
Effects of Biogenic ZnO Nanoparticles on Growth, Physiological, Biochemical Traits and Antioxidants on Olive Tree In Vitro

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions of Lemna Minor and Synthesis and Characterization of Biogenic ZnO-NPs
2.2. Plant Material, ZnO-NPs Treatment, and Growing Conditions
2.3. Growth Parameters
2.4. Determinations on In Vitro Proliferated Shoots of Chlorophylls, Carotenoid, Soluble Protein, Anthocyanins, Total Phenol Content (TPC) and Radical Scavenging Rate (DPPH)
2.5. Statistical Analysis
3. Results
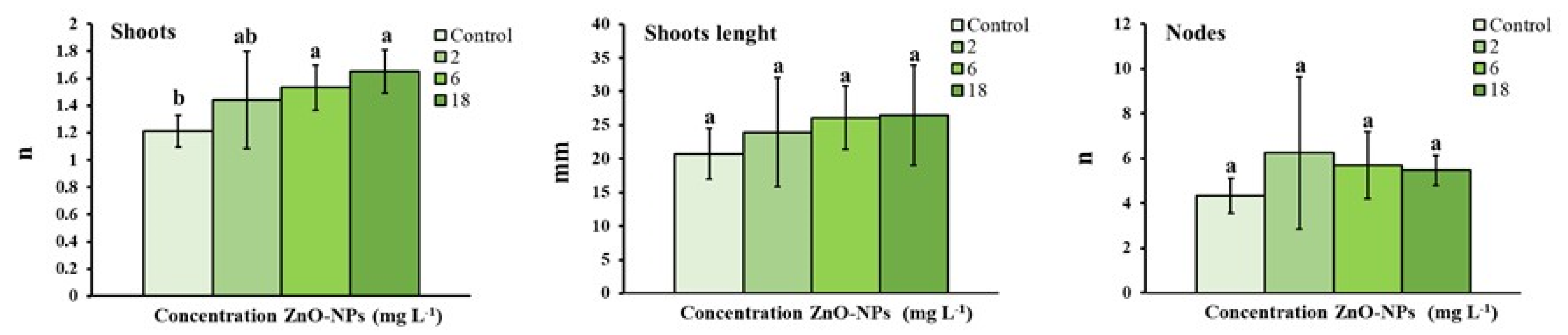
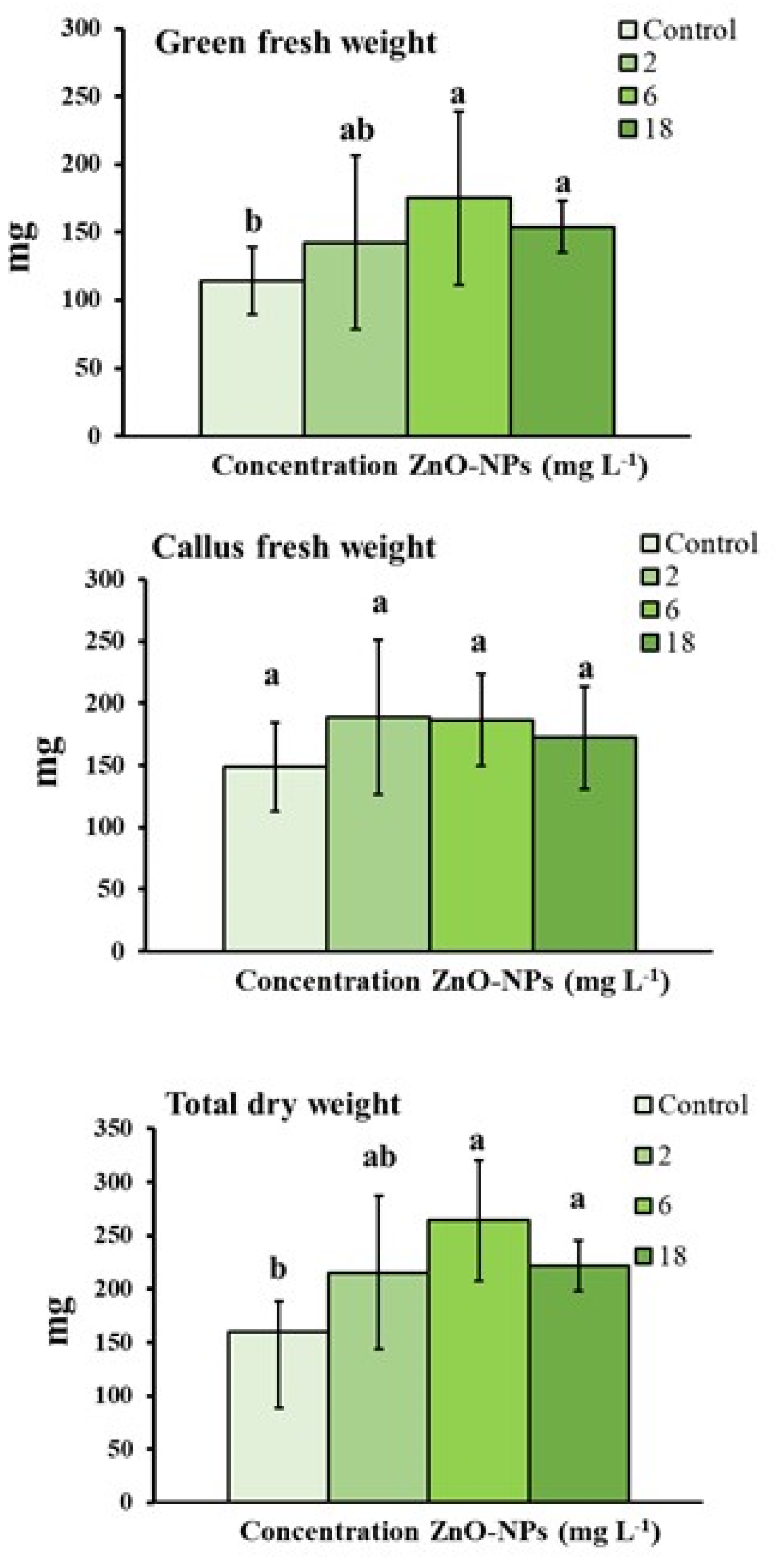
3.1. Growth Parameters
3.2. Effect of ZnO-NPs on Pigment and Soluble Protein
3.3. Effects of ZnO-NPs on Carotenoids, Anthocyanins, Total Phenol Content (TPC) and Radical Scavenging Rate (DPPH)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sturikova, H.; Krystofova, O.; Huska, D.; Adam, V. Zinc, Zinc Nanoparticles and Plants. J. Hazard. Mater. 2018, 349, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.M.; Guerinot, M.L. Facing the Challenges of Cu, Fe and Zn Homeostasis in Plants. Nat. Chem. Biol. 2009, 5, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hänsch, R.; Mendel, R.R. Physiological Functions of Mineral Micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Physiol. Metab. 2009, 12, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Patni, B.; Shankhdhar, D.; Shankhdhar, S.C. Zinc—An Indispensable Micronutrient. Physiol. Mol. Biol. Plants 2013, 19, 11–20. [Google Scholar] [CrossRef]
- Ren, F.-C.; Liu, T.-C.; Liu, H.-Q.; Hu, B.-Y. Influence of Zinc on the Growth, Distribution of Elements, and Metabolism of One-Year Old American Ginseng Plants. J. Plant Nutr. 1993, 16, 393–405. [Google Scholar] [CrossRef]
- Del Buono, D.; Luzi, F.; Puglia, D. Lignin Nanoparticles: A Promising Tool to Improve Maize Physiological, Biochemical, and Chemical Traits. Nanomaterials 2021, 11, 846. [Google Scholar] [CrossRef]
- Del Buono, D.; Di Michele, A.; Costantino, F.; Trevisan, M.; Lucini, L. Biogenic Zno Nanoparticles Synthesized Using a Novel Plant Extract: Application to Enhance Physiological and Biochemical Traits in Maize. Nanomaterials 2021, 11, 1270. [Google Scholar] [CrossRef]
- Khot, L.R.; Sankaran, S.; Maja, J.M.; Ehsani, R.; Schuster, E.W. Applications of Nanomaterials in Agricultural Production and Crop Protection: A Review. Crop Prot. 2012, 35, 64–70. [Google Scholar] [CrossRef]
- Matussin, S.; Harunsani, M.H.; Tan, A.L.; Khan, M.M. Plant-Extract-Mediated SnO2 Nanoparticles: Synthesis and Applications. ACS Sustain. Chem. Eng. 2020, 8, 3040–3054. [Google Scholar] [CrossRef]
