189–1197. [Google Scholar] [CrossRef]
- Larbat, R.; Le Bot, J.; Bourgaud, F.; Robin, C.; Adamowicz, S. Organ-Specific Responses of Tomato Growth and Phenolic Metabolism to Nitrate Limitation. Plant Biol. 2012, 14, 760–769. [Google Scholar] [CrossRef]
- Yañez-Mansilla, E.; Cartes, P.; Díaz, M.R.; Ribera-Fonseca, A.; Alberdi, M. Photosynthetic and Antioxidant Performance Are Differentially Affected by Short-Term Nitrogen Supply in Highbush Blueberry Cultivars. Cienc. Investig. Agrar. 2014, 41, 61–70. [Google Scholar] [CrossRef]
- Lee, S.K.; Kader, A.A. Preharvest and Postharvest Factors Influencing Vitamin C Content of Horticultural Crops. Postharvest Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Lisiewska, Z.; Kmiecik, W. Effects of Level of Nitrogen Fertilizer, Processing Conditions and Period of Storage of Frozen Broccoli and Cauliflower on Vitamin C Retention. Food Chem. 1996, 57, 267–270. [Google Scholar] [CrossRef]
- Nagy, P.T.; Thurzó, S.; Szabó, Z.; Nyéki, J.; Silva, A.P.; Gonçalves, B. Influence of Foliar Fertilization on Mineral Composition, Sugar and Organic Acid Content of Sweet Cherry. Acta Hortic. 2010, 868, 353–358. [Google Scholar] [CrossRef]
- Sorensen, N. Dietary Fiber and Ascorbic Acid in White Cabbage as Affected by Fertilization. Act. Hort 1984, 1984. 163, 221–230. [Google Scholar] [CrossRef]
- Freyman, P.; Toivonen, P.; Lin, W.C.; Perrin, P.W.; Hall, J.W. Effect of Nitrogen Fertilization on Yield, Storage Losses and Chemical Composition of Winter Cabbage. Can. J. Plant Sci. 1991, 71, 943–946. [Google Scholar] [CrossRef]
- Cardeñosa, V.; Medrano, E.; Lorenzo, P.; Sanchez-Guerrero, M.; Cuevas, F.; Pradas, I.; Moreno-Rojas, J. Effects of Salinity and Nitrogen Supply on the Quality and Health-related Compounds of Strawberry Fruits (Fragaria × ananassa Cv. Primoris). J. Sci. Food Agric. 2015, 95, 2924–2930. [Google Scholar] [CrossRef] [PubMed]
- Rutkowski, K.; Łysiak, G.P.; Zydlik, Z. Effect of Nitrogen Fertilization in the Sour Cherry Orchard on Soil Enzymatic Activities, Microbial Population, and Fruit Quality. Agriculture 2022, 12, 2069. [Google Scholar] [CrossRef]
- Rutkowski, K.; Łysiak, G.P. Weather Conditions, Orchard Age and Nitrogen Fertilization Influences Yield and Quality of ‘Łutówka’ Sour Cherry Fruit. Agriculture 2022, 12, 2008. [Google Scholar] [CrossRef]
- Guo, K.; Peng, L.; Hong, Y.; Qiao, G. Optimizing Nitrogen, Phosphorus, and Potassium Fertilization Rates for Fruit Performance of Chinese Cherry (Prunus pseudocerasus Lindl.). Int. J. Fruit Sci. 2022, 22, 769–778. [Google Scholar] [CrossRef]
- Augustin, J. Variations in the Nutritional Composition of Fresh Potatoes. J. Food. Sci. 1975, 40, 1295–1299. [Google Scholar] [CrossRef]
- Mozafar, A. Nitrogen Fertilizers and the Amount of Vitamins in Plants: A Review. J. Plant Nut. 1993, 16, 2479–2506. [Google Scholar] [CrossRef]
- Bai, Q.; Shen, Y.; Huang, Y. Advances in Mineral Nutrition Transport and Signal Transduction in Rosaceae Fruit Quality and Postharvest Storage. Front. Plant. Sci. 2021, 12, 620018. [Google Scholar] [CrossRef] [PubMed]
- Knowles, L.; Trimble, M.R.; Knowles, N.R. Phosphorus Status Affects Postharvest Respiration, Membrane Permeability and Lipid Chemistry of European Seedless Cucumber Fruit (Cucumis sativus L.). J. Postharvest Biol. Tech. 2001, 21, 179–188. [Google Scholar] [CrossRef]
- Afroz, C.A.; Shimul, M.A.; Siddiky, M.A.; Razzaque, M.A. Effects of Nitrogen, Phosphorus, Potassium and Sulphur on Growth, Yield and Nutrient Content of Strawberry (Fragaria ananassa). Ph.D. Thesis, Sher-e-Bangla Agricultural University, Dhaka, Bangladesh, 2011. Available online: http://archive.saulibrary.edu.bd:8080/xmlui/handle/123456789/1944 (accessed on 24 October 2022).
