
The Effective Role of Nano-Silicon Application in Improving the Productivity and Quality of Grafted Tomato Grown under Salinity Stress


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Grafting of Test Plants
2.3. Greenhouse Experiment
2.3.1. Growth Conditions
2.3.2. Salinity Treatments
2.3.3. Preparation of Nano-Silicon
2.3.4. Plant Growth and Yield
2.3.5. Characteristics of Fruit Quality
2.3.6. Physiological Parameters
2.3.7. Activity of Antioxidant Enzymes, Gibberellic Acid, Abscisic Acid Content, and Leaf Proline
2.3.8. Mineral Composition in Tomato Plants
2.3.9. Statistical Analysis
3. Results
3.1. Plant Growth Parameters
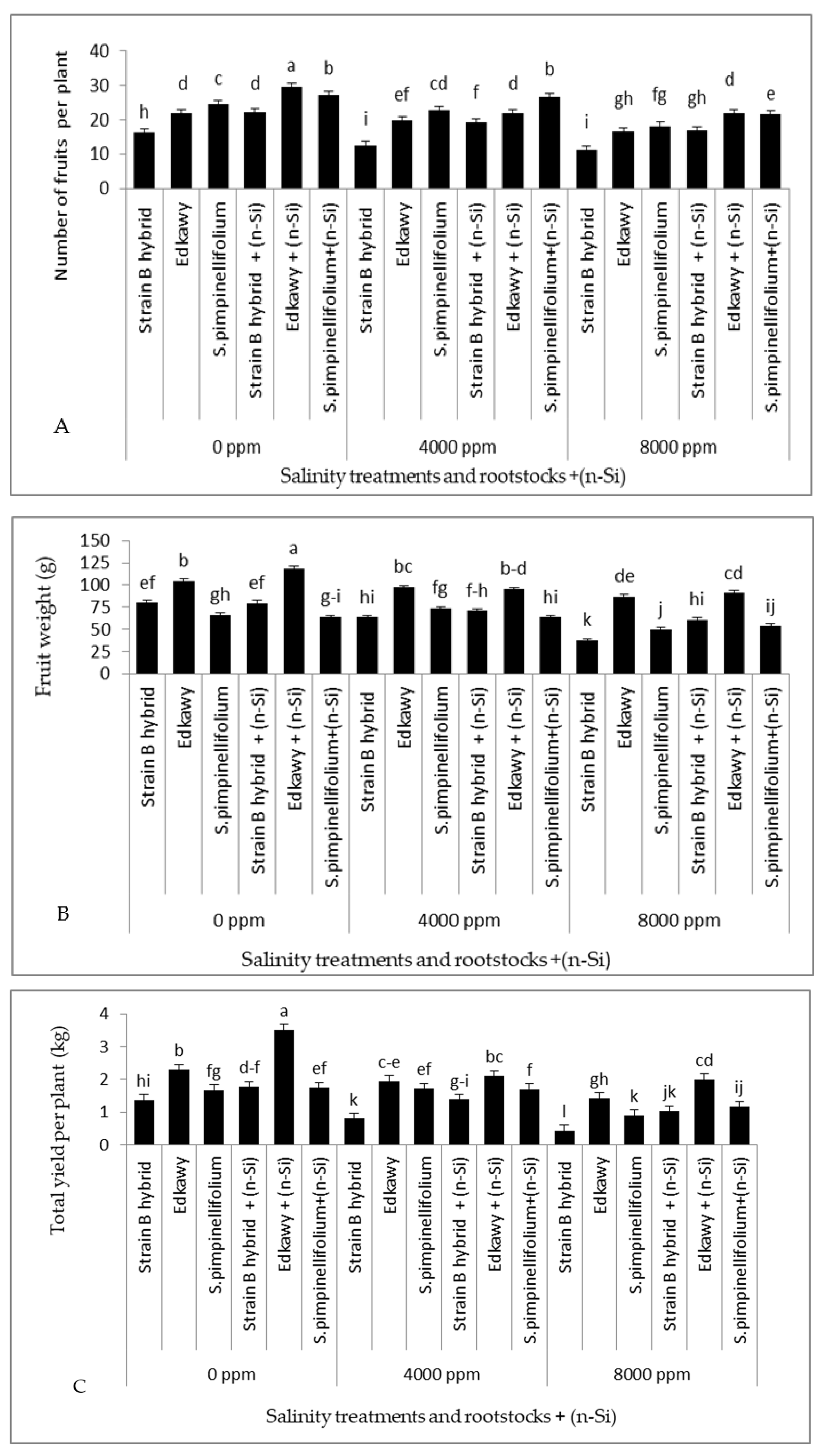
3.2. Tomato Fruit Yield and Its Components
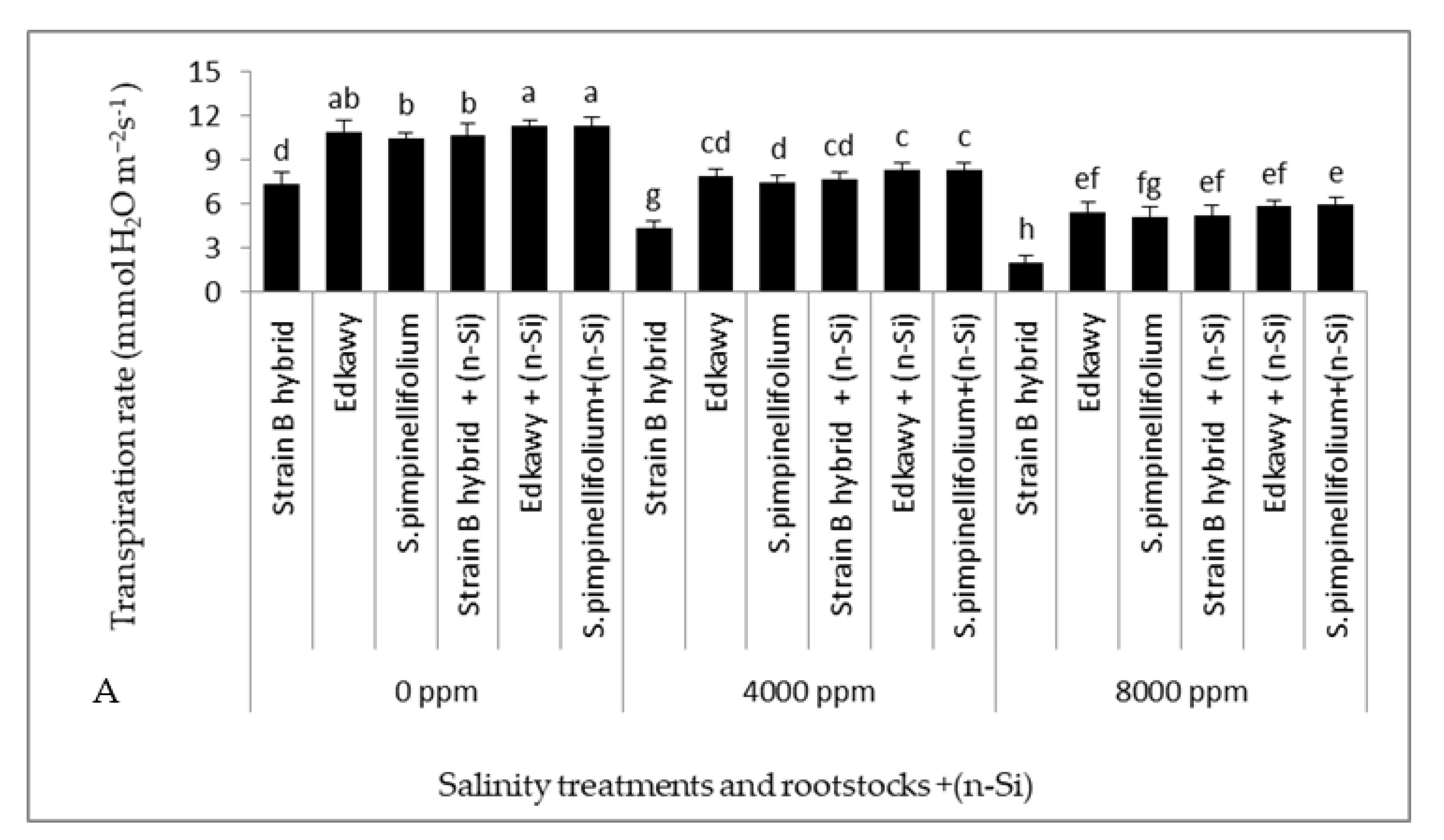
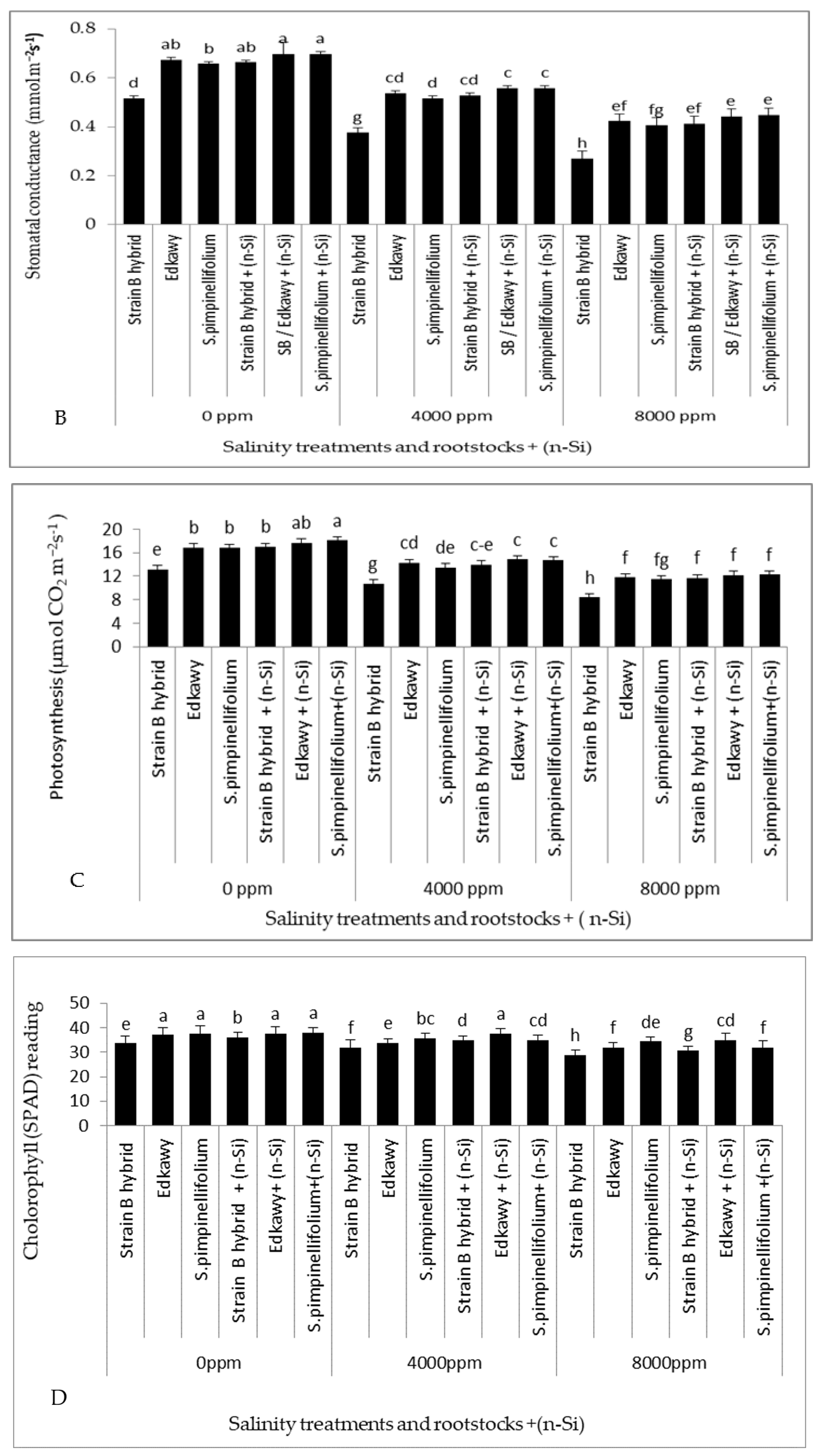
3.3. Physiological Traits of Tomato Plants
3.4. Quality Parameters of Tomato Fruits
3.5. Mineral Content in Tomato Shoots
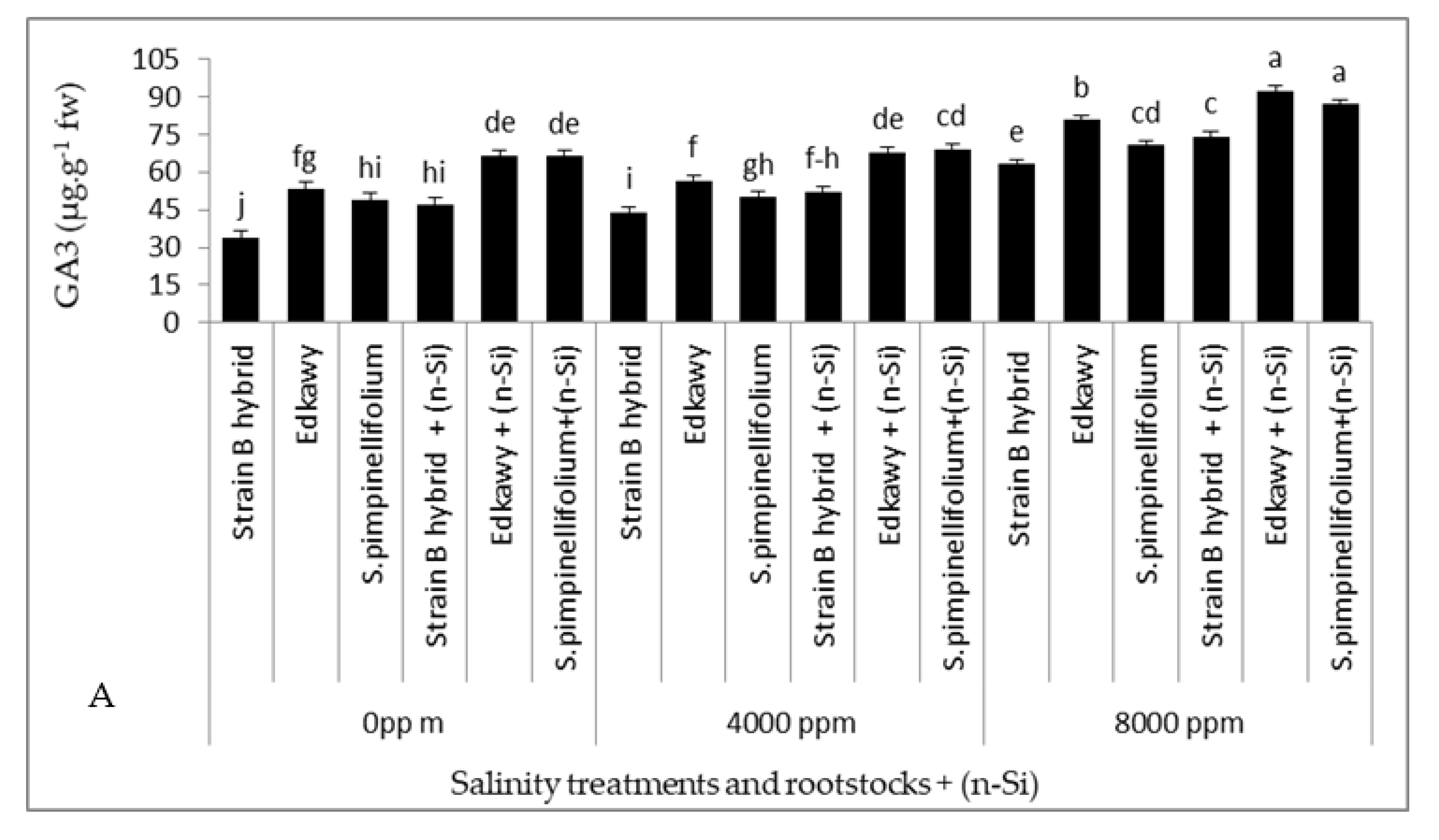
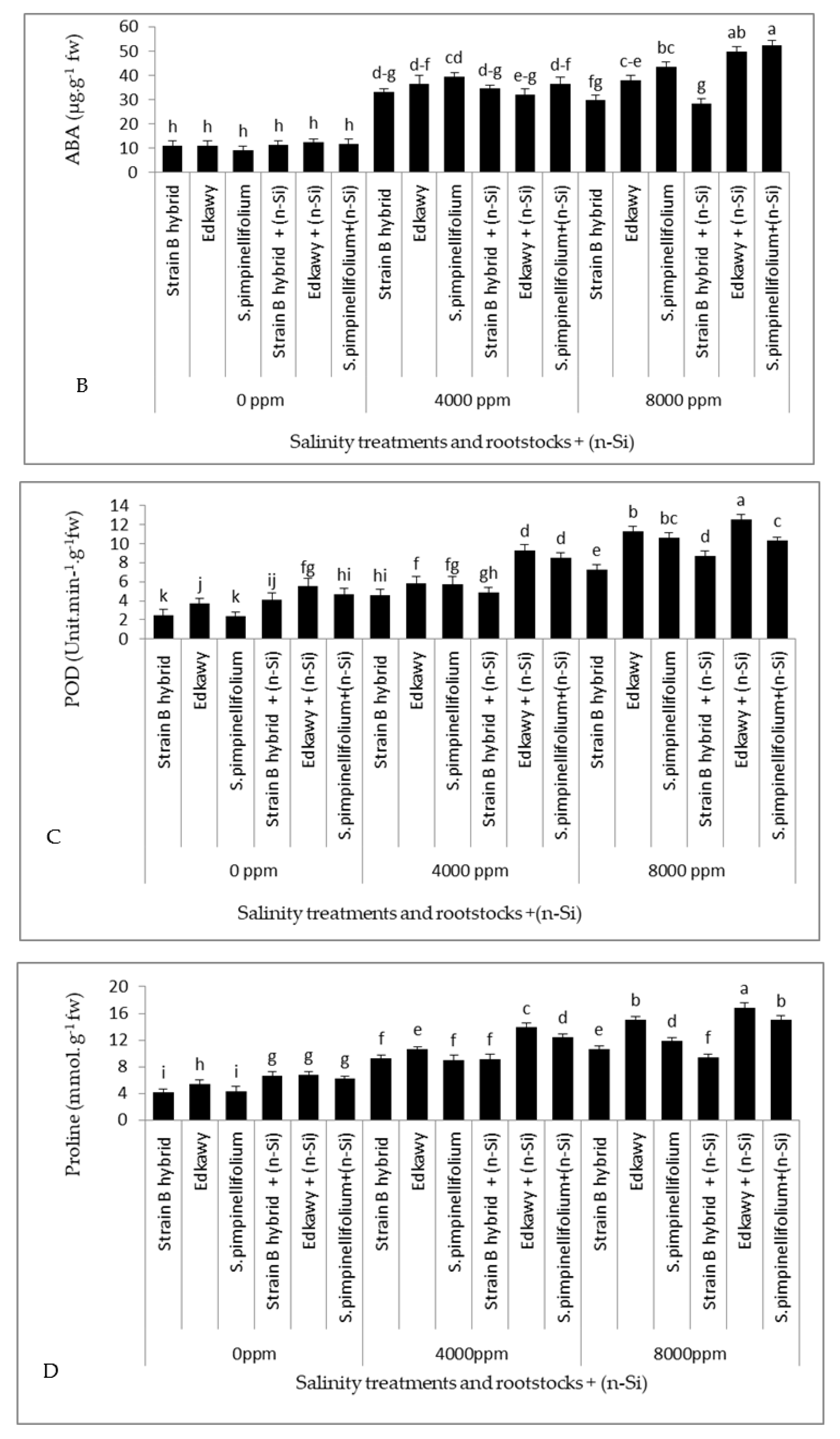
3.6. Plant Hormones, Antioxidant Enzymes, and Proline Content in Tomato Shoots
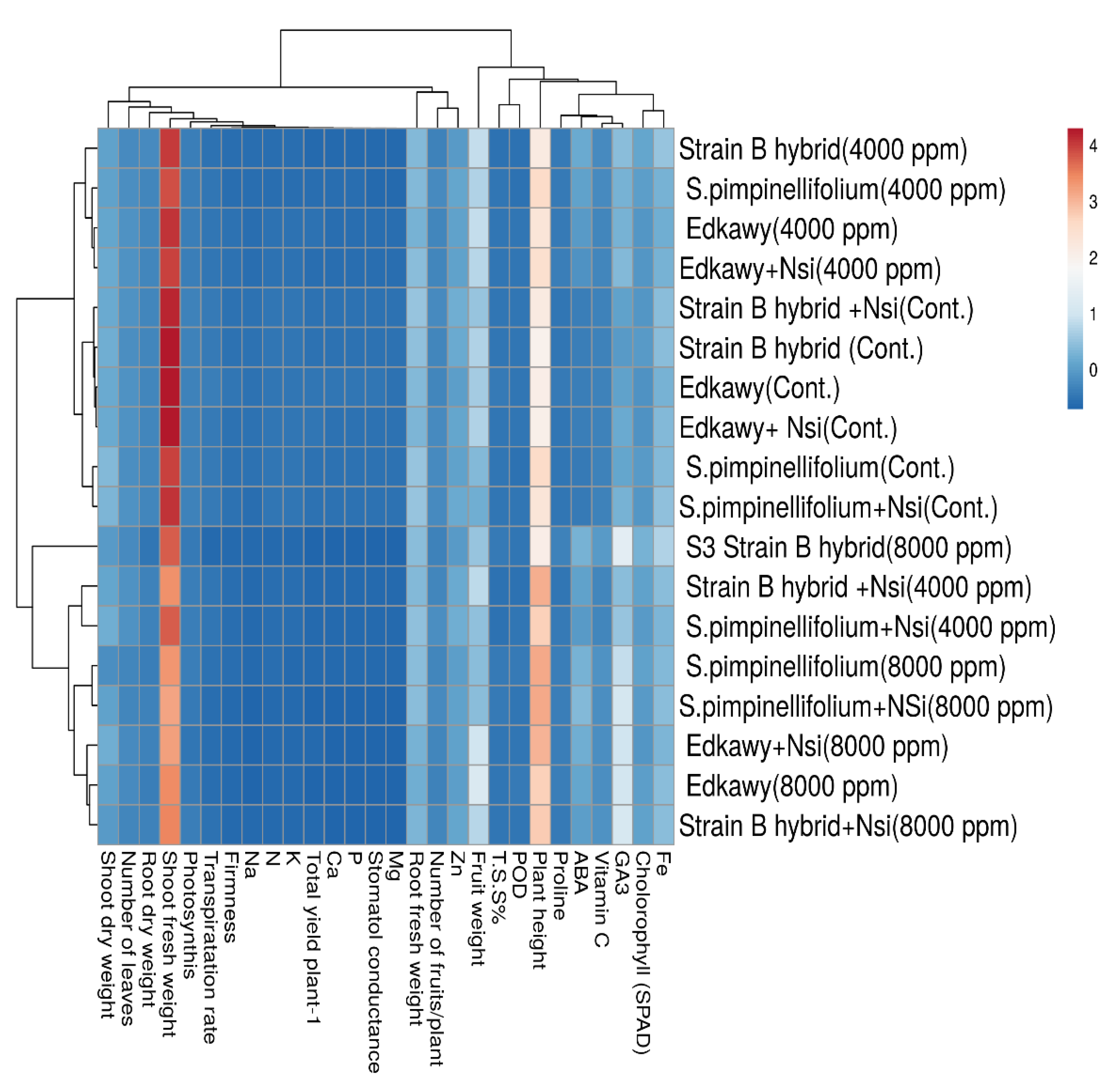
3.7. Clustering Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Cesare, L.F.; Migliori, C.; Ferrari, V.; Parisi, M.; Campanelli, G.; Candido, V.; Perrone, D. Effects of irrigation-fertilization and irrigation-mycorrhization on the alimentary and nutraceutical properties of tomatoes. In Irrigation Systems and Practices in Challenging Environments; TECH Press: Rijeka, Croatia, 2012. [Google Scholar]
- Abdel-Monaim, M.F. Induced Systemic Resistance in Tomato Plants against Fusarium Wilt Disease. Int. Res. J. Microbiol. 2012, 3, 14–23. [Google Scholar]
- Gerster, H. The potential role of lycopene for human health. J. Am. Coll. Nutr. 1997, 16, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Ahanger, M.A.; Alam, P.; Alyemeni, M.N.; Wijaya, L.; Ali, S.; Ashraf, M. Silicon (Si) Supplementation Alleviates NaCl Toxicity in Mung Bean [Vigna radiata (L.) Wilczek] Through the Modifications of Physio-biochemical Attributes and Key Antioxidant Enzymes. J. Plant Growth Regul. 2019, 38, 70–82. [Google Scholar] [CrossRef]
- Shahid, S.A.; Zaman, M.; Heng, L. Soil Salinity: Historical Perspectives and a World Overview of the Problem. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer: Cham, Switzerland, 2018; pp. 43–53. [Google Scholar]
- Singh, K.; Chatrath, R. Application of Physiology in Wheat Breeding; International Maize and Wheat Improvement Center (CIMMYT): Texcoco, Mexico, 2001. [Google Scholar]
- Ashraf, M. Some important physiological selection criteria for salt tolerance in plants. Flora-Morphol. Distrib. Funct. Ecol. Plants 2004, 199, 361–376. [Google Scholar] [CrossRef]
- El-Mogy, M.M.; Atia, M.A.M.; Dhawi, F.; Fouad, A.S.; Bendary, E.S.A.; Khojah, E.; Samra, B.N.; Abdelgawad, K.F.; Ibrahim, M.F.M.; Abdeldaym, E.A. Towards Better Grafting: SCoT and CDDP Analyses for Prediction of the Tomato Rootstocks Performance under Drought Stress. Agronomy 2022, 12, 153. [Google Scholar] [CrossRef]