- Nayantara; Kaur, P. Biosynthesis of Nanoparticles Using Eco-Friendly Factories and Their Role in Plant Pathogenicity: A Review. Biotechnol. Res. Innov. 2018, 2, 63–73. [Google Scholar] [CrossRef]
- Karnan, T.; Selvakumar, S.A.S. Biosynthesis of ZnO Nanoparticles Using Rambutan (Nephelium Lappaceum, L.) Peel Extract and Their Photocatalytic Activity on Methyl Orange Dye. J. Mol. Struct. 2016, 1125, 358–365. [Google Scholar] [CrossRef]
- Hessien, M.; Da’na, E.; Taha, A. Phytoextract Assisted Hydrothermal Synthesis of ZnO–NiO Nanocomposites Using Neem Leaves Extract. Ceram. Int. 2021, 47, 811–816. [Google Scholar] [CrossRef]
- Jangannanavar, V.D.; Patil, M.K.; Chougala, L.S.; Inamdar, S.R.; Goudar, K.M. Biogenic Synthesis and Characterization of ZnO Nanoparticles from Aloe Barbadensis Miller Leaf Extract. Macromol. Symp. 2021, 400, 2100177. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Divya, M.; Vaseeharan, B.; Ranjan, S.; Kalaiselvi, V.; Dasgupta, N.; Chen, J.; Durán-Lara, E.F. Biogenic Preparation and Characterization of ZnO Nanoparticles from Natural Polysaccharide Azadirachta Indica.L. (Neem Gum) and Its Clinical Implications. J. Clust. Sci. 2021, 32, 983–993. [Google Scholar] [CrossRef]
- Ahmed, S.; Annu; Chaudhry, S.A.; Ikram, S. A Review on Biogenic Synthesis of ZnO Nanoparticles Using Plant Extracts and Microbes: A Prospect towards Green Chemistry. J. Photochem. Photobiol. B 2017, 166, 272–284. [Google Scholar] [CrossRef]
- El-Mahdy, M.T.; Elazab, D.S. Impact of Zinc Oxide Nanoparticles on Pomegranate Growth under In Vitro Conditions. Russ. J. Plant Physiol. 2020, 67, 162–167. [Google Scholar] [CrossRef]
- Sun, L.; Wang, Y.; Wang, R.; Wang, R.; Zhang, P.; Ju, Q.; Xu, J. Physiological, Transcriptomic, and Metabolomic Analyses Reveal Zinc Oxide Nanoparticles Modulate Plant Growth in Tomato. Environ. Sci. Nano 2020, 7, 3587–3604. [Google Scholar] [CrossRef]
- Pandey, A.C.; Sanjay, S.S.; Yadav, R.S. Application of ZnO Nanoparticles in Influencing the Growth Rate of Cicer Arietinum. J. Exp. Nanosci. 2010, 5, 488–497. [Google Scholar] [CrossRef]
- Lin, D.; Xing, B. Root Uptake and Phytotoxicity of ZnO Nanoparticles. Environ. Sci. Technol. 2008, 42, 5580–5585. [Google Scholar] [CrossRef] [PubMed]
- Prasad, T.N.V.K.V.; Sudhakar, P.; Sreenivasulu, Y.; Latha, P.; Munaswamy, V.; Raja Reddy, K.; Sreeprasad, T.S.; Sajanlal, P.R.; Pradeep, T. Effect of Nanoscale Zinc Oxide Particles on the Germination, Growth and Yield of Peanut. J. Plant Nutr. 2012, 35, 905–927. [Google Scholar] [CrossRef]