- Valentinuzzi, F.; Mason, M.; Scampicchio, M.; Andreotti, S.; Mimmo, T. Enhancement of the Bioactive Compound Content in Strawberry Fruits Grown under Iron and Phosphorus Deficiency. J. Sci. Food Agric. 2014, 95, 2088–2094. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Guan, C.; Dai, H.; Li, X.; Zhang, Z. Soluble Solids Content Is Positively Correlated with Phosphorus Content in Ripening Strawberry Fruits. Sci. Hortic. 2015, 195, 183–187. [Google Scholar] [CrossRef]
- Valentinuzzi, F.; Pii, Y.; Vigani, G.; Lehmann, M.; Cesco, S.; Mimmo, T. Phosphorus and Iron Deficiencies Induce a Metabolic Reprogramming and Affect the Exudation Traits of the Woody Plant Fragaria × ananassa. EXBOTJ 2015, 66, 6483–6495. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Liu, W.; Feng, Y.; Li, D.; Li, K.; Sun, Q.; Zhai, L.; Wu, T.; Zhang, X.; Xu, X.; et al. Ethylene Response Factors MbERF4 and MbERF72 Suppress Iron Uptake in Woody Apple Plants by Modulating Rhizosphere PH. Plant Cell Physiol. 2020, 61, 699–711. [Google Scholar] [CrossRef]
- Matubara, H.; Miharu, K.; Kinoshita, K.; Koyama, K.; Ye, Y.; Takahashi, K.; Yoshimura, I.; Yamamoto, Y.; Miura, Y.; Kinoshita, Y. Inhibitory Effect of Lichen Metabolites and Their Synthetic Analogues on Melanin Biosynthesis in Cultured B-16 Mouse Melanoma Cells. Nat. Prod. Sci. 1998, 4, 161–169. [Google Scholar]
- Ross, K.; Neilsen, G.; Neilsen, D. The Effect of Irrigation Frequency, Phosphorus Fertigation, and Cultivar on Levels of Phenolic Compounds in Sweet Cherries. HortScience 2018, 53, 1507–1512. [Google Scholar] [CrossRef] [Green Version]
- Lux-Endrich, A.; Treutter, D.; Feucht, W. Influence of Nutrients and Carbohydrate Supply on the Phenol Composition of Apple Shoot Cultures. J. Plant. C. 2000, 60, 15–21. [Google Scholar] [CrossRef]
- Estrada-Ortiz, E.; Trejo-Téllez, L.I.; Gómez-Merino, F.C.; Núñez-Escobar, R.; Sandoval-Villa, M. The Effects of Phosphite on Strawberry Yield and Fruit Quality. J. Soil. Sci. Plant. Nutr. 2013, 13, 612–620. [Google Scholar] [CrossRef] [Green Version]
- Kader, A. Influence of Preharvest and Postharvest Environment on Nutritional Composition of Fruits and Vegetables; Hortic. Human Health; F.A. Bliss.; Prentice Hall: Hoboken, NJ, USA, 1988. [Google Scholar]
- Ge, Y.; Chen, Y.; Li, C.; Wei, M.; Li, X.; Li, S.; Lu, S.; Li, J. Effect of Trisodium Phosphate Dipping Treatment on the Quality and Energy Metabolism of Apples. J. Agric. Food Chem. 2019, 274, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Yogaratnam, N.; Sharples, R.O. Supplementing the Nutrition of Bramley’s Seedling Apple with Phosphorus Sprays II. Effects on Fruit Composition and Storage Quality. J. Hortic. Sci. 1982, 57, 53–59. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology. Ann. Bot. 2003, 91, 750–751. [Google Scholar] [CrossRef] [Green Version]
- Perring, M.A. The Mineral Composition of Apples. II.—Analytical Methods. J. Agric. Food Chem. 1964, 15, 752–759. [Google Scholar] [CrossRef]