- Santa-Cruz, A.; Martinez-Rodriguez, M.M.; Perez-Alfocea, F.; Romero-Aranda, R.; Bolarin, M.C. The rootstockeffect on the tomato salinity response depends on the shoot genotype. Plant Sci. 2002, 162, 825–831. [Google Scholar] [CrossRef]
- Razali, R.; Bougouffa, S.; Morton, M.J.L.; Lightfoot, D.J.; Alam, I.; Essack, M.; Arold, S.T.; Kamau, A.A.; Schmöckel, S.M.; Pailles, Y.; et al. The Genome Sequence of the Wild Tomato S. pimpinellifolium Provides Insights Into SalinityTolerance. Front. Plant Sci. 2018, 9, 1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouphael, Y.; Petropoulos, S.A.; Cardarelli, M.; Colla, G. Salinity as eustress or for enhancing the quality of vegetables. Sci. Hortic. 2018, 234, 361–369. [Google Scholar] [CrossRef]
- Hoffmann, J.; Berni, R.; Hausman, J.F.; Guerriero, G. A Review on the Beneficial Role of Silicon against Salinity in Non-Accumulator Crops: Tomato as a Model. Biomolecules 2020, 10, 1284. [Google Scholar] [CrossRef]
- Almeida, D.M.; Margarida Oliveira, M.; Saibo, N.J.M. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40, 326–345. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Yin, Q.; Yan, Z.; Wang, Y.; Niu, J.; Zhang, J.; Fan, K. Exogenous Silicon Enhanced Salt Resistance by Maintaining K+/Na+ Homeostasis and Antioxidant Performance in Alfalfa Leaves. Front. Plant Sci. 2020, 11, 1183. [Google Scholar] [CrossRef] [PubMed]
- Tuna, A.L.; Kaya, C.; Higgs, D.; Murillo-Amador, B.; Aydemir, S.; Girgin, A.R. Silicon improves salinity tolerance in wheat plants. Environ. Exp. Bot. 2008, 62, 10–16. [Google Scholar] [CrossRef]
- Sommer, M.; Kaczorek, D.; Kuzyakov, Y.; Breuer, J. Silicon pools and fluxes in soils and landscapes—A review. J. Plant Nutr. Soil Sci. 2006, 169, 310–329. [Google Scholar] [CrossRef]
- Frew, A.; Weston, L.A.; Reynolds, O.L.; Gurr, G.M. The role of silicon in plant biology: A paradigm shift in research approach. Ann. Bot. 2018, 121, 1265–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barcelo, J.; Guevara, P.; Poschenrieder, C. Silicon amelioration of aluminium toxicity in teosinte (Zea mays L. ssp. mexicana). Plant Soil 2004, 154, 249–255. [Google Scholar] [CrossRef]
- Cocker, K.M.; Evans, D.E.; Hodson, M.J. The amelioration of aluminium toxicity by silicon in wheat (Triticumaestivum L.): Malate exudation as evidence for an in planta mechanism. Planta 1998, 204, 318–323. [Google Scholar] [CrossRef]
- Shen, X.; Xiao, X.; Dong, Z.; Chen, Y. Silicon effects on antioxidative enzymes and lipid peroxidation in leaves and roots of peanut under aluminum stress. Acta Physiol. Plant. 2014, 36, 3063–3069. [Google Scholar] [CrossRef]
- Sivanesan, I.; Jeong, B.R. Silicon promotes adventitious shoot regeneration and enhances salinity tolerance of Ajuga multiflora bunge by altering activity of antioxidant enzyme. Sci. World J. 2014, 2014, 521703. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Lee, I.J. Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress: A review. Front. Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.; Bashri, G.; Shweta Singh, S.; Ahmad, P.; Singh, V.; Prasad, S.; Dubey, N.; Chauhan, D. Chapter 19 Efficacy of Silicon against Aluminum Toxicity in Plants: An Overview. In Silicon in Plants: Advances and Future Prospects; CRC Press: Boca Raton, FL, USA, 2016; pp. 241–258. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alamri, S.A.; Alam, P.; Ashraf, M.; Ahmad, P. Potential of exogenously sourced kinetin in protecting Solanumlycopersicum from NaCl-induced oxidative stress through up-regulation of the antioxidant system, ascorbate-glutathione cycle and glyoxalase system. PLoS ONE 2018, 13, e0202175. [Google Scholar] [CrossRef]
- Ahmad, R.; Hussain, S.; Anjum, M.A.; Khalid, M.F.; Saqib, M.; Zakir, I.; Hassan, A.; Fahad, S.; Ahmad, S. Oxidative Stress and Antioxidant Defense Mechanisms in Plants Under Salt Stress. In Plant Abiotic Stress Tolerance; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Stamatakis, A.; Papadantonakis, N.; Lydakis-Simantiris, N.; Kefalas, P.; Savvas, D. Effects of silicon and salinity on fruit yield and quality of tomato grown hydroponically. Acta Hortic. 2003, 609, 141–147. [Google Scholar] [CrossRef]
- Al-Aghabary, K.; Zhu, Z.; Shi, Q. Influence of Silicon Supply on Chlorophyll Content, Chlorophyll Fluorescence, and Antioxidative Enzyme Activities in Tomato Plants Under Salt Stress. J. Plant Nutr. 2005, 27, 2101–2115. [Google Scholar] [CrossRef]
- Romero-Aranda, M.R.; Jurado, O.; Cuartero, J. Silicon alleviates the deleterious salt effect on tomato plant growth by improving plant water status. J. Plant Physiol. 2006, 163, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Seal, P.; Das, P.; Biswas, A.K.; Seal, P.; Das, P.; Biswas, A.K. Versatile Potentiality of Silicon in Mitigation of Biotic and Abiotic Stresses in Plants: A Review. Am. J. Plant Sci. 2018, 9, 1433–1454. [Google Scholar] [CrossRef] [Green Version]
- Haghighi, M.; Pessarakli, M. Influence of silicon and nano--silicon on salinity tolerance of cherry tomatoes (Solanum lycopersicum L.) at early growth stage. Sci. Hortic. 2013, 161, 111–117. [Google Scholar] [CrossRef]
- Tondey, M.; Kalia, A.; Singh, A.; Dheri, G.S.; Taggar, M.S.; Nepovimova, E.; Krejcar, O.; Kuca, K. Seed Priming and Coating by Nano-Scale Zinc Oxide Particles Improved Vegetative Growth, Yield and Quality of Fodder Maize (Zea mays). Agronomy 2021, 11, 729. [Google Scholar] [CrossRef]
- Mahmoud, A.W.M.; Samy, M.M.; Sany, H.; Eid, R.R.; Rashad, H.M.; Abdeldaym, E.A. Nano-potassium, Nano-silicon, and Biochar Applications Improve Potato Salt Tolerance by Modulating Photosynthesis, Water Status, and Biochemical Constituents. Sustainability 2022, 14, 723. [Google Scholar] [CrossRef]
- Nasrallah, A.K.; Kheder, A.A.; Kord, M.A.; Fouad, A.S.; El-Mogy, M.M.; Atia, M.A.M. Mitigation of Salinity Stress Effects on Broad Bean Productivity Using Calcium Phosphate Nanoparticles Application. Horticulturae 2022, 8, 75. [Google Scholar] [CrossRef]
- Yuvakkumar, R.; Elango, V.; Rajendran, V.; Kannan, N.S.; Prabu, P. Influence of nanosilica powder on the growth of maize crop. Int. J. Green Nanotechnol. 2011, 3, 180–190. [Google Scholar] [CrossRef]