- Sheoran, P.; Grewal, S.; Kumari, S.; Goel, S. Enhancement of Growth and Yield, Leaching Reduction in Triticum Aestivum Using Biogenic Synthesized Zinc Oxide Nanofertilizer. Biocatal. Agric. Biotechnol. 2021, 32, 101938. [Google Scholar] [CrossRef]
- Ghosh, M.; Jana, A.; Sinha, S.; Jothiramajayam, M.; Nag, A.; Chakraborty, A.; Mukherjee, A.; Mukherjee, A. Effects of ZnO Nanoparticles in Plants: Cytotoxicity, Genotoxicity, Deregulation of Antioxidant Defenses, and Cell-Cycle Arrest. Mutat. Res.—Genet. Toxicol. Environ. Mutagen. 2016, 807, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Mishra, R.K.; Singh, S.; Singh, S.; Vishwakarma, K.; Sharma, S.; Singh, V.P.; Singh, P.K.; Prasad, S.M.; Dubey, N.K.; et al. Nitric Oxide Ameliorates Zinc Oxide Nanoparticles Phytotoxicity in Wheat Seedlings: Implication of the Ascorbate–Glutathione Cycle. Front. Plant Sci. 2017, 8, 1. [Google Scholar] [CrossRef]
- Kumar, V.; Parvatam, G.; Ravishankar, G.A. AgNO3—A Potential Regulator of Ethylene Activity and Plant Growth Modulator. Electron. J. Biotechnol. 2009, 12, 8–9. [Google Scholar] [CrossRef] [Green Version]
- Aghdaei, M.; Salehi, H.; Sarmast, M.K. Effects of Silver Nanoparticles on Tecomella Undulata (Roxb.) Seem. Micropropagation. Adv. Hortic. Sci. 2012, 26, 21–24. [Google Scholar]
- Wang, P.; Lombi, E.; Zhao, F.-J.; Kopittke, P.M. Nanotechnology: A New Opportunity in Plant Sciences. Trends Plant Sci. 2016, 21, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Spinoso-Castillo, J.L.; Chavez-Santoscoy, R.A.; Bogdanchikova, N.; Pérez-Sato, J.A.; Morales-Ramos, V.; Bello-Bello, J.J. Antimicrobial and Hormetic Effects of Silver Nanoparticles on in Vitro Regeneration of Vanilla (Vanilla Planifolia Jacks. Ex Andrews) Using a Temporary Immersion System. Plant Cell Tissue Organ Cult. PCTOC 2017, 129, 195–207. [Google Scholar] [CrossRef]
- Asgari-Targhi, G.; Iranbakhsh, A.; Ardebili, Z.O. Potential Benefits and Phytotoxicity of Bulk and Nano-Chitosan on the Growth, Morphogenesis, Physiology, and Micropropagation of Capsicum Annuum. Plant Physiol. Biochem. 2018, 127, 393–402. [Google Scholar] [CrossRef]
- Regni, L.; Micheli, M.; Del Pino, A.M.; Palmerini, C.A.; D’Amato, R.; Facchin, S.L.; Famiani, F.; Peruzzi, A.; Mairech, H.; Proietti, P. The First Evidence of the Beneficial Effects of Se-Supplementation on in Vitro Cultivated Olive Tree Explants. Plants 2021, 10, 1630. [Google Scholar] [CrossRef]
- Balla, I.; Mansvelt, L. Micropropagation of Peach Rootstocks and Cultivars. Methods Mol. Biol. 2013, 11013, 148. [Google Scholar] [CrossRef]
- Dobránszki, J.; Teixeira da Silva, J.A. Micropropagation of Apple—A Review. Biotechnol. Adv. 2010, 28, 462–488. [Google Scholar] [CrossRef]
