1. Introduction
Solanum melongena L., commonly known as eggplant, varies significantly in color, size, and form, depending on the specific variety [
1]. Even though eggplant is commonly thought of as a vegetable, it is botanically classified as a fruit. The world production of eggplant is around 55.2 million tons [
2]. It plays an important role as a food crop in many countries and is most widespread in Asia, North African, and Southern European cuisines. One of the most popular cultivars of eggplant in Japan today is a variety of “Shikon Sendai Naga”, a bluish-purple pericarp with an elongated shape, and it is perfect for making pickles, served as a side dish [
3].
For its high phenolic content, eggplant is considered to be one of the greatest vegetables in terms of oxygen radical absorbance capacity [
4]. Aside from its antioxidant activity, related polyphenols and anthocyanins also demonstrate various effective biological activities, including anti-inflammatory, antimutagenic, and antiproliferative activities [
5]. It has been reported that chlorogenic acid (5-
O-caffeoylquinic acid) is the main component and represents 50–96% of all phenolic compounds in eggplant [
6]. However, a significant decrease of six-fold in the level of chlorogenic acid was observed in the first week of storage. Therefore, producing eggplant, particularly for higher levels of antioxidant and bioactive compounds, is an attempt to increase dietary antioxidant and health benefit intake.
Light can influence biological processes, whereby photo-stress can activate defense systems, resulting in an increase in the accumulation of phenylpropanoids, such as phenolic acids and flavonoids [
7]. Generally, photoreceptors that intercept visible wavebands, including phytochromes, cryptochromes, and phototropins, have a synergistic influence on plant development [
8]. Various light wavelengths have different effects on plant physiology and secondary metabolic compounds. Red light is considered to be essential for the photosynthetic apparatus and alters plant morphology through a response involving the action of phytochromes [
8]. Blue light, conversely, mediated mostly by cryptochromes and phototropins, stimulates the biosynthesis of secondary metabolites that have photoprotective functions, such as carotenoid and anthocyanin [
8]. This explains the selective use of light-emitting diodes (LEDs), which produce minimal longwave radiation that lacks external heat, unlike fluorescent or incandescent light [
9]. Currently, LEDs have frequently been applied as a light source, emitting near infrared (NIR) radiation for indoor plant growth [
8]. Irradiation with LEDs of different wavelengths has been studied for color development and the accumulation of bioactive compounds during the growth of several fruits and vegetables [
7,
8].
The effects of light quality and quantity are versatile and can lead to obtaining a variety of results. The response highly depends on the plant species, genotype, and growing condition. Blue and red LED treatments, for example, could improve the growth of different cultivars and microgreens, while also increasing the content of phenolic compounds, which are valuable phytochemicals. However, this depends on the pigmentation of the cultivar/species. The green cultivars such as broccoli sprouts are the most stimulated by a higher proportion of red light, while the red cultivars react the most to higher ratios of blue light [
10]. The nutritional content of different sprouts (radish, soybean, mung bean, and pumpkin) grown under different light conditions (red, blue, and white light) depends on the dark or specific spectral wavelength used for their growth [
11]. However, an increase in polyphenols was induced only in soybeans and only by red light. For the results during postharvest storage, irradiation with different light spectra has been studied in broccoli [
12], tomato [
13], cherry tomato [
9], and apple [
7]. Postharvest blue-light irradiation appears to enhance color and nutrition quality of red apples [
7] and strawberry [
14]. Red LEDs irradiation positively influence metabolic processes and contribute to a higher content of health-promoting substances in tomatoes [
13].
The combination of red and blue LED light is one of the most efficient light sources for plant growth and development, as well as improving the quality of plant nutrition [
15]. According to a previous report, red and blue (1:1) LEDs increased the flavonoid and ferric-reducing antioxidant potential (FRAP) during tomato fruit development [
15]. Moreover, red (70%) and blue (30%) LED light enhances the antioxidant capacity in lemon balm plants [
16].
Despite the fact that there have been consistent studies on plants grown under LED lighting, up to the best of our knowledge, the effect of different LED wavelengths on the postharvest of eggplants has not been studied and reported. In this work, the effect of red, blue, or the combination of red and blue (red + blue) LEDs irradiation during postharvest to increase antioxidant activity and bioactive compounds such as carotenoid, flavonoid, anthocyanin, and chlorogenic acid contents of eggplant fruits was elucidated and demonstrated. It is recognized that the phytochemical compounds of fruits vary between parts of the same fruit [
17]. In addition to the tailored changes in fruit quality under different LEDs lights, variations in the effect of LEDs light on the phytochemical compounds of the fruit components should be considered. Thus, the investigation was also extended to measure the antioxidant activity and bioactive compounds accumulated in the peel and flesh components of eggplants.
2. Materials and Methods
2.1. Plant Materials and LEDs Irradiation
The fresh fruits of the Japanese eggplant (
Solanum melongena L.) variety, of a bluish purple with an elongated shape, named “Shikon Sendai Naga” [
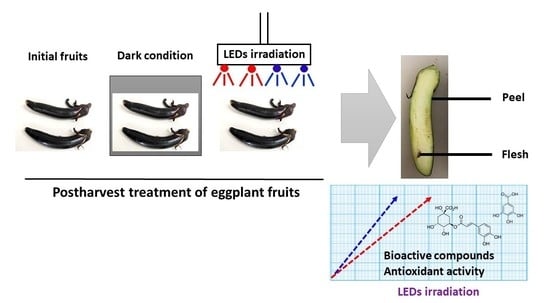
3], at commercial maturity with an intermediate size (fruit length of 8–10 cm, weight 17–22 g) were manually harvested after 30 ± 5 days of anthesis by Hsu Chuan Foods Co., Ltd. (Chiang Rai, Thailand). Eggplants were transported to our laboratory by a temperature controlled vehicle within 1 h. After selection, having being free of defects and diseases, eggplants were cleaned in circulated tap water and dried at room temperature. The eggplants were sampled on harvesting day, and they were used as an initial condition prior to exposing the fruit samples to darkness or LEDs irradiance.
To investigate the effect of LED irradiation, 30 eggplants per treatment were exposed under continuous red (650–660 nm), blue (450–460 nm), and red + blue (red 70%, blue 30%) LED light (Growlab Agritech Co., Ltd., Bangkok, Thailand) for 2 days in a controlled room temperature at 30 ± 5 °C, and 70% relative humidity (RH). Lighting array of LEDs was 30 cm distance above the eggplant samples. The light intensity was maintained to 55 µmol photons/m2/s, measured with a LI-COR Quantum meter (Li-Cor Inc., Lincoln, NE, USA). The control was stored in darkness in the same conditions. Samples were collected by separating the peel and flesh in strips of uniform thickness before further analysis.
2.2. Firmness Measurement
Eggplant firmness of 10 randomly selected fruits per treatment was measured by the TA-XT plus texture analyzer with a p/2 probe. Penetration tests were conducted with p/2-cylinder probe, penetrating in 6 mm. The loading rate was 1 mm/s. The firmness was determined to be the mean of the maximum force (newton, N).
2.3. Color Determination
Ten eggplants per treatment were randomly selected and measured for surface color by the L* a* b* system using a colorimeter (Minolta, Japan). Color components L*, a*, and b* were measured on three random points on the surface of eggplant fruits and reported as L* (lightness component) and the other two chromatic components, a* (from green to red) and b* (from blue to yellow). Hue angle (h°) was calculated as h° = tan−1(b*/a*) when a* and b* > 0.
2.4. Carotenoid Content Analysis
Carotenoid content of flesh and peel tissues was determined according to Davies et al. [
18] with some modifications. One gram of sample was homogenized and extracted with 10 mL ethanol. Then, the extract sample was sonicated for 15 min and centrifuged at 10,000×
g for 10 min. The absorbance of supernatant was measured using an Evolution 201 UV-Vis spectrophotometer (Thermo Scientific, Madison, WI, USA) at 450 nm. Total carotenoid content was evaluated using a standard curve of β-carotene and expressed as milligrams per gram extracts of β-carotene equivalents.
2.5. Anthocyanin Content Analysis
Peel tissues (1 g) was homogenized and extracted with 15 mL HCl-methanol (0.15% HCl: 95% methanol = 15:85) for 4 h. The extract was centrifuged at 10,000×
g for 20 min and the supernatant was measured for absorbance at 530, 620, and 650 nm. The anthocyanin content was based on the formula: ∆
A/mL = (
A530 −
A620) − 0.1(
A650 −
A620) [
19]. The anthocyanin content was expressed as ∆
A/100 g fresh weight (FW).
2.6. Total Phenolic Content Analysis
Eggplant samples were selected and peeled to separate the peel and flesh. Total phenolic content was analyzed using the method of Ketsa and Atantee [
20]. Fresh flesh or peel (3 g) was homogenized in 25 mL of 80% methanol and centrifuged at 10,000×
g for 20 min at 4 °C. The obtained supernatant was used to analyze the bioactive compounds and antioxidant activity. The reaction was started when 1 mL of supernatant was mixed with Folin-Ciocalteu reagent solution and 2 mL of saturated Na
2CO
3. The mixture was then incubated in the dark at room temperature for 60 min. Total phenolic content was determined by measuring the absorbance at 765 nm and evaluated using a standard curve of gallic acid. The contents were expressed as milligrams of gallic acid equivalent per 100 g fresh weight (mg GAE/100 g FW).
2.7. Total Flavonoid Content Analysis
Total flavonoid content was adapted from the method of Youryon and Supapvanich [
21] with slight modifications. The reaction was started when 0.5 mL of supernatant was added into a mixture of 1.25 mL of distilled water and 75 µL of 0.5% NaNO
2. After being left for 6 min, 150 µL of 10% AlCl
3·6H
2O and 0.5 mL of 1 M NaOH were added to the mixture. Total flavonoids content was determined by measuring the absorbance at 510 nm. The data were expressed as milligrams of catechin equivalent per 100 g fresh weight (mg CE/100 g FW).
2.8. Determination of Antioxidant Activity by DPPH and FRAP Assays
The antioxidant activity was determined using a DPPH (2,2-diphenyl-2-picrylhydrazyl) radical scavenging assay according to the method of Brand-Williams et al. [
22], with slight modifications. The supernatant was obtained from the same method of total phenolic measurement. The supernatant (150 µL) was added to 2850 µL DPPH in methanol. The reaction mixture was mixed and allowed to stand in the dark for 60 min at room temperature. The absorbance of the solution was measured at 515 nm using an Evolution 201 UV-Vis spectrophotometer. The percentage of inhibition assessed as antioxidant activity was calculated against blank:
where A blank is the absorbance of the control reaction (containing all reagents except the test sample) and A sample is the absorbance of the reaction mixture.
The FRAP assay was done according to Thaipong et al. [
23] with some modifications. The working solution was freshly prepared by mixing 25 mL of 300 mM acetate buffer (3.1 g C
2H
3NaO
2·3H
2O and 16 mL C
2H
4O
2), pH 3.6, 2.5 mL of 10 mM TPTZ (2,4,6-tripyridyl-
s-triazine) solution, and 2.5 mL of 20 mM FeCl
3·6H
2O, and was then warmed at 37 °C. The supernatant of eggplant extracts (150 µL) was added to 2850 µL of the FRAP solution and allowed to react for 30 min in dark conditions. The absorbance of the color product (ferrous tripyridyltrizine complex) was measured at 593 nm. The standard curve was linear between FeSO
4 and FRAP solution. Results are expressed in µmol Fe(II)/100 g fresh weight.
2.9. HPLC Analysis of Gallic and Chlorogenic Acids
The quantification of gallic acid and chlorogenic acid was carried out using a RP-HPLC (Jasco, Japan) at 280 and 325 nm, respectively, with an external standard of authentic gallic and chlorogenic acids. A binary mobile phase gradient of methanol in 0.01% phosphoric acid was used, according to Mennella et al. [
24] with minor modifications. The gradient program of the mobile phase at 0–15 min, a linear increase from 5 to 25% methanol; 15–28 min, linear increase from 25 to 50% methanol; 28–30 min, linear increase from 50 to 100% methanol; 30–32 min, 100% methanol; 32–36 min, linear decrease from 100 to 5% methanol; 36–43 min, 5% methanol. The flow rate was 0.8 mL/min.
2.10. Statistical Analysis
The experiments were conducted using a completely randomized design (CRD). An analysis of variance (ANOVA) to evaluate treatment effects was performed using Duncan’s multiple range test (p < 0.05) by SPSS software program version 18. The data were presented as the mean values ± SD. Except for the analysis of firmness and surface color of eggplant fruits (n = 10), all data are expressed as means and standard deviation (SD) (n = 3).
4. Discussion
The postharvest quality of fruit and vegetables is influenced by tissue turgidity, which is primarily measured by the dehydration during storage [
12]. In our study, LED light treatment had no significant effect on eggplant firmness, although red and blue LEDs slightly decreased the fruit firmness (
Table 1). Ngcobo et al. [
9] also reported that LED light did not have a main effect on the activity of the enzymes involved in cell wall degradation in cherry tomato, which belong to the same genus as eggplant.
It has been reported that after harvest, the intensity, duration, and quality of light have a significant impact on phytochemical accumulation [
13]. Light affects pigment metabolism, enhancing the color of various vegetables, such as tomatoes, broccoli, and other crops [
9,
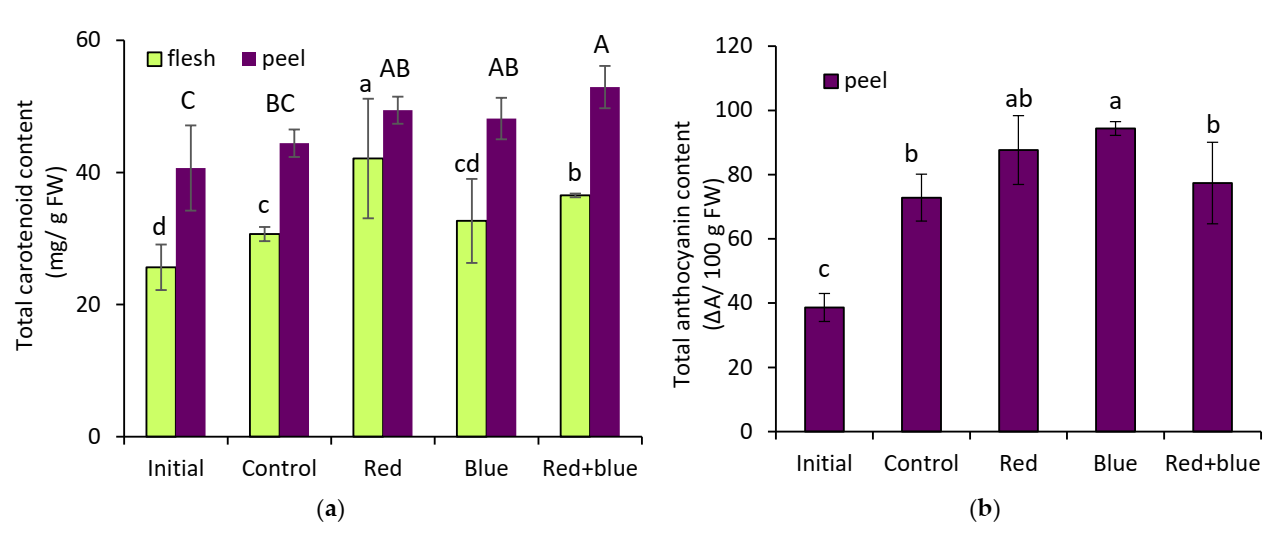
25]. In present study, carotenoid content was greatly increased by irradiation with red LED light. Red + blue LEDs with 70% red and 30% blue also increased the concentration of total carotenoid more than blue light alone in the flesh of eggplant during postharvest (
Figure 1a). The result of our study is in accordance with the findings of Ngcobo et al. [
9], who found that red light affects the concentration or presence of carotenoids in the postharvest of red tomato tissues, and who also proposed that phytochrome, a photoreceptor response to red wavelength, appears to promote carotenoid biosynthesis in green mature tomato tissues. Samuolienė et al. [
25] showed that red LED light apparently had a stronger effect than blue on the color-altering steps of baby leaf lettuce. In contrast, the highest carotenoid content was detected in broccoli stored under continuous postharvest white–blue LED [
12].
Anthocyanins are red plant pigments that, among many other roles, protect plants from photo inhibition and photo damage by absorbing solar radiation [
25]. In this study, blue LEDs enhanced the concentration of anthocyanin in eggplant peel. The effect of red light on anthocyanin accumulation was relatively low, but still showed a positive effect when compared to red + blue light (
Figure 1b). It was reported that blue light regulates anthocyanin biosynthesis through the phytochrome system [
25]. Anthocyanin synthesis is light-regulated, and the red leaf color appears on lettuce leaves in varying intensities and distributions. Blue light treatment improved total anthocyanin content by activating its related enzymes during the postharvest of strawberry [
14]. Li et al. [
26] also demonstrated the involvement of apple anthocyanin production through a blue-light photoreceptor cryptochrome.
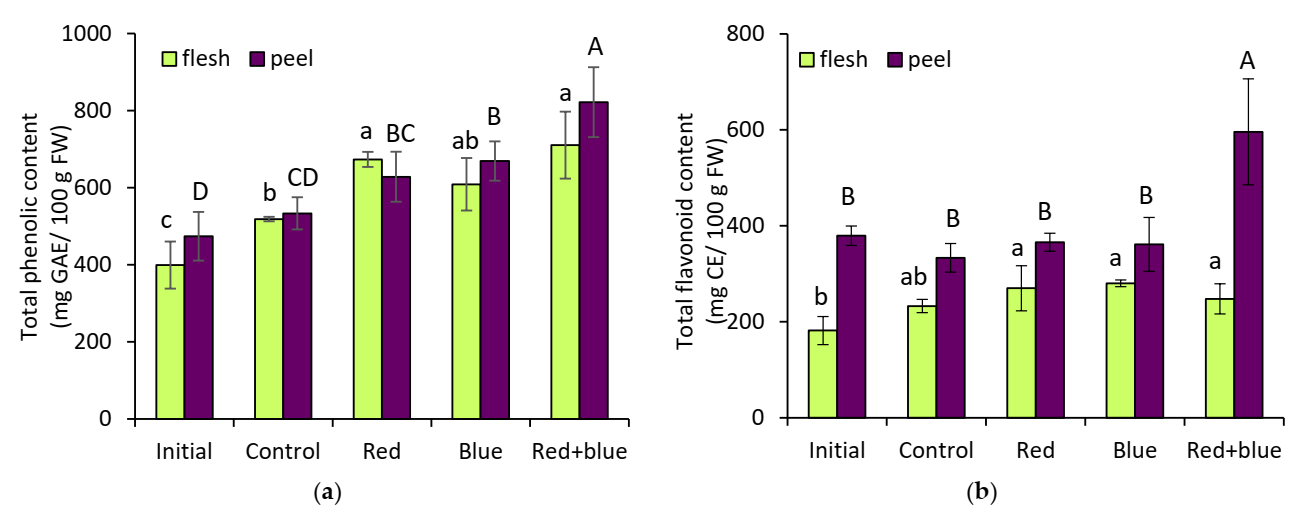
The total phenolic content was related to the accumulation of phenolic compounds, including total anthocyanin content (
Figure 1b), total flavonoid content (
Figure 2b), gallic acid, and chlorogenic acid contents (
Table 2). Our findings are in agreement with a previous study of Mannella et al. [
24], who have observed the significantly correlation of chlorogenic acid content to the total phenolic amount in eggplants. In addition to the pigment of anthocyanin, it appeared that the eggplant peel also contained highly bioactive compounds, thus, increasing the antioxidant capacity. Our results indicated that the highest total phenolic content was correlated with a significant increase in the DPPH free-radical scavenging capacity and FRAP assay in the postharvest eggplant under red + blue irradiation. A previous study of various eggplant species revealed that total phenolic compounds and chlorogenic acid appear to have the main influence on antioxidant capacity [
24]. For the results regarding plant pigment, light has been reported to affect pigment metabolism, enhancing the color of various vegetables, such as tomatoes, broccoli, and other crops [
25]. The pigment groups of plants confer certain health benefits to humans, acting as powerful antioxidants.
Bioactive compounds, including phenolic acids and flavonoids, play an important role in scavenging free radicals and oxidants to protect cell damage [
27]. In the present study, a significant increase in the level of chlorogenic acid was observed in red + blue LED-treated eggplant. The increase in bioactive compounds in red + blue LED-exposed eggplants is probably caused by the stimulation of genes, owing to the phenylpropanoid pathway, which is related to the biosynthesis of phenolic acids and flavonoid content in plants [
8,
28]. The other explanation for the higher antioxidant activity of plants exposed to blue LEDs is that the higher energy levels of short wavelengths (blue at 450–460 nm for example) stimulate oxidative stress, thus enhancing the scavenging of reactive oxygen species [
16]. Park et al. [
29] recently demonstrated that exposure to blue LED light can enhance the production of the majority of phenolic compounds, including gallate, quercetin, caffeate, and chlorogenic acid in
Brassica juncea sprouts. The effect of red LEDs in combination with blue LEDs has been observed by Ahmadi et al. [
16] in lemon balm, where red + blue lights maintained higher antioxidant and phenylalanine ammonialyase enzyme activities. One of the reasons for the high production of bioactive compounds and antioxidant activities is the presence of red and blue wavelengths in the light spectrum. The higher energy levels of blue wavelengths and the induction of oxidative stress due to the presence of red wavelengths, both of which exist in the spectrum of red + blue LED light [
16].
Polyphenols with antioxidant activity, predominantly found in the peel, represent the fruit defense mechanism against external agents. The different parts of mandarin fruit are rigorously related to relevant species characteristics [
17]. The results of eggplant showed that LEDs light stimulated the accumulation of bioactive compounds and antioxidant activity both in flesh and peel tissues. In addition, it was noticeable that the peel was rich in bioprotective compounds after being irradiated with red + blue LEDs. Whether LED irradiation stimulate phenolic compounds and flavonoid biosynthesis in eggplant fruits is still unclear at present, and needs further analysis.
Many previous studies have reported that blue light increases the production of phenolic compounds. However, the current and previous studies propose that the effect of different light sources and wavelengths on plant secondary metabolite accumulation varies depending on species, cell and tissue type, and plant organ. Further research is necessary to determine the specific intensity and duration of LED lighting that can be used to enhance the production of key substances.
5. Conclusions
We have demonstrated that LED irradiation revealed its effective use in the postharvest of eggplant fruits, in terms of increasing the accumulation of bioactive compounds, including total phenolic, total flavonoid, carotenoid, and anthocyanin contents, as well as antioxidant activity.
Anthocyanin content was increased under red (650–660 nm) and blue (450–460 nm) monochrome of LEDs irradiation. Carotenoid content, however, was higher when accumulated after 48 h using postharvest red LED irradiation. The accumulation of phenolic compounds that exhibited ferric ion-reducing antioxidant power (FRAP assay) and free radical-scavenging activity (DPPH inhibition) were increased by predominantly red + blue light in the eggplant fruits. Major individual phenolic compounds were determined as chlorogenic and gallic acids. Under predominantly red + blue irradiation, chlorogenic acid accumulation was relatively high (up to 2.3- and 3.7-fold, in peel and flesh, respectively) when compared to control treatment. Combinations of red and blue LEDs can be tailored to increase superior antioxidant activity and phenolic content in eggplant, as an effective and convenient tool for increasing the quality of postharvest eggplant fruits.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}