- Suriyaprabha, R.; Karunakaran, G.; Yuvakkumar, R.; Rajendran, V.; Kannan, N. Silica nanoparticles for increased silica availability in maize (Zea mays L.) seeds under hydroponic conditions. Curr. Nanosci. 2012, 8, 902–908. [Google Scholar] [CrossRef]
- Haghighi, M.; Afifipour, Z.; Mozafarian, M. The effect of N-Si on tomato seed germination under salinity levels. J. Biol. Environ. Sci. 2012, 6, 87–90. [Google Scholar]
- Sabaghnia, N.; Janmohammadi, M. Graphic analysis of nano-silicon by salinity stress interaction on germination properties of lentil using the biplot method. Agric. For. 2014, 60, 29–40. [Google Scholar]
- Xie, Y.; Li, B.; Tao, G.; Zhang, Q.; Zhang, C. Effects of nano--silicon dioxide on photosynthetic fluorescence characteristics of Indocalamus barbatus McClure. J. Nanjing For. Univ. 2012, 36, 59–63. [Google Scholar]
- Mahmoud, A.W.M.; Sweaty, H.M. Comparison between commercial and nano-nPk in presence of nano- ze-polite on sage plant yield and its components under water stress. Agriculture 2020, 66, 24–39. [Google Scholar] [CrossRef]
- Janmohammadi, M.; Amanzadeh, T.; Sabaghnia, N.; Ion, V. Effect of nano-silicon foliar application on safflower growth under organic and inorganic fertilizer regimes. Bot. Lith. 2016, 22, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Gong, H. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2014, 34, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.W.M.; Abdeldaym, E.A.; Abdelaziz, S.M.; El-Sawy, M.B.I.; Mottaleb, S.A. Synergetic Effects of Zinc, Boron, Silicon, and Zeolite Nano-particles on Confer Tolerance in Potato Plants Subjected to Salinity. Agronomy 2019, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Polle, A.; Otter, T.; Mehne-Jakobs, B. Effect of magnesium deficiency on antioxidative systems in needles of Norway spruce [Picea abies (L) Karst.] grown with different ratios of nitrate and ammonium as nitrogen souices. New Phytol. 1994, 128, 621–628. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Fales, T.M.; Jaouni, J.F.; Babashak, I. Simple device for preparing ethereal diazomethane without resorting to codistillation. Ann. Chem. 1973, 45, 2302–2303. [Google Scholar] [CrossRef]
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis; Association of Official Analytical Chemists: Washington, DC, USA, 1990. [Google Scholar]
- Singh, A.K.; Dubey, R.S. Changes in chlorophyll a and b contents and activities of photosystems 1 and 2 in rice seedlings inducedby NaCl. Photosynthetica 1995, 31, 489–499. [Google Scholar]
- Gomez, K.N.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley and Sons: New York, NY, USA, 1984. [Google Scholar]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; Iowa State University Press: Ames, IA, USA, 1980; p. 507. [Google Scholar]
- Abdeldym, E.A.; El-Mogy, M.M.; Abdellateaf, H.R.L.; Atia, M.A.M. Genetic Characterization, Agro-Morphological and Physiological Evaluation of Grafted Tomato under Salinity Stress Conditions. Agronomy 2020, 10, 1948. [Google Scholar] [CrossRef]
- Singh, H.; Kumar, P.; Kumar, A.; Kyriacou, M.C.; Colla, G.; Rouphael, Y. Grafting Tomato as a Tool to Improve Salt Tolerance. Agronomy 2020, 10, 263. [Google Scholar] [CrossRef] [Green Version]