- Kavand, S.; Kermani, M.J.; Haghnazari, A.; Khosravi, P.; Azimi, M.R. Micropropagation and Medium-Term Conservation of Rosa Pulverulenta. Acta Sci.—Agron. 2011, 33, 297–301. [Google Scholar] [CrossRef]
- Rugini, E. In Vitro Propagation of Some Olive (Olea Europaea Sativa L.) Cultivars with Different Root-Ability, and Medium Development Using Analytical Data from Developing Shoots and Embryos. Sci. Hortic. 1984, 24, 123–134. [Google Scholar] [CrossRef]
- Venkatachalam, P.; Priyanka, N.; Manikandan, K.; Ganeshbabu, I.; Indiraarulselvi, P.; Geetha, N.; Muralikrishna, K.; Bhattacharya, R.C.; Tiwari, M.; Sharma, N.; et al. Enhanced Plant Growth Promoting Role of Phycomolecules Coated Zinc Oxide Nanoparticles with P Supplementation in Cotton (Gossypium Hirsutum L.). Eff. Nanomater. Plants 2017, 110, 118–127. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- EL Arroussi, H.; Benhima, R.; Elbaouchi, A.; Sijilmassi, B.; EL Mernissi, N.; Aafsar, A.; Meftah-Kadmiri, I.; Bendaou, N.; Smouni, A. Dunaliella Salina Exopolysaccharides: A Promising Biostimulant for Salt Stress Tolerance in Tomato (Solanum Lycopersicum). J. Appl. Phycol. 2018, 30, 2929–2941. [Google Scholar] [CrossRef]
- Paradiković, N.; Vinković, T.; Vinković Vrček, I.; Žuntar, I.; Bojić, M.; Medić-Šarić, M. Effect of Natural Biostimulants on Yield and Nutritional Quality: An Example of Sweet Yellow Pepper (Capsicum Annuum L.) Plants. J. Sci. Food Agric. 2011, 91, 2146–2152. [Google Scholar] [CrossRef] [PubMed]
- Sheikhalipour, M.; Esmaielpour, B.; Behnamian, M.; Gohari, G.; Giglou, M.T.; Vachova, P.; Rastogi, A.; Brestic, M.; Skalicky, M. Chitosan–Selenium Nanoparticle (Cs–Se Np) Foliar Spray Alleviates Salt Stress in Bitter Melon. Nanomaterials 2021, 11, 684. [Google Scholar] [CrossRef] [PubMed]
- Lenth, R.V. Least-Squares Means: The R Package Lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Peixe, A.; Raposo, A.; Lourenço, R.; Cardoso, H.; Macedo, E. Coconut Water and BAP Successfully Replaced Zeatin in Olive (Olea Europaea L.) Micropropagation. Sci. Hortic. 2007, 113, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Micheli, M.; da Silva, D.F.; Farinelli, D.; Agate, G.; Pio, R.; Famiani, F. Neem Oil Used as a “Complex Mixture” to Improve in Vitro Shoot Proliferation in Olive. HortScience 2018, 53, 531–534. [Google Scholar] [CrossRef] [Green Version]
- Lambardi, M.; Ozudogru, E.A.; Roncasaglia, R. In Vitro Propagation of Olive (Olea Europaea L.) by Nodal Segmentation of Elongated Shoots. In Protocols for Micropropagation of Selected Economically-Important Horticultural Plants; Lambardi, M., Ozudogru, E.A., Jain, S.M., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 33–44. ISBN 978-1-62703-074-8. [Google Scholar] [CrossRef]
