
Effects of Vermicompost Substrates and Coconut Fibers Used against the Background of Various Biofertilizers on the Yields of Cucumis melo L. and Solanum lycopersicum L.

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Location, and Experimental Design
2.2. Description of Treatments: Substrates Used and PGPMs Applied
2.2.1. Substrates
2.2.2. Biofertilizers: Plant-Growth-Promoting Microorganisms
- Arbuscular mycorrhizal fungi (AMF) consortium: This treatment contained a mix of five different strains of mycorrhizal fungi at 5% (w/w) (equivalent to 150 spores g−1) belonging to five species of the Glomeraceae family—Glomus intraradices (Rhizophagus intraradices, ID 4876), Glomus deserticola (Septoglomus desertícola, ID 1838035), Glomus clarum (Rhizophagus clarus, ID 94130), Glomus mosseae (Funneliformis mosseae, ID 27381) and Glomus aggregatum (ID 241619) [28]. These arbuscular mycorrhizal fungi (AMF) were isolated and extracted from the commercial product Bioradis Tablet (Bioera SL-Constantí, Tarragona, Spain). They were applied during three growth stages (transplant, flowering, and pre-harvest) in each crop using 1 g of AMF (containing 150 spores). This was introduced into the substrate (container) at a 10 cm depth in each plant [28] at each application. In the study, microbial activity was indirectly assessed through enzymatic activity and sap levels, but it was not assessed whether they achieved symbiosis with the plants in the experiment.