- Cuartero, J.; Fernández-Muñoz, R. Tomato and salinity. Sci. Hortic. 1998, 78, 83–125. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, K.A.A.; Mazrou, Y.S.; Hafez, Y.M. Silicon Foliar Application Mitigates Salt Stress in Sweet Pepper Plants by Enhancing Water Status, Photosynthesis, Antioxidant Enzyme Activity and Fruit Yield. Plants 2020, 9, 733. [Google Scholar] [CrossRef]
- Chaichi, M.R.; Keshavarz-Afshar, R.; Lu, B.; Rostamza, M. Growth and nutrient uptake of tomato in response to application of saline water, biological fertilizer, and surfactant. J. Plant Nutr. 2017, 40, 457–466. [Google Scholar] [CrossRef]
- Ali, E.G.; Mohamed, M.F.; Farghali, M.A.; Abdel-Rahman, M.S. Productivity of Grafted Tomato Grown in the Summer Season Under The New Valley Environmental Conditions. Assiut J. Agric. Sci. 2014, 45, 2. [Google Scholar]
- Mohamed, F.M.; Abd ElKader, M.F.; Shalaby, G.I. New Potential Hybrid ‘Assiut-15’ for Production of Tomato under Adverse High Temperature Conditions. Proc. Third Sci. Conf. Agric. Sci. 2002, 385–392. [Google Scholar]
- Aloni, B.; Cohen, R.; Karni, L.; Aktas, H.; Edelstein, M. Hormonal signaling in rootstock–scion interactions. Sci. Hortic. 2010, 127, 119–126. [Google Scholar] [CrossRef]
- Harada, T. Grafting and RNAtransport via phloem tissue in horticultural plants. Sci. Hortic. 2010, 125, 545–550. [Google Scholar] [CrossRef]
- Effat, A.; Sanaa, A.; Horeya, M.; Ramzy, M.; Yehia, A.; Manal, A. Comparative Performance of Five Genotypes of Tomato to Salt Stress. Alex. Sci. Exch. J. 2018, 39, 3. [Google Scholar]
- Zhang, Y.; Shi, Y.; Gong, H.-J.; Zhao, H.-L.; Li, H.-L.; Hu, Y.-H.; Wang, Y.-C. Beneficial effects of silicon on photosynthesis of tomato seedlings under water stress. J. Integr. Agric. 2018, 17, 2151–2159. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.H.; Houborg, R.; McCabe, M.F. Response of Chlorophyll, Carotenoid and SPAD-502 Measurement to Salinity and Nutrient Stress in Wheat (Triticumaestivum L.). Agronomy 2017, 7, 61. [Google Scholar] [CrossRef] [Green Version]
- Kusumi, K.; Hirotsuka, S.; Kumamaru, T.; Iba, K. Increased leaf photosynthesis caused by elevated stomatal conductance in a rice mutant deficient in SLAC1, a guard cell anion channel protein. J. Exp. Bot. 2012, 63, 5635–5644. [Google Scholar] [CrossRef] [Green Version]
- Saglam, A.; Terzi, R.; Demiralay, M. Effect of polyethylene glycol induced drought stress on photosynthesis in two chickpea genotypes with different drought tolerance. Acta Biol. Hung. 2014, 65, 178–188. [Google Scholar] [CrossRef]
- Gong, H.; Chen, K. The regulatory role of silicon on water relations, photosynthetic gas exchange, and carboxylation activities of wheat leaves in field drought conditions. Acta Physiol. Plant. 2012, 34, 1589–1594. [Google Scholar] [CrossRef]
- Al-Ismaily, S.S.; Al-Yahyai, R.A.; Al-Rawahy, S.A. Mixed Fertilizer can Improve Fruit Yield and Quality of Field-Grown Tomatoes Irrigated with Saline Water. J. Plant Nutr. 2014, 37, 1981–1996. [Google Scholar] [CrossRef]