- García, J.L.; Troncoso, J.; Sarmiento, R.; Troncoso, A. Influence of Carbon Source and Concentration on the in Vitro Development of Olive Zygotic Embryos and Explants Raised from Them. Plant Cell Tissue Organ Cult. 2002, 69, 95–100. [Google Scholar] [CrossRef]
- Javed, R.; Usman, M.; Yücesan, B.; Zia, M.; Gürel, E. Effect of Zinc Oxide (ZnO) Nanoparticles on Physiology and Steviol Glycosides Production in Micropropagated Shoots of Stevia Rebaudiana Bertoni. Plant Physiol. Biochem. 2017, 110, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Zafar, H.; Ali, A.; Ali, J.S.; Haq, I.U.; Zia, M. Effect of ZnO Nanoparticles on Brassica Nigra Seedlings and Stem Explants: Growth Dynamics and Antioxidative Response. Front. Plant Sci. 2016, 7, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.; Raja, N.; Mashwani, Z.-R.; Iqbal, M.; Ejaz, M.; Yasmeen, F. In Vitro Germination and Biochemical Profiling of Citrus Reticulata in Response to Green Synthesized Zinc and Copper Nanoparticles. IET Nanobiotechnol. 2017, 11, 790–796. [Google Scholar] [CrossRef]
- Ali, A.; Ahmad, T.; Abbasi, N.A.; Hafiz, I.A. Effect of Different Media and Growth Regulators on in Vitro Shoot Proliferation of Olive Cultivar “Moraiolo”. Pak. J. Bot. 2009, 41, 783–795. [Google Scholar]
- Mendoza-De Gyves, E.; Rosana Mira, F.; Ruiu, F.; Rugini, E. Stimulation of Node and Lateral Shoot Formation in Micropropagation of Olive (Olea Europaea L.) by Using Dikegulac. Plant Cell Tissue Organ Cult. 2008, 92, 233–238. [Google Scholar] [CrossRef]
- Sonobe, R.; Yamashita, H.; Mihara, H.; Morita, A.; Ikka, T. Estimation of Leaf Chlorophyll a, b and Carotenoid Contents and Their Ratios Using Hyperspectral Reflectance. Remote Sens. 2020, 12, 3265. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Desoky, E.-S.M.; Saad, A.M.; Eid, R.S.M.; Selem, E.; Elrys, A.S. Biological Silicon Nanoparticles Improve Phaseolus Vulgaris L. Yield and Minimize Its Contaminant Contents on a Heavy Metals-Contaminated Saline Soil. J. Environ. Sci. China 2021, 106, 1–14. [Google Scholar] [CrossRef]
- Manzoor, N.; Ahmed, T.; Noman, M.; Shahid, M.; Nazir, M.M.; Ali, L.; Alnusaire, T.S.; Li, B.; Schulin, R.; Wang, G. Iron Oxide Nanoparticles Ameliorated the Cadmium and Salinity Stresses in Wheat Plants, Facilitating Photosynthetic Pigments and Restricting Cadmium Uptake. Sci. Total Environ. 2021, 769, 145221. [Google Scholar] [CrossRef]