- Plant-growth-promoting rhizobacteria (PGPR) consortium: This treatment contained a mix of rhizobacteria, with 5 × 109 CFU (colony forming units) g−1 of isolates of the Paenibacillaceae family—Paenibacillus azotofixans (Paenibacillus durus, ID ATCC 35681)—and Bacillaceae family—Bacillus coagulans (ID 941639) and Bacillus pumilus (ID ATCC 7061) [29]. Plant growth-promoting bacteria were isolated and extracted from the commercial product Bactel (Bioera SL-Spain). They were applied during three growth stages (transplant, flowering, and pre-harvest) in each crop at 100 mL from a suspension of 50 g L−1 PGPR. We applied 100 mL to each plant (container) at 10 cm around the plant. This was equivalent to 2.5 × 1010 CFU per plant [29] at each application.
- Trichoderma sp. (TRICH) consortium: This treatment contained a mix of Trichoderma asperellum (ID CBS 433.97) 0.5% g 100 g−1 (w/w), 1 × 108 CFU g−1 and Trichoderma atroviride (ID IMI206040) 0.5% w/w, 1 × 108 CFU g−1 [30]. Isolates of Trichoderma fungi were isolated and extracted from the commercial product Tusal (CERTIS-Spain). These were applied during three growth stages (transplant, flowering, and pre-harvest) in each crop at 100 g TRICH concentration diluted in 1 L−1 of water, and 100 mL of this mixture was applied to each plant (container) at 10 cm around the plant. This was equivalent to 109 CFU plant−1 [30] at each application.
2.3. Irrigation Management and Nutrition
2.4. Crop Yield
2.5. Dehydrogenase Activity (DHA)
2.6. Data Analysis
3. Results
3.1. Yield: PGPMs and Substrates in Each Crop (Melon, Tomatoes)
3.2. Petiole Sap Test
3.3. Drainage Test
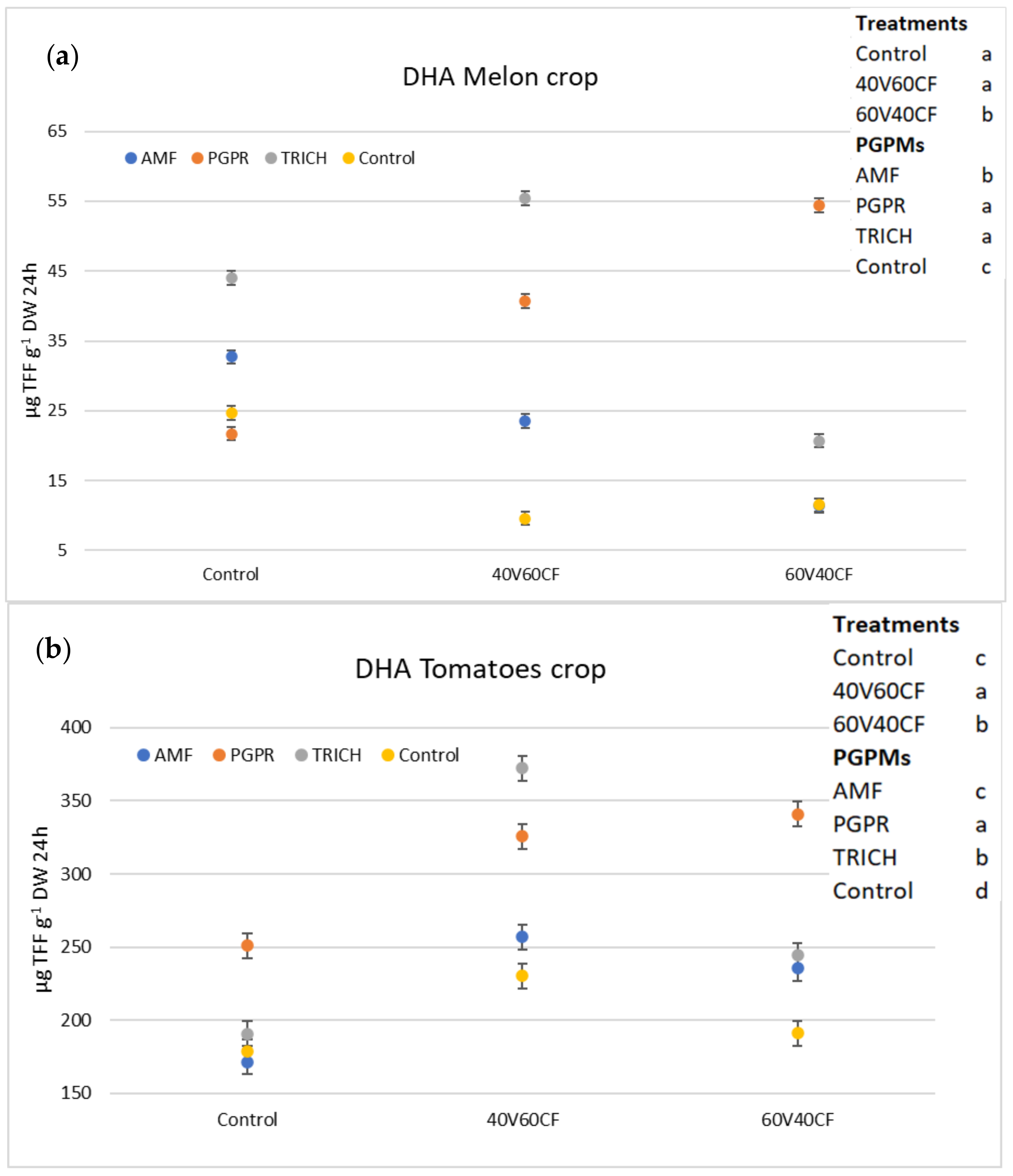
3.4. Dehydrogenase Activity (DHA)
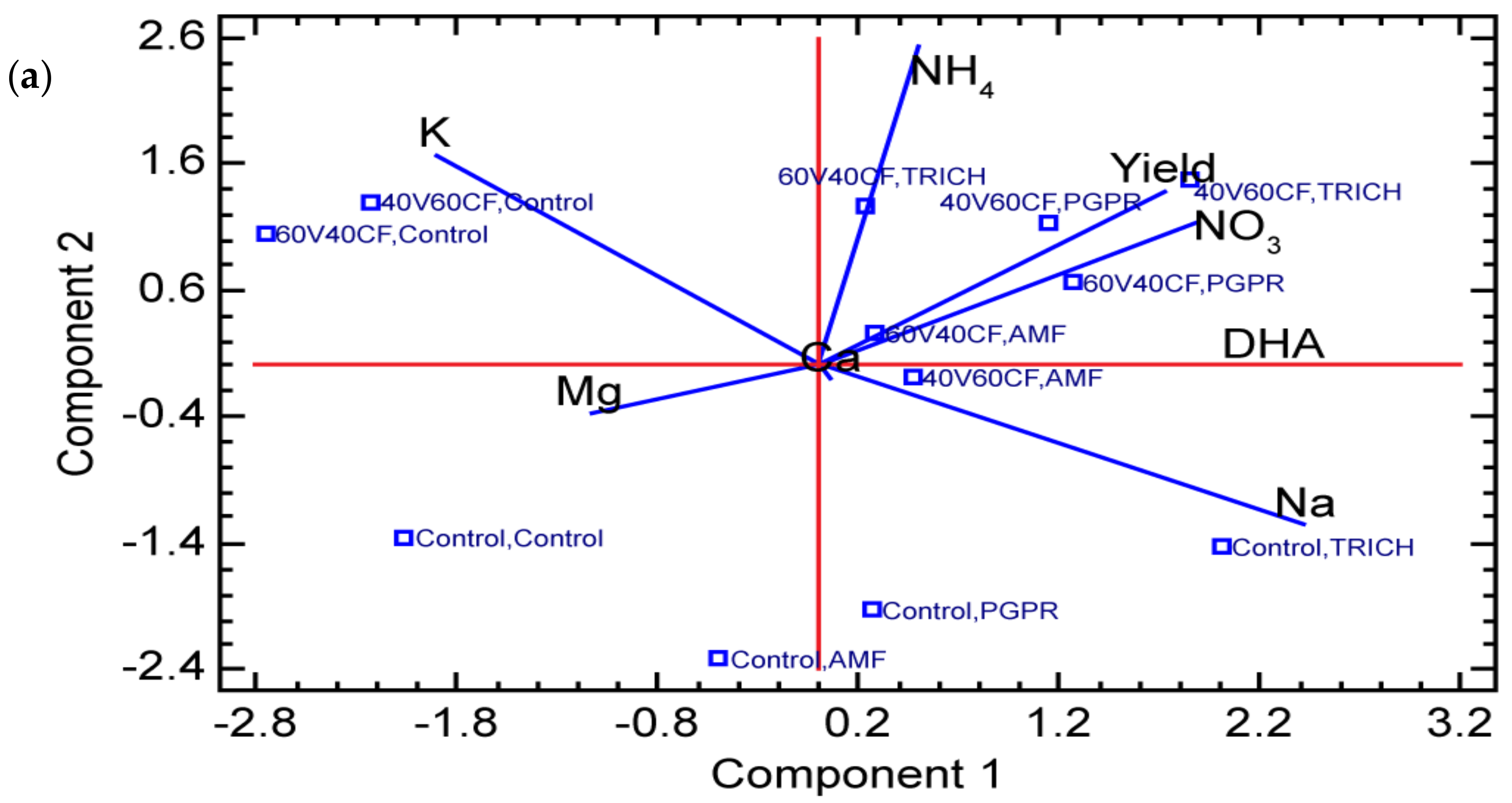