- Abdelgawad, K.; El-Mogy, M.M.; Mohamed, M.I.A.; Garchery, C.; Stevens, R.G. Increasing Ascorbic Acid Content and Salinity Tolerance of Cherry Tomato Plants by Suppressed Expression of the Ascorbate Oxidase Gene. Agronomy 2019, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- El-Mogy, M.M.; Garchery, C.; Stevens, R. Irrigation with salt water affects growth, yield, fruit quality, storability and marker-gene expression in cherry tomato. Acta Agric. Scand. Sect. B Soil Plant Sci. 2018, 68, 727–737. [Google Scholar] [CrossRef]
- Dehghanipoodeh, S.; Ghobadi, C.; Baninasab, B.; Gheysari, M.; Bidabadi, S.S. Effects of potassium silicate and nano-silica on quantitative and qualitative characteristics of a commercial strawberry (Fragaria × ananassa cv. ‘camarosa’). J. Plant Nutr. 2016, 39, 502–507. [Google Scholar] [CrossRef]
- Khan, A.; Khan, A.L.; Muneer, S.; Kim, Y.H.; Al-Rawahi, A.; Al-Harrasi, A. Silicon and Salinity: Crosstalk in Crop-Mediated Stress Tolerance Mechanisms. Front. Plant Sci. 2019, 10, 1429. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.K.; Sohn, E.Y.; Hamayun, M.; Yoon, J.Y.; Lee, I.J. Effect of silicon on growth and salinity stress of soybean plant grown under the hydroponic system. Agrofor. Syst. 2010, 80, 333–340. [Google Scholar] [CrossRef]
- Pena-Cortes, H.; Sanchez-Serrano, J.J.; Mertens, R.; Willmitzer, L.; Prat, S. Abscisic acid is involved in the wound-induced expression of the proteinase inhibitor II gene in potato and tomato. Proc. Natl. Acad. Sci. USA 1989, 86, 9851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.; Kamran, M.; Imran, M.; Al-Harrasi, A.; Al-Rawahi, A.; Al-Amri, I.; Lee, I.J.; Khan, A.L. Silicon and salicylic acid confer high-pH stress tolerance in tomato seedlings. Sci. Rep. 2019, 9, 19788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahlaoui, B.; Hachicha, M.; Misle, E.; Fidalgo, F.; Teixeira, J. Physiological and biochemical responses to the exogenous application of proline of tomato plants irrigated with saline water. J. Saudi Soc. Agric. Sci. 2018, 17, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Kafi, M.; Nabati, J.; Ahmadi-Lahijani, M.J.; Oskoueian, A. Silicon compounds and potassium sulfate improve salinity tolerance of potato plants through instigating the defense mechanisms, cell membrane stability, and accumulation of osmolytes. Commun. Soil Sci. Plant Anal. 2021, 52, 843–858. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salinity Levels | Rootstocks + (n-Si) | Plant Height (cm) | Number of Leaves/Plant | Shoot Fresh Weight (g) | Shoot Dry Weight (g) | Root Fresh Weight (g) | Root Dry Weight (g) |
|---|---|---|---|---|---|---|---|
| 0 ppm | Strain B hybrid | 161.7 ij | 24.7 e | 306.8 d | 49.37 e | 63.2 e | 17.6 ef |
| Edkawy | 239.0 ab | 35.3 a | 440.7 a | 62.50 c | 81 ab | 21.7 b | |
| S. pimpinellifolium | 221.0 b–d | 25.7 de | 320.7 cd | 65.37 b | 75.3 c | 18.8 de | |
| Strain B hybrid + (n-Si) | 199.3 e–g | 30.0 c | 343.7 c | 53.87 d | 78.0 bc | 20.3 b–d | |
| Edkawy + (n-Si) | 248.0 a | 37.0 a | 463.3 a | 66.43 b | 84.0 a | 25.7 a | |
| S. pimpinellifolium + (n-Si) | 228.3 bc | 34.0 ab | 368.3 b | 69.77 a | 84.0 a | 21.2 bc | |