- Singh, P.; Arif, Y.; Siddiqui, H.; Sami, F.; Zaidi, R.; Azam, A.; Alam, P.; Hayat, S. Nanoparticles Enhances the Salinity Toxicity Tolerance in Linum Usitatissimum L. by Modulating the Antioxidative Enzymes, Photosynthetic Efficiency, Redox Status and Cellular Damage. Ecotoxicol. Environ. Saf. 2021, 213, 112020. [Google Scholar] [CrossRef]
- Salama, D.M.; Osman, S.A.; Abd El-Aziz, M.E.; Abd Elwahed, M.S.A.; Shaaban, E.A. Effect of Zinc Oxide Nanoparticles on the Growth, Genomic DNA, Production and the Quality of Common Dry Bean (Phaseolus Vulgaris). Biocatal. Agric. Biotechnol. 2019, 18, 101083. [Google Scholar] [CrossRef]
- Mingyu, S.; Xiao, W.; Chao, L.; Chunxiang, Q.; Xiaoqing, L.; Liang, C.; Hao, H.; Fashui, H. Promotion of Energy Transfer and Oxygen Evolution in Spinach Photosystem Ii by Nano-Anatase TiO2. Biol. Trace Elem. Res. 2007, 119, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Al-Mayahi, A.M.W. The Effect of Humic Acid (HA) and Zinc Oxide Nanoparticles (ZnO-NPS) on in Vitro Regeneration of Date Palm (Phoenix Dactylifera L.) Cv. Quntar. Plant Cell Tissue Organ Cult. PCTOC 2021, 145, 445–456. [Google Scholar] [CrossRef]
- El-Badri, A.M.A.; Batool, M.; Mohamed, I.A.A.; Khatab, A.; Sherif, A.; Wang, Z.; Salah, A.; Nishawy, E.; Ayaad, M.; Kuai, J.; et al. Modulation of Salinity Impact on Early Seedling Stage via Nano-Priming Application of Zinc Oxide on Rapeseed (Brassica Napus L.). Plant Physiol. Biochem. 2021, 166, 376–392. [Google Scholar] [CrossRef]
- Sohail; Kamran, K.; Kemmerling, B.; Shutaywi, M.; Mashwani, Z.U.R. Nano Zinc Elicited Biochemical Characterization, Nutritional Assessment, Antioxidant Enzymes and Fatty Acid Profiling of Rapeseed. PLoS ONE 2020, 15, e0241568. [Google Scholar] [CrossRef]
- Abdel-Wahab, D.A.; Othman, N.A.R.M.; Hamada, A.M. Zinc Oxide Nanoparticles Induce Changes in the Antioxidant Systems and Macromolecules in the Solanum Nigrum Callus. Egypt. J. Bot. 2020, 60, 503–517. [Google Scholar] [CrossRef]
- Tedeschini, E.; Proietti, P.; Timorato, V.; D’Amato, R.; Nasini, L.; Dei Buono, D.; Businelli, D.; Frenguelli, G. Selenium as Stressor and Antioxidant Affects Pollen Performance in Olea Europaea. Flora—Morphol. Distrib. Funct. Ecol. Plants 2015, 215, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Bartucca, M.L.; Guiducci, M.; Falcinelli, B.; Del Buono, D.; Benincasa, P. Blue:Red LED Light Proportion Affects Vegetative Parameters, Pigment Content, and Oxidative Status of Einkorn (Triticum Monococcum L. Ssp. Monococcum) Wheatgrass. J. Agric. Food Chem. 2020, 68, 8757–8763. [Google Scholar] [CrossRef]
- Peng, C.; Lin, Z.; Lin, G.; Chen, S. The Anti-Photooxidation of Anthocyanins-Rich Leaves of a Purple Rice Cultivar. Sci. China C Life Sci. 2006, 49, 543–551. [Google Scholar] [CrossRef]