- Petriccione, M.; De Sanctis, F.; Pasquariello, M.S.; Mastrobuoni, F.; Rega, P.; Scortichini, M.; Mencarelli, F. The Effect of Chitosan Coating on the Quality and Nutraceutical Traits of Sweet Cherry During Postharvest Life. Food Bioprocess Technol. 2015, 8, 394–408. [Google Scholar] [CrossRef]


| Species | Treatments | Nitrogen Treatment | Effects | References |
|---|---|---|---|---|
| Prunus avium L. | Fertigation | Annual Rate 160 kg ha−1 Decreasing monthly application | No effect with last month N | [112] |
| Prunus cerasus L. | 34% ammonium nitrate | 0, 60, 120 kg N ha−1 | Reduction of fruit weight, extract content, acidity, and ratio of the total soluble solids to a titratable acidity | [143,144] |
| Prunus pseudocerasus Lindl. | Urea | 0–0.8 kg plot | N content showed an optimum below the highest dose | [145] |
| Malus domestica L. | 0–5–10–15–20 mM NH4NO3 | 15–20 mM NH4NO3 | Increase nonstructural carbohydrates increase the nitrogen concentration in plants, decrease of the C/N ratios | [103] |
| Prunus Avium L. | 50% of shading and NO3:NH4 ratios (0:100, 75:25, 50:50 and 25:75) | 75:25 and 50:50—Higher NO3 ratios—Higher NH4 ratios | Increased yield, fruit size. Increased titratable acidity. Increased total soluble content | [109] |
| Fragaria ananassa | 0–60–120 Kg N ha−1 | 60–120 Kg ha−1 | No effect on the fruit quality | [110] |
| Brassica Juncea Coss | 10–25 mM N and 0.5–1.0–2.0 mM S | 25 mM N | Increased antioxidant activity and decreased total phenolic concentration | [127] |
| Fragaria ananassa | 150–225–300–450–600 Kg ha−1 N | 450–600 Kg ha−1 N | Decreased solid soluble content and increased blemishes in fruits and acid content | [111] |
| Ocimum basilicum | N,P, K (1:1:1), 6 different levels (0, 25, 50, 75, 100 and 125 mg kg−1 soil) | 50 mg Kg−1 of soil | Increased total phenolic content | [128] |
| Rubus ulmifolius | 60–100 Kg ha−1 N and 66.4–104 Kg ha −1 K | 100 Kg ha−1 N | Increased fructose, glucose, sucrose content, pH and secondary metabolites in fruit | [92] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palacios-Peralta, C.; Reyes-Díaz, M.; González-Villagra, J.; Ribera-Fonseca, A. The Potential Roles of the N and P Supplies on the Internal Browning Incidence in Sweet Cherries in the Southern Chile. Horticulturae 2022, 8, 1209. https://doi.org/10.3390/horticulturae8121209
Palacios-Peralta C, Reyes-Díaz M, González-Villagra J, Ribera-Fonseca A. The Potential Roles of the N and P Supplies on the Internal Browning Incidence in Sweet Cherries in the Southern Chile. Horticulturae. 2022; 8(12):1209. https://doi.org/10.3390/horticulturae8121209
Chicago/Turabian StylePalacios-Peralta, Cristóbal, Marjorie Reyes-Díaz, Jorge González-Villagra, and Alejandra Ribera-Fonseca. 2022. "The Potential Roles of the N and P Supplies on the Internal Browning Incidence in Sweet Cherries in the Southern Chile" Horticulturae 8, no. 12: 1209. https://doi.org/10.3390/horticulturae8121209
APA StylePalacios-Peralta, C., Reyes-Díaz, M., González-Villagra, J., & Ribera-Fonseca, A. (2022). The Potential Roles of the N and P Supplies on the Internal Browning Incidence in Sweet Cherries in the Southern Chile. Horticulturae, 8(12), 1209. https://doi.org/10.3390/horticulturae8121209