| 4000 ppm | Strain B hybrid | 117.7 l | 18 gh | 195.8 f–h | 29.33 i | 43.2 h | 16.8 f |
| Edkawy | 194.7 f–h | 29 cd | 314.3 d | 46.70 f | 56.6 f | 19.3 c–e | |
| S. pimpinellifolium | 178.7 hi | 25.3 e | 252.7 e | 39.37 g | 56.0 f | 18.2 ef | |
| Strain B hybrid + (n-Si) | 182.0 gh | 24. 7 e | 198.3 fg | 36.87 h | 50.67 g | 14.8 gh | |
| Edkawy + (n-Si) | 213.7 c–e | 31.3 bc | 320.0 cd | 55.27 d | 70.60 d | 21.3 b | |
| S. pimpinellifolium + (n-Si) | 196.0 e–h | 31.0 bc | 256.3 e | 49.43 e | 63.67 e | 17.6 ef | |
| 8000 ppm | Strain B hybrid | 82.67 m | 14.3 i | 133.7 i | 20.17l g | 33.90 i | 7.2 i |
| Edkawy | 158.0 jk | 22.3 ef | 191.7 gh | 35.53 h | 43.27 h | 14.7 h | |
| S. pimpinellifolium | 178.7 hi | 17.7 g–i | 188.3 gh | 22.87 k | 49.33 g | 13.2 h | |
| Strain B hybrid + (n-Si) | 142.0 k | 15.3 hi | 171.7 h | 26.63 j | 40.67 h | 13.5 h | |
| Edkawy + (n-Si) | 207.0 d–f | 24.7 e | 216.3 f | 49.53 e | 53.93 fg | 18.5 d–f | |
| S. pimpinellifolium + (n-Si) | 189.3 f–h | 20.0 fg | 191.7 gh | 36.60 h | 53.33 fg | 16.7 fg | |
| LSD value at 0.05: | 18.5 | 3.7 | 24.24 | 2 | 4.6 | 1.97 | |
| Salinity Levels | Rootstocks + (n-Si) | TSS% (Brix°) | Firmness (n) | Vitamin C (mg/100 g FW) |
|---|---|---|---|---|
| 0 ppm | Strain B hybrid | 4.43 h | 1.467 j | 14.67 j |
| Edkawy | 5.60 fg | 2.150 hi | 18.27 hi | |
| S. pimpinellifolium | 6.20 ef | 2.08 i | 16.33 ij | |
| Strain B hybrid + (n-Si) | 4.33 h | 1.67 j | 16.33 ij | |
| Edkawy + (n-Si) | 5.67 f | 2.43 e–h | 19.27 f-h | |
| S. pimpinellifolium + (n-Si) | 6.23 ef | 2.37 f–i | 18.93 gh | |
| 4000 ppm | Strain Bhybrid | 5.10 gh | 2.22 g–i | 17.67 hi |
| Edkawy | 6.43 ef | 3.017 ab | 20.60 e–g | |
| S. pimpinellifolium | 6.37 ef | 2.62 d-f | 20.33 e–g | |
| Strain B hybrid + (n-Si) | 6.07 ef | 2.48 e–g | 18.67 gh | |
| Edkawy + (n-Si) | 6.53 e | 3.017 ab | 23.33 b-d | |
| S.pimpinellifolium + (n-Si) | 6.43 ef | 3.02 ab | 21.33 de | |
| 8000 ppm | Strain B hybrid | 7.87 d | 2.78 b–d | 21.07 ef |
| Edkawy | 9.53 c | 3.18 a | 23.67 bc | |
| S. pimpinellifolium | 10.43 b | 2.98 a–c | 23.67 bc | |
| Strain B hybrid + (n-Si) | 8.23 d | 2.72 c–e | 22.0 de | |
| Edkawy + (n-Si) | 11.30 a | 3.12 a | 26.67 a | |
| S. pimpinellifolium + (n-Si) | 11.47 a | 3.08 a | 25.33 ab | |
| LSD value at 0.05: | 0.86 | 0.28 | 2.03 | |
| Salinity Levels | Rootstocks + (n-Si) | N (%) | P (%) | K (%) | Ca (%) | Mg (%) |
|---|---|---|---|---|---|---|
| 0 ppm | Strain B hybrid | 3.70 de | 0.51 f | 4.23 b | 0.38 h | 0.49 d |
| Edkawy | 4.29 c | 0.64 c | 4.72 a | 1.11 c | 0.58 c | |
| S. pimpinellifolium | 3.83 d | 0.57 d | 3.93 c | 0.99 de | 0.48 d | |
| Strain B hybrid + (n-Si) | 4.53 bc | 0.66 c | 4.49 ab | 1.13 c | 0.64 b | |
| Edkawy + (n-Si) | 4.87 a | 0.89 a | 4.71 a | 1.45 a | 0.67 a | |
| S. pimpinellifolium + (n-Si) | 4.617 ab | 0.76 b | 4.70 a | 1.1 c | 0.59 c | |
| 4000 ppm | Strain B hybrid | 2.58 jk | 0.45 g | 2.42 g | 0.34 h | 0.33 h |
| Edkawy | 2.96 g–i | 0.54 ef | 3.21 d | 1.08 cd | 0.37 fg | |
| S. pimpinellifolium | 2.860 h–j | 0.55 de | 2.85 ef | 0.93 e | 0.36 f-h | |
| Strain B hybrid + (n-Si) | 3.260 fg | 0.56 de | 3.11 de | 1.06 cd | 0.41 e | |