- Tsuda, T.; Shiga, K.; Ohshima, K.; Kawakishi, S.; Osawa, T. Inhibition of Lipid Peroxidation and the Active Oxygen Radical Scavenging Effect of Anthocyanin Pigments Isolated from Phaseolus Vulgaris L. Biochem. Pharmacol. 1996, 52, 1033–1039. [Google Scholar] [CrossRef]
- Gould, K.S.; Kuhn, D.N.; Lee, D.W.; Oberbauer, S.F. Why Leaves Are Sometimes Red. Nature 1995, 378, 241–242. [Google Scholar] [CrossRef]
- Bustos-Torres, K.A.; Vazquez-Rodriguez, S.; la Cruz, A.M.; Sepulveda-Guzman, S.; Benavides, R.; Lopez-Gonzalez, R.; Torres-Martínez, L.M. Influence of the Morphology of ZnO Nanomaterials on Photooxidation of Polypropylene/ZnO Composites. Mater. Sci. Semicond. Process. 2017, 68, 217–225. [Google Scholar] [CrossRef]
- Baskar, V.; Nayeem, S.; Kuppuraj, S.P.; Muthu, T.; Ramalingam, S. Assessment of the Effects of Metal Oxide Nanoparticles on the Growth, Physiology and Metabolic Responses in in Vitro Grown Eggplant (Solanum Melongena). 3 Biotech 2018, 8, 362. [Google Scholar] [CrossRef] [PubMed]
- Raigond, P.; Raigond, B.; Kaundal, B.; Singh, B.; Joshi, A.; Dutt, S. Effect of Zinc Nanoparticles on Antioxidative System of Potato Plants. J. Environ. Biol. 2017, 38, 435–439. [Google Scholar] [CrossRef]
- Dobrikova, A.G.; Apostolova, E.L.; Hanć, A.; Yotsova, E.; Borisova, P.; Sperdouli, I.; Adamakis, I.-D.S.; Moustakas, M. Cadmium Toxicity in Salvia Sclarea L.: An Integrative Response of Element Uptake, Oxidative Stress Markers, Leaf Structure and Photosynthesis. Ecotoxicol. Environ. Saf. 2021, 209, 111851. [Google Scholar] [CrossRef]
- Salih, A.M.; Al-Qurainy, F.; Khan, S.; Tarroum, M.; Nadeem, M.; Shaikhaldein, H.O.; Gaafar, A.-R.Z.; Alfarraj, N.S. Biosynthesis of Zinc Oxide Nanoparticles Using Phoenix Dactylifera and Their Effect on Biomass and Phytochemical Compounds in Juniperus Procera. Sci. Rep. 2021, 11, 19136. [Google Scholar] [CrossRef]
- Zafar, H.; Abbasi, B.H.; Zia, M. Physiological and Antioxidative Response of Brassica Nigra (L.) to ZnO Nanoparticles Grown in Culture Media and Soil. Toxicol. Environ. Chem. 2019, 101, 281–299. [Google Scholar] [CrossRef]
- Tombuloglu, H.; Slimani, Y.; Tombuloglu, G.; Alshammari, T.; Almessiere, M.; Korkmaz, A.D.; Baykal, A.; Samia, A.C.S. Engineered Magnetic Nanoparticles Enhance Chlorophyll Content and Growth of Barley through the Induction of Photosystem Genes. Environ. Sci. Pollut. Res. 2020, 27, 34311–34321. [Google Scholar] [CrossRef]
- Tombuloglu, H.; Albenayyan, N.; Slimani, Y.; Akhtar, S.; Tombuloglu, G.; Almessiere, M.; Baykal, A.; Ercan, I.; Sabit, H.; Manikandan, A. Fate and Impact of Maghemite (γ-Fe2O3) and Magnetite (Fe3O4) Nanoparticles in Barley (Hordeum Vulgare L.). Environ. Sci. Pollut. Res. 2022, 29, 4710–4721. [Google Scholar] [CrossRef]
- Tombuloglu, H.; Slimani, Y.; AlShammari, T.M.; Bargouti, M.; Ozdemir, M.; Tombuloglu, G.; Akhtar, S.; Sabit, H.; Hakeem, K.R.; Almessiere, M.; et al. Uptake, Translocation, and Physiological Effects of Hematite (α-Fe2O3) Nanoparticles in Barley (Hordeum Vulgare L.). Environ. Pollut. 2020, 266, 115391. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Chl a (mg g−1 FW) | Chl b (mg g−1 FW) | Tot Chl (mg g−1 FW) | Soluble Protein (mg g−1 FW) | |
|---|---|---|---|---|
| Control | 2.25 ± 0.22 b | 0.76 ± 0.25 c | 3.01 ± 0.81 c | 9.7 ± 0.4 c |
| 2 mg L−1 | 2.27 ± 0.27 b | 0.91 ± 0.10 bc | 3.18 ± 0.32 bc | 11.4 ± 1.2 b |
| 6 mg L−1 | 2.80 ± 0.19 a | 1.20 ± 0.13 ab | 4.00 ± 0.29 ab | 14.2 ± 0.5 a |
| 18 mg L−1 | 3.13 ± 0.46 a | 1.30 ± 0.21 ab | 4.43 ± 0.70 a | 15.3 ± 2.0 a |
| Carotenoids (mg g−1 FW) | Anthocyanins (mg g−1 FW) | TPC (mg g−1 FW GAE) | DPPH (%) | |
|---|---|---|---|---|
| Control | 3.23 ± 0.59 bc | 0.083 ± 0.010 b | 2.11 ± 0.10 b | 52.7 ± 3.1 b |
| 2 mg L−1 | 2.70 ± 0.73 c | 0.099 ± 0.014 ab | 2.63 ± 0.36 ab | 49.7 ± 3.6 b |
| 6 mg L−1 | 4.24 ± 0.05 ab | 0.098 ± 0.110 ab | 2.75 ± 0.22 a | 60.3 ± 2.0 a |
| 18 mg L−1 | 4.80 ± 0.90 a | 0.110 ± 0.015 a | 2.62 ± 0.17 a | 64.5 ± 2.9 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Regni, L.; Del Buono, D.; Micheli, M.; Facchin, S.L.; Tolisano, C.; Proietti, P. Effects of Biogenic ZnO Nanoparticles on Growth, Physiological, Biochemical Traits and Antioxidants on Olive Tree In Vitro. Horticulturae 2022, 8, 161. https://doi.org/10.3390/horticulturae8020161
Regni L, Del Buono D, Micheli M, Facchin SL, Tolisano C, Proietti P. Effects of Biogenic ZnO Nanoparticles on Growth, Physiological, Biochemical Traits and Antioxidants on Olive Tree In Vitro. Horticulturae. 2022; 8(2):161. https://doi.org/10.3390/horticulturae8020161
Chicago/Turabian StyleRegni, Luca, Daniele Del Buono, Maurizio Micheli, Simona Lucia Facchin, Ciro Tolisano, and Primo Proietti. 2022. "Effects of Biogenic ZnO Nanoparticles on Growth, Physiological, Biochemical Traits and Antioxidants on Olive Tree In Vitro" Horticulturae 8, no. 2: 161. https://doi.org/10.3390/horticulturae8020161
APA StyleRegni, L., Del Buono, D., Micheli, M., Facchin, S. L., Tolisano, C., & Proietti, P. (2022). Effects of Biogenic ZnO Nanoparticles on Growth, Physiological, Biochemical Traits and Antioxidants on Olive Tree In Vitro. Horticulturae, 8(2), 161. https://doi.org/10.3390/horticulturae8020161