
Determination of Phylloplane Associated Bacteria of Lettuce from a Small-Scale Aquaponic System via 16S rRNA Gene Amplicon Sequence Analysis



Abstract
:1. Introduction
2. Materials and Methods
2.1. System Design
2.2. Lettuce Growth Conditions
2.3. Addition of Probiotics
2.4. Sampling for Water Quality Parameters
2.5. Chlorophyll Fluorescence and Vegetative Growth of Lettuce
2.6. Sample Collection and Isolation of Leaf Epiphytic Bacteria
2.7. DNA Extraction, PCR Amplification of 16SrRNA Gene and Sequencing
2.8. Sequence Analysis
2.9. Analysis of Lettuce Growth and Water Quality
3. Results
3.1. Vegetative Growth of Lettuce and Chlorophyll Fluorescence
3.2. Water Quality Parameters in the Deep-Water Growth Beds
3.3. Sequencing Summary and Diversity of Leaf Epiphytic Bacteria
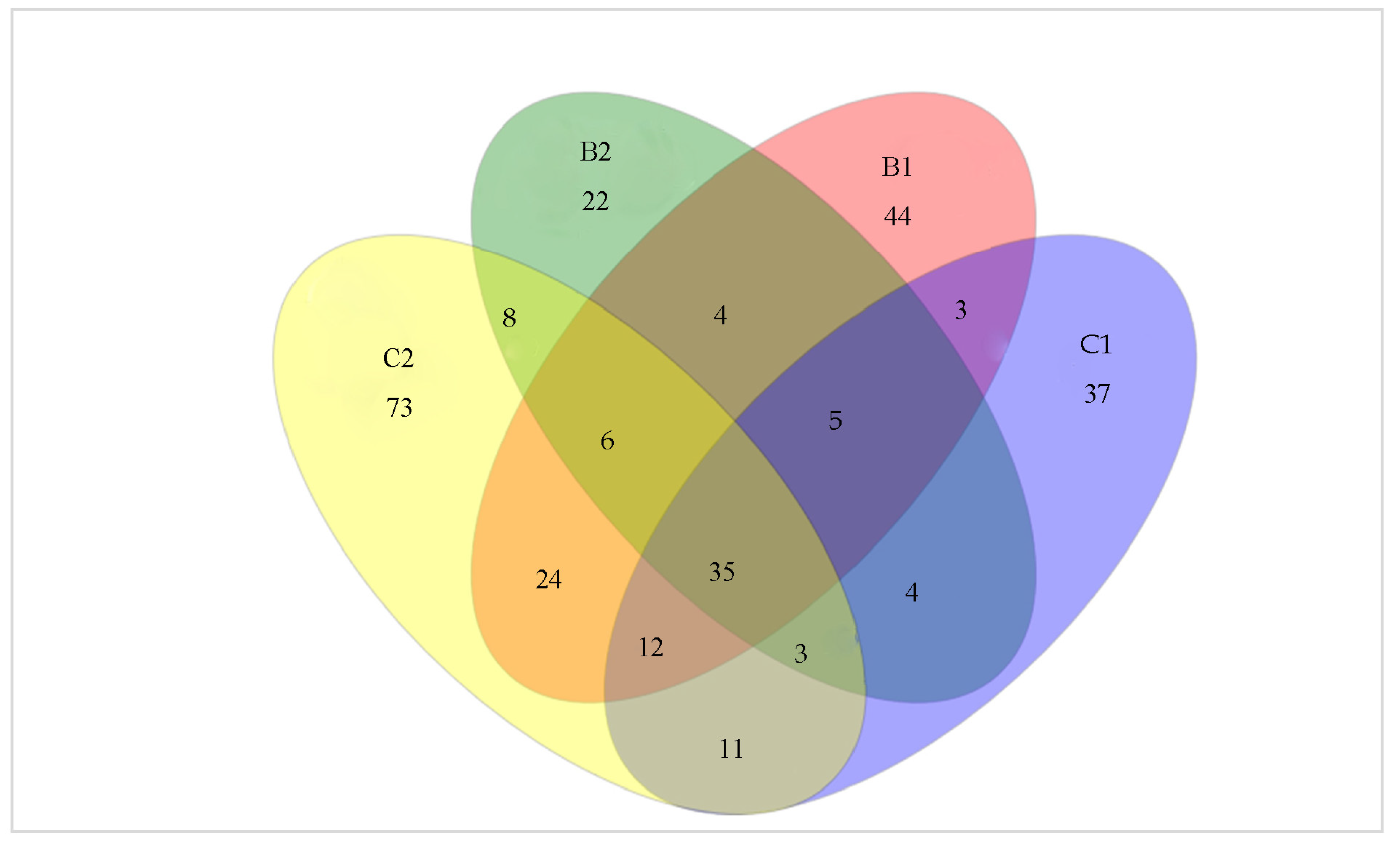
3.4. Shared and Unique Operational Taxonomic Units
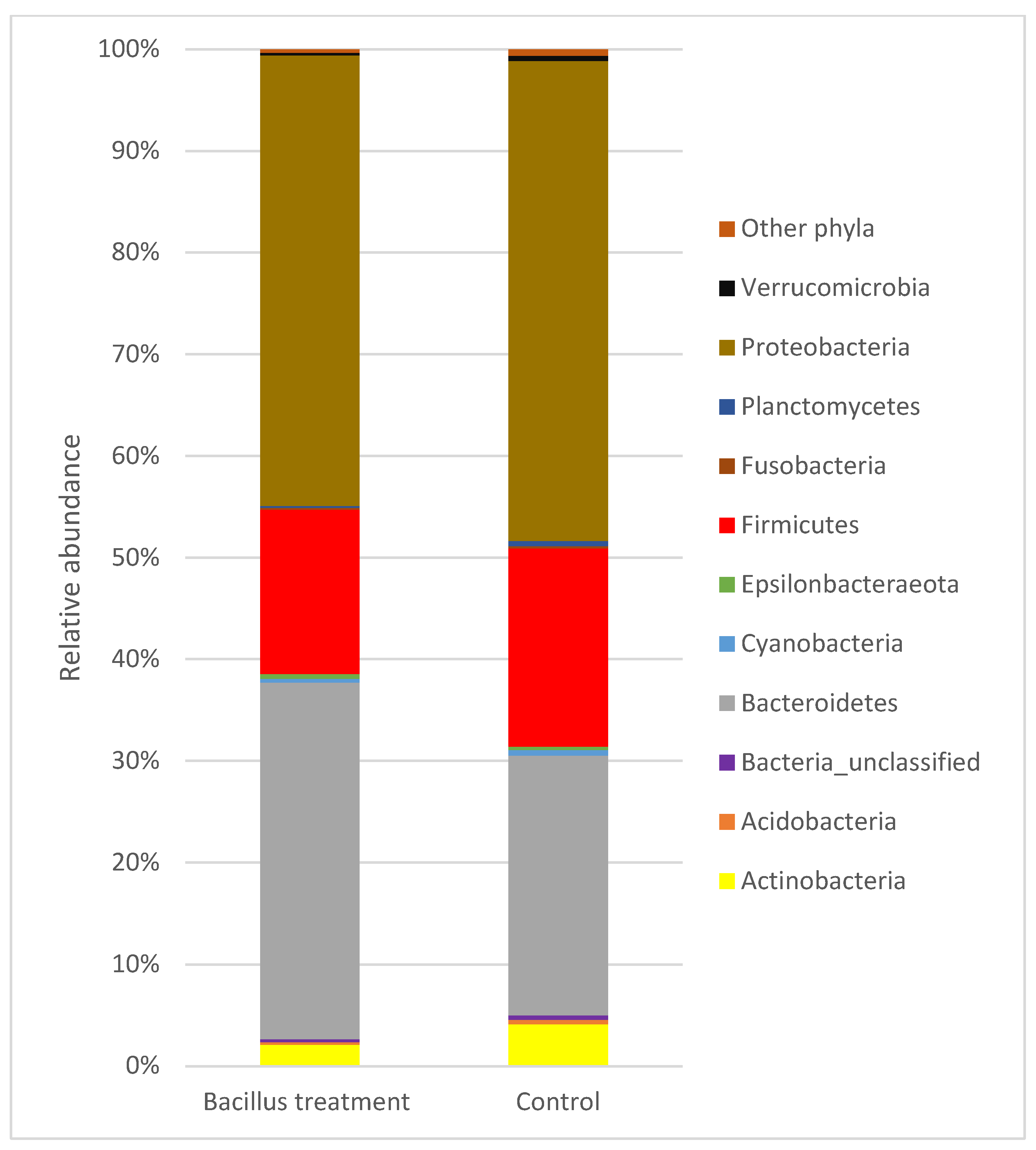
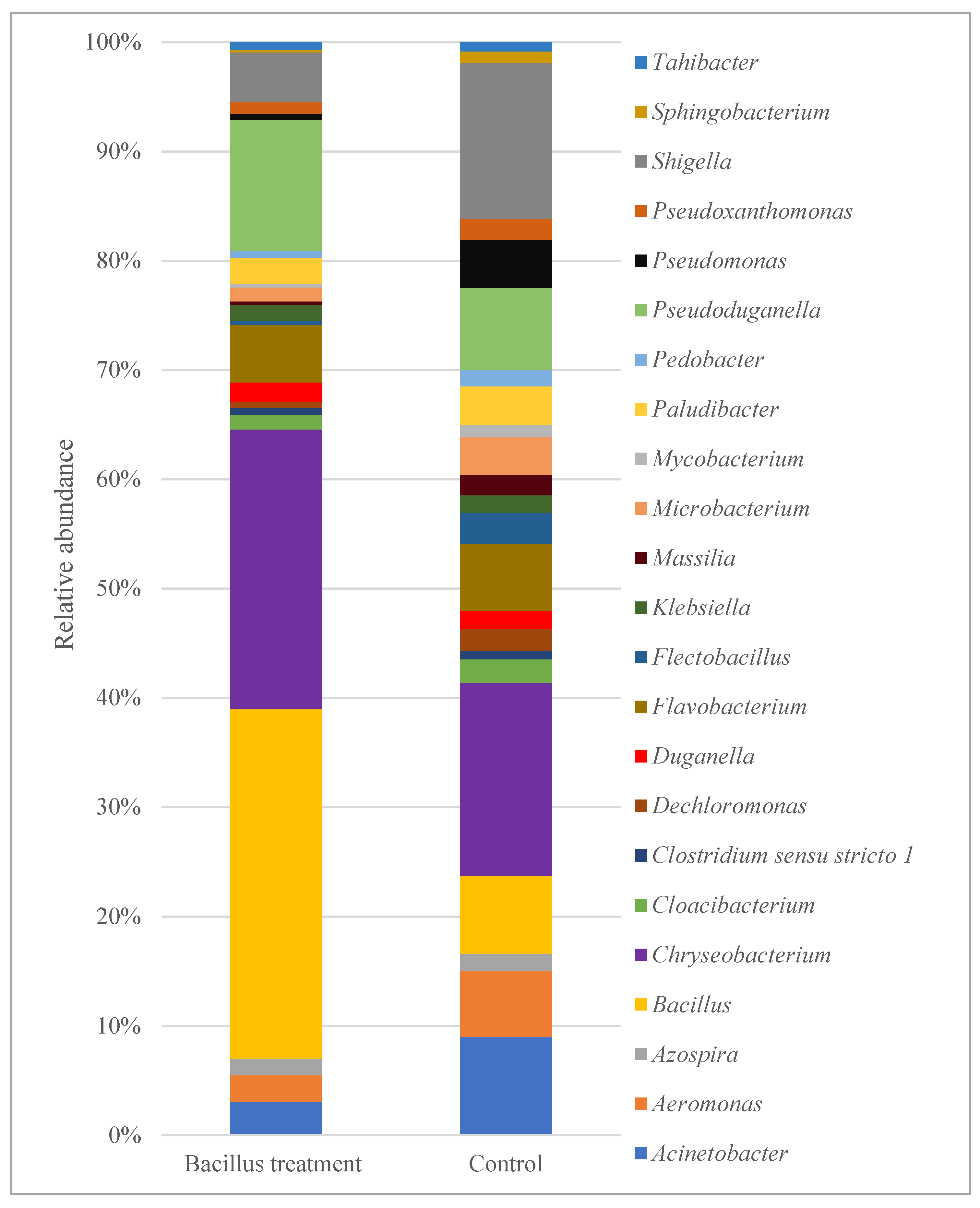
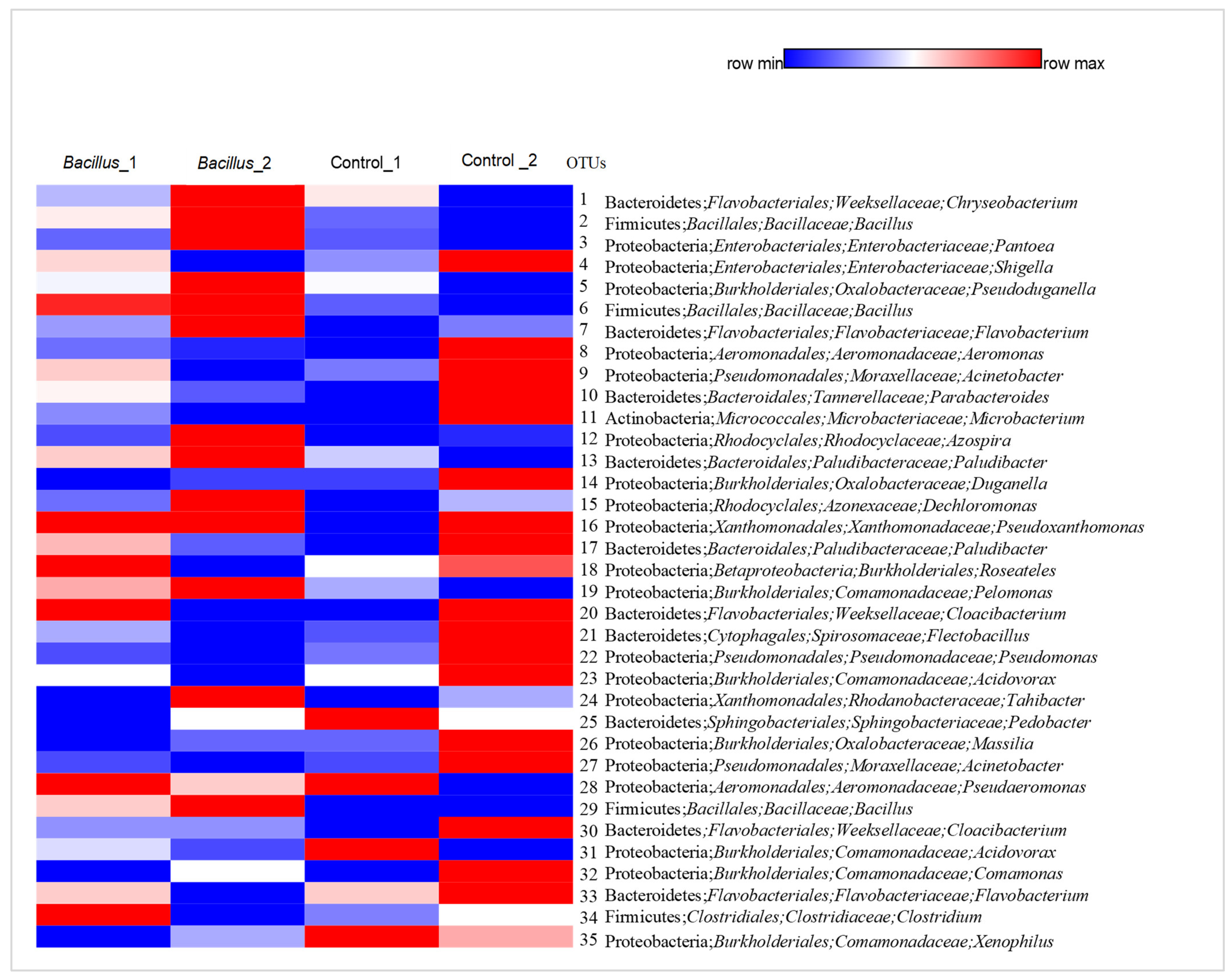
3.5. Overview of Bacterial Taxa Associated with Lettuce Leaf Samples
4. Discussion
4.1. Water Quality Management and Lettuce Growth
4.2. Bacterial Community Composition and Diversity
4.3. Occurrence of Shigella and Aeromonas Species on Leaf Surfaces of Lettuce
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lindow, S.E.; Brandl, M.T. Microbiology of the Phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, T.R.; Marco, M.L. Phyllosphere Microbiota Composition and Microbial Community Transplantation on Lettuce Plants Grown Indoors. mBio 2014, 5, 01564-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Genet. 2012, 10, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Bringel, F.; Couée, I. Pivotal roles of phyllosphere microorganisms at the interface between plant functioning and atmospheric trace gas dynamics. Front. Microbiol. 2015, 6, 486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leveau, J.H.J. Microbial communities in the phyllosphere. In Biology of the Plant Cuticle; Riederer, M., Mueller, C., Eds.; Blackwell Publishing: Oxford, UK, 2006; pp. 334–367. [Google Scholar]
- Sánchez-López, A.S.; González-Chávez, M.D.C.A.; Solís-Domínguez, F.A.; Carrillo-González, R.; Rosas-Saito, G.H. Leaf Epiphytic Bacteria of Plants Colonizing Mine Residues: Possible Exploitation for Remediation of Air Pollutants. Front. Microbiol. 2018, 9, 3028. [Google Scholar] [CrossRef]
- Fürnkranz, M.; Wanek, W.; Richter, A.; Abell, G.; Rasche, F.; Sessitsch, A. Nitrogen fixation by phyllosphere bacteria associated with higher plants and their colonizing epiphytes of a tropical lowland rainforest of Costa Rica. ISME J. 2008, 2, 561–570. [Google Scholar] [CrossRef]
- Lindow, S.E.; Leveau, J.H. Phyllosphere microbiology. Curr. Opin. Biotechnol. 2002, 13, 238–243. [Google Scholar] [CrossRef]
- Rastogi, G.; Coaker, G.L.; Leveau, J.H. New insights into the structure and function of phyllosphere microbiota through high-throughput molecular approaches. FEMS Microbiol. Lett. 2013, 348, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Berlec, A. Novel techniques and findings in the study of plant microbiota: Search for plant probiotics. Plant Sci. 2012, 193–194, 96–102. [Google Scholar] [CrossRef]
- Erickson, M.C.; Webb, C.C.; Diaz-Perez, J.C.; Phatak, S.C.; Silvoy, J.J.; Davey, L.; Payton, A.S.; Liao, J.; Ma, L.; Doyle, M.P. Surface and Internalized Escherichia coli O157:H7 on Field-Grown Spinach and Lettuce Treated with Spray-Contaminated Irrigation Water. J. Food Prot. 2010, 73, 1023–1029. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Deering, A.J.; Kim, H.-J. The Occurrence of Shiga Toxin-Producing E. coli in Aquaponic and Hydroponic Systems. Horticulturae 2020, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Weller, D.; Saylor, L.; Turkon, P. Total Coliform and Generic E. coli Levels, and Salmonella Presence in Eight Experimental Aquaponics and Hydroponics Systems: A Brief Report Highlighting Exploratory Data. Horticulturae 2020, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Yep, B.; Zheng, Y. Aquaponic trends and challenges—A review. J. Clean. Prod. 2019, 228, 1586–1599. [Google Scholar] [CrossRef]
- Stone, B.W.G.; Weingarten, E.A.; Jackson, C.R. The Role of the Phyllosphere Microbiome in Plant Health and Function. Annu. Plant Rev. Online 2018, 1, 533–556. [Google Scholar] [CrossRef]
- Riggio, G.; Jones, S.; Gibson, K. Risk of Human Pathogen Internalization in Leafy Vegetables during Lab-Scale Hydroponic Cultivation. Horticulturae 2019, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Wall, G.L.; Clements, D.P.; Fisk, C.L.; Stoeckel, D.M.; Woods, K.L.; Bihn, E.A. Meeting Report: Key Outcomes from a Collaborative Summit on Agricultural Water Standards for Fresh Produce. Compr. Rev. Food Sci. Food Saf. 2019, 18, 723–737. [Google Scholar] [CrossRef] [Green Version]
- Fox, B.K.; Tamaru, C.S.; Hollyer, J.; Castro, L.F.; Fonseca, J.M.; Jay-russell, M.; Low, T. A preliminary study of microbial water quality related to food safety in recirculating aquaponic fish and vegetable production systems. Food Saf. Technol. 2012, 51, 1–11. [Google Scholar]
- Radhakrishnan, R.; Hashem, A.; Abd_Allah, E.F. Bacillus: A Biological Tool for Crop Improvement through Bio-Molecular Changes in Adverse Environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- da Silva Cerozi, B.; Fitzsimmons, K. Use of Bacillus spp. to enhance phosphorus availability and serve as a plant growth promoter in aquaponics systems. Sci. Hortic. 2016, 211, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Hlordzi, V.; Kuebutornye, F.K.; Afriyie, G.; Abarike, E.D.; Lu, Y.; Chi, S.; Anokyewaa, M.A. The use of Bacillus species in maintenance of water quality in aquaculture: A review. Aquac. Rep. 2020, 18, 100503. [Google Scholar] [CrossRef]
- Hai, N.V. The use of probiotics in aquaculture. J. Appl. Microbiol. 2015, 119, 917–935. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.; Ahmad, I.; Khan, M. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Kasozi, N.; Kaiser, H.; Wilhelmi, B. Effect of Bacillus spp. on Lettuce Growth and Root Associated Bacterial Community in a Small-Scale Aquaponics System. Agronomy 2021, 11, 947. [Google Scholar] [CrossRef]
- Sare, A.R.; Stouvenakers, G.; Eck, M.; Lampens, A.; Goormachtig, S.; Jijakli, M.H.; Massart, S. Standardization of Plant Microbiome Studies: Which Proportion of the Microbiota is Really Harvested? Microorganisms 2020, 8, 342. [Google Scholar] [CrossRef] [Green Version]
- Kasozi, N.; Kaiser, H.; Wilhelmi, B. Metabarcoding Analysis of Bacterial Communities Associated with Media Grow Bed Zones in an Aquaponic System. Int. J. Microbiol. 2020, 2020, 1–10. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- De Winter, J.C.F. Using the Student’s t-test with extremely small sample sizes. Pract. Asses. Res. Eval. 2013, 18, 1–12. [Google Scholar] [CrossRef]
- Somerville, C.; Cohen, M.; Pantanella, E.; Stankus, A.; Lovatelli, A. Small-Scale Aquaponic Food Production. In Integrated Fish and Plant Farming; Technical Paper No. 589; FAO Fisheries and Aquaculture: Rome, Italy, 2014; pp. 90–91. [Google Scholar]
- Yanes, A.R.; Martinez, P.; Ahmad, R. Towards automated aquaponics: A review on monitoring, IoT, and smart systems. J. Clean. Prod. 2020, 263, 121571. [Google Scholar] [CrossRef]
- Kim, B.-R.; Shin, J.; Guevarra, R.B.; Lee, J.H.; Kim, D.W.; Seol, K.-H.; Lee, J.-H.; Kim, H.B.; Isaacson, R.E. Deciphering Diversity Indices for a Better Understanding of Microbial Communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercier, J.; Lindow, S.E. Role of Leaf Surface Sugars in Colonization of Plants by Bacterial Epiphytes. Appl. Environ. Microbiol. 2000, 66, 369–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlechter, R.O.; Miebach, M.; Remus-Emsermann, M.N. Driving factors of epiphytic bacterial communities: A review. J. Adv. Res. 2019, 19, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.J.; Wang, L.L.; Li, Q.; Shang, Q.M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of tomato plants. PLoS ONE 2019, 14, e0223847. [Google Scholar] [CrossRef] [PubMed]
- Montero-Calasanz, M.C.; Göker, M.; Rohde, M.; Spröer, C.; Schumann, P.; Busse, H.J.; Schmid, M.; Klenk, H.P.; Tindall, B.J.; Camacho, M. Chryseobacterium oleae sp. nov., an efficient plant growth promoting bacterium in the rooting induction of olive tree (Olea europaea L.) cuttings and emended descriptions of the genus Chryseobacterium, C. daecheongense, C. gambrini, C. gleum, C. joostei, C. jejuense, C. luteum, C. shigense, C. taiwanense, C. ureilyticum and C. vrystaatense. Syst. Appl. Microbiol. 2014, 37, 342–350. [Google Scholar] [CrossRef]
- Kumar, M.; Charishma, K.; Sahu, K.; Sheoran, N.; Patel, A.; Kundu, A.; Kumar, A. Rice leaf associated Chryseobacterium species: An untapped antagonistic flavobacterium displays volatile mediated suppression of rice blast disease. Biol. Control 2021, 161, 104703. [Google Scholar] [CrossRef]
- De Maayer, P.; Chan, W.-Y.; Blom, J.; Venter, S.N.; Duffy, B.; Smits, T.H.M.; Coutinho, T.A. The large universal Pantoea plasmid LPP-1 plays a major role in biological and ecological diversification. BMC Genom. 2012, 13, 625. [Google Scholar] [CrossRef] [Green Version]
- Pusey, P.L.; Stockwell, V.O.; Reardon, C.L.; Smits, T.H.; Duffy, B. Antibiosis Activity of Pantoea agglomerans Biocontrol Strain E325 Against Erwinia amylovora on Apple Flower Stigmas. Phytopathology 2011, 101, 1234–1241. [Google Scholar] [CrossRef] [Green Version]
- Baldani, J.I.; Rouws, L.; Cruz, L.M.; Olivares, F.L.; Schmid, M.; Hartmann, A. The Family Oxalobacteraceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 943–946. [Google Scholar]
- Kolton, M.; Erlacher, A.; Berg, G.; Cytryn, E. The Flavobacterium Genus in the Plant Holobiont: Ecological, Physiological, and Applicative Insights. In Microbial Models: From Environmental to Industrial Sustainability; Microorganisms for, Sustainability; Castro-Sowinski, S., Ed.; Springer: Singapore, 2016; pp. 189–207. [Google Scholar]
- Kolton, M.; Frenkel, O.; Elad, Y.; Cytryn, E. Potential Role of Flavobacterial Gliding-Motility and Type IX Secretion System Complex in Root Colonization and Plant Defense. Mol. Plant-Microbe Interact. 2014, 27, 1005–1013. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Yan, Z.; Jia, L.; Zhang, S.; Gao, L.; Wei, X.; Mei, Z.; Liu, X. The composition, localization and function of low-temperature-adapted microbial communities involved in methanogenic degradations of cellulose and chitin from Qinghai-Tibetan Plateau wetland soils. J. Appl. Microbiol. 2016, 121, 163–176. [Google Scholar] [CrossRef]
- Wang, X.; Hua, Z.; Mao, H. Influential factors for metal ions removal using extracellular polymeric substances produced by Cloacibacterium normanense. Water Environ. J. 2018, 32, 650–656. [Google Scholar] [CrossRef]
- Kapperud, G.; Rørvik, L.M.; Hasseltvedt, V.; Høiby, E.A.; Iversen, B.G.; Staveland, K.; Johnsen, G.; Leitao, J.; Herikstad, H.; Andersson, Y.; et al. Outbreak of Shigella sonnei infection traced to imported iceberg lettuce. J. Clin. Microbiol. 1995, 33, 609–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, M.B.; Estrada-García, T. Shigella: A Highly Virulent and Elusive Pathogen. Curr. Trop. Med. Rep. 2014, 1, 81–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagamboula, C.F.; Uyttendaele, M.; Debevere, J. Acid tolerance of Shigella sonnei and Shigella flexneri. J. Appl. Microbiol. 2002, 93, 479–486. [Google Scholar] [CrossRef] [Green Version]
- McMahon, M.A.; Wilson, I.G. The occurrence of enteric pathogens and Aeromonas species in organic vegetables. Int J. Food Microbiol. 2001, 70, 155–162. [Google Scholar] [CrossRef]
- Umutoni, N.; Jakobsen, A.N.; Mukhatov, K.; Thomassen, G.M.B.; Karlsen, H.; Mehli, L. Occurrence, diversity and temperature-dependent growth kinetics of Aeromonas spp. in lettuce. Int. J. Food Microbiol. 2020, 335, 108852. [Google Scholar] [CrossRef] [PubMed]
- Chitmanat, C.; Pimpimol, T.; Chaibu, P. Investigation of Bacteria and Fish Pathogenic Bacteria Found in Freshwater Aquaponic System. J. Agric. Sci. 2015, 7, 254–259. [Google Scholar] [CrossRef] [Green Version]
- Schmautz, Z.; Graber, A.; Jaenicke, S.; Goesmann, A.; Junge, R.; Smits, T.H.M. Microbial diversity in different compartments of an aquaponics system. Arch. Microbiol. 2017, 199, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Cipriano, R.L. Aeromonas Hydrophila and Motile Aeromonad Septicemias of Fish; US Fish & Wildlife Publications: Washington, DC, USA, 1984; Volume 68, pp. 1–23. [Google Scholar]
- Lehane, L.; Rawlin, G.T. Topically acquired bacterial zoonoses from fish: A review. Med. J. Aust. 2000, 173, 256–259. [Google Scholar] [CrossRef]
- Tekedar, H.C.; Karsi, A.; Akgul, A.; Kalindamar, S.; Waldbieser, G.C.; Sonstegard, T.; Schroeder, S.G.; Lawrence, M.L. Complete Genome Sequence of Fish Pathogen Aeromonas hydrophila AL06-06. Genome Announc. 2015, 3, e00368-15. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.I.; Cavaco, A.; Gouveia, J.; Novais, M.; Nogueira, P.; Pedroso, L.; Ferreira, M.A.S.S. Evaluation of minimally processed salads commercialized in Portugal. Food Control 2012, 23, 275–281. [Google Scholar] [CrossRef]
- Blake, C.; Christensen, M.N.; Kovács, Á.T. Molecular Aspects of Plant Growth Promotion and Protection by Bacillus subtilis. Mol. Plant-Microbe Interact. 2021, 34, 15–25. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Bacillus Treatment | Control | p-Value |
|---|---|---|---|
| Initial plant mass (g plant−1) | 6.18 ± 0.02 | 6.21 ± 0.04 | 0.619 |
| Shoot fresh weight (g plant−1) | 45.03 ± 0.06 | 39.07 ± 0.91 | 0.022 * |
| Root fresh weight (g plant−1) | 2.42 ± 0.005 | 2.00 ± 0.04 | 0.011 * |
| Height gain (cm plant−1) | 26.66 ± 0.48 | 21.60 ± 0.38 | 0.014 * |
| Leaf number (plant−1) | 19.60 ± 0.22 | 15.0 ± 0.19 | 0.004 * |
| Absolute growth rate (g day−1) | 1.38 ± 0.005 | 1.16 ± 0.03 | 0.019 * |
| Fv/Fm | 0.82 ± 0.05 | 0.72 ± 0.01 | 0.011 * |
| CCI | 2.36 ± 0.05 | 2.25 ± 0.05 | 0.260 |
| Shoot dry weight (g plant−1) | 2.07 ± 0.07 | 1.72 ± 0.01 | 0.043 * |
| Root dry weight (g plant−1) | 0.27 ± 0.005 | 0.17 ± 0.01 | 0.016 * |
| Variables | Bacillus Treatment | Control |
|---|---|---|
| Temperature (°C) | 25.94 (24.96–26.50) | 25.94 (25.04–26.50) |
| DO (mg L−1) | 6.72 (6.53–6.83) | 6.79 (6.67–6.90) |
| TDS (mg L−1) | 741 (661–839) | 835 (734–931) |
| pH | 6.87 (6.74–6.97) | 6.79 (6.68–6.90) |
| EC (ms cm−1) | 1.05 (1.05–1.19) | 1.20 (1.03–1.35) |
| Nitrate (mg L−1) | 33.25 (25.27–43.60) | 19.73 (14.50–27.37) |
| Phosphate (mg L−1) | 2.92 (2.34–4.43) | 1.60 (1.14–2.39) |
| Treatment | Number of Raw Sequences | Number of Sequences before Chimeras | Number of Sequences after Chimeras | Eukaryotes and Unknown Reads | Number of Reads after Screening and Filtering |
|---|---|---|---|---|---|
| Bacillus treatment | |||||
| Sample B1 | 48,788 | 35,348 | 34,874 | 24,544 | 622 |
| Sample B2 | 47,791 | 36,016 | 33,784 | 17,175 | 6349 |
| Control | |||||
| Sample C1 | 63,098 | 41,061 | 40,336 | 16,544 | 1119 |
| Sample C2 | 41,566 | 35,042 | 34,494 | 29,884 | 749 |
| Coverage and Indices | Treatment | p-Value | |
|---|---|---|---|
| Bacillus | Control | ||
| InvSimpson | 13.53 (7.95–19.12) | 23.25 (10.34–36.15) | 0.561 |
| Observed richness | 110 (87–133) | 141 (110–172) | 0.506 |
| Good’s coverage (%) | 90.0 (87.62–92.44) | 88.9 (87.29–90.51) | 0.735 |
| Chao1 | 218.65 (177.08–260.22) | 255.41 (195.55–255.27) | 0.907 |
| Shannon | 3.32 (2.94–3.70) | 3.81 (3.23–4.39) | 0.554 |
| Bacterial OTUs | Related Species (NCBI-Blast) | Similarity (%) | Accession |
|---|---|---|---|
| OTU1 | Chryseobacterium rhizoplanae strain JM-534 | 98.27% | NR_134711.1 |
| OTU2 | Bacillus australimaris strain MCCC 1A05787 | 98.72% | NR_148787.1 |
| OTU3 | Pantoea rwandensis strain LMG 26275 | 99.14% | NR_118121.1 |
| OTU4 | Shigella sonnei strain CECT 4887 | 98.28% | NR_104826.1 |
| OTU5 | Pseudoduganella eburnea strain 10R 5-21 | 98.28% | NR_159256.1 |
| OTU6 | Bacillus haynesii strain NRRL B-41327 | 98.50% | NR_157609.1 |
| OTU7 | Flavobacterium lindanitolerans strain IP-10 | 98.26% | NR_044208.1 |
| OTU8 | Aeromonas hydrophila strain DSM 30187 | 99.36% | NR_119190.1 |
| OTU9 | Acinetobacter vivianii strain NIPH 2168 | 98.71% | NR_148847.1 |
| OTU10 | Parabacteroides chartae strain NS31-3 | 98.92% | NR_109439.1 |
| OTU11 | Microbacterium laevaniformans strain DSM 20140 | 98.72% | NR_044935.1 |
| OTU12 | Azospira oryzae PS | 98.72% | NR_074103.1 |
| OTU13 | Paludibacter propionicigenes WB4 | 97.41% | NR_074577.1 |
| OTU14 | Duganella sacchari strain Sac-22 | 98.28% | NR_108216.1 |
| OTU15 | Dechloromonas hortensis strain MA-1 | 98.72% | NR_042819.1 |
| OTU16 | Pseudoxanthomonas mexicana strain NBRC 101034 | 97.43% | NR_113973.1 |
| OTU17 | Paludibacter propionicigenes WB4 | 97.41% | NR_074577.1 |
| OTU18 | Roseateles depolymerans strain 61A | 96.56% | NR_111995.1 |
| OTU19 | Pelomonas saccharophila strain NBRC 103037 | 98.71% | NR_114189.1 |
| OTU20 | Cloacibacterium normanense strain NRS1 | 98.92% | NR_042187.1 |
| OTU21 | Flectobacillus roseus strain GFA-11 | 98.70% | NR_116312.1 |
| OTU22 | Pseudomonas plecoglossicida strain NBRC 103162 | 99.36% | NR_114226.1 |
| OTU23 | Acidovorax temperans strain PHL | 99.35% | NR_028715.1 |
| OTU24 | Tahibacter aquaticus strain PYM5-11 | 97.22% | NR_115098.1 |
| OTU25 | Pedobacter glacialis strain 8-24 | 98.70% | NR_134125.1 |
| OTU26 | Massilia alkalitolerans DSM 17462 strain YIM 31775 | 98.92% | NR_043094.1 |
| OTU27 | Acinetobacter lwoffii strain JCM 6840 | 99.14% | NR_113346.1 |
| OTU28 | Pseudaeromonas sharmana strain GPTSA-6 | 98.28% | NR_043470.1 |
| OTU29 | Bacillus haynesii strain NRRL B-41327 | 97.64% | NR_157609.1 |
| OTU30 | Cloacibacterium rupense strain NBRC 104931 | 98.92% | NR_114274.1 |
| OTU31 | Acidovorax monticola strain K-4-16 | 97.63% | NR_164911.1 |
| OTU32 | Comamonas composti strain YY287 | 99.57% | NR_044039.1 |
| OTU33 | Flavobacterium notoginsengisoli strain SYP-B540 | 98.92% | NR_145940.1 |
| OTU34 | Clostridium beijerinckii strain JCM 1390 | 99.57% | NR_113388.1 |
| OTU35 | Xenophilus aerolatus strain 5516S-2 | 99.35% | NR_116099.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasozi, N.; Kaiser, H.; Wilhelmi, B. Determination of Phylloplane Associated Bacteria of Lettuce from a Small-Scale Aquaponic System via 16S rRNA Gene Amplicon Sequence Analysis. Horticulturae 2022, 8, 151. https://doi.org/10.3390/horticulturae8020151
Kasozi N, Kaiser H, Wilhelmi B. Determination of Phylloplane Associated Bacteria of Lettuce from a Small-Scale Aquaponic System via 16S rRNA Gene Amplicon Sequence Analysis. Horticulturae. 2022; 8(2):151. https://doi.org/10.3390/horticulturae8020151
Chicago/Turabian StyleKasozi, Nasser, Horst Kaiser, and Brendan Wilhelmi. 2022. "Determination of Phylloplane Associated Bacteria of Lettuce from a Small-Scale Aquaponic System via 16S rRNA Gene Amplicon Sequence Analysis" Horticulturae 8, no. 2: 151. https://doi.org/10.3390/horticulturae8020151
APA StyleKasozi, N., Kaiser, H., & Wilhelmi, B. (2022). Determination of Phylloplane Associated Bacteria of Lettuce from a Small-Scale Aquaponic System via 16S rRNA Gene Amplicon Sequence Analysis. Horticulturae, 8(2), 151. https://doi.org/10.3390/horticulturae8020151