| Edkawy + (n-Si) | 3.52 d–f | 0.59 d | 3.31 d | 1.31 b | 0.42 e | |
| S. pimpinellifolium + (n-Si) | 3.45 ef | 0.55 de | 3.27 d | 1.03 cd | 0.40 e | |
| 8000 ppm | Strain B hybrid | 1.987 l | 0.41 h | 1.90 h | 0.6 g | 0.19 j |
| Edkawy | 2.66 i–k | 0.51 f | 2.40 g | 0.69 fg | 0.28 i | |
| S. pimpinellifolium | 2.170 l | 0.46 g | 1.90 h | 0.63 g | 0.18 j | |
| Strain B hybrid + (n-Si) | 2.52 k | 0.44 gh | 2.82 f | 0.64 g | 0.33 gh | |
| Edkawy + (n-Si) | 3.44 ef | 0.56 de | 3.11 de | 0.74 f | 0.37 f | |
| S. pimpinellifolium+ (n-Si) | 2.97 gh | 0.50 f | 2.31 g | 0.73 f | 0.29 i | |
| LSD value at 0.05: | 0.3 | 0.04 | 0.3 | 0.1 | 0.1 | |
| Salinity Levels | Rootstocks + (n-Si) | Na (%) | Fe (ppm) | Zn (ppm) |
|---|---|---|---|---|
| 0 ppm | Strain B hybrid | 0.15 f | 62.28 cd | 44.55 d–g |
| Edkawy | 0.11 fg | 72.01 b | 47.77 de | |
| S. pimpinellifolium | 0.13 fg | 65.18 c | 45.5 d–f | |
| Strain B hybrid + (n-Si) | 0.10 fg | 71.28 b | 54.62 bc | |
| Edkawy + (n-Si) | 0.09 g | 85.30 a | 64.15 a | |
| S. pimpinellifolium + (n-Si) | 0.1 fg | 82.33 a | 58.61 ab | |
| 4000 ppm | Strain B hybrid | 1.670 b | 49.66 jk | 26.43 kl |
| Edkawy | 1.102 de | 52.34 g–j | 42.31 e–h | |
| S. pimpinellifolium | 1.108 de | 50.28 i–k | 41.53 f–h | |
| Strain B hybrid + (n-Si) | 1.070 e | 53.47 f-i | 39.71 g–i | |
| Edkawy + (n-Si) | 1.085 e | 59.4 de | 48.60 d | |
| S. pimpinellifolium + (n-Si) | 1.140 cd | 55.87 fg | 49.54 cd | |
| 8000 ppm | Strain B hybrid | 1.740 a | 42.19 m | 20.91 i |
| Edkawy | 1.187 c | 51.44 h–k | 34.09 ij | |
| S. pimpinellifolium | 1.188 c | 48.50 kl | 32.18 jk | |
| Strain B hybrid + (n-Si) | 1.17 c | 44.94 lm | 33.69 j | |
| Edkawy + (n-Si) | 1.11 de | 57.06 ef | 41.08 f–h | |
| S. pimpinellifolium + (n-Si) | 1.142 cd | 54.20 fh | 36.65 h–j | |
| LSD value at 0.05: | 0.05 | 3.58 | 5.7 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayed, E.G.; Mahmoud, A.W.M.; El-Mogy, M.M.; Ali, M.A.A.; Fahmy, M.A.M.; Tawfic, G.A. The Effective Role of Nano-Silicon Application in Improving the Productivity and Quality of Grafted Tomato Grown under Salinity Stress. Horticulturae 2022, 8, 293. https://doi.org/10.3390/horticulturae8040293
Sayed EG, Mahmoud AWM, El-Mogy MM, Ali MAA, Fahmy MAM, Tawfic GA. The Effective Role of Nano-Silicon Application in Improving the Productivity and Quality of Grafted Tomato Grown under Salinity Stress. Horticulturae. 2022; 8(4):293. https://doi.org/10.3390/horticulturae8040293
Chicago/Turabian StyleSayed, Eman G., Abdel Wahab M. Mahmoud, Mohamed M. El-Mogy, Mahmoud A. A. Ali, Mahmoud A. M. Fahmy, and Ghada A. Tawfic. 2022. "The Effective Role of Nano-Silicon Application in Improving the Productivity and Quality of Grafted Tomato Grown under Salinity Stress" Horticulturae 8, no. 4: 293. https://doi.org/10.3390/horticulturae8040293
APA StyleSayed, E. G., Mahmoud, A. W. M., El-Mogy, M. M., Ali, M. A. A., Fahmy, M. A. M., & Tawfic, G. A. (2022). The Effective Role of Nano-Silicon Application in Improving the Productivity and Quality of Grafted Tomato Grown under Salinity Stress. Horticulturae, 8(4), 293. https://doi.org/10.3390/horticulturae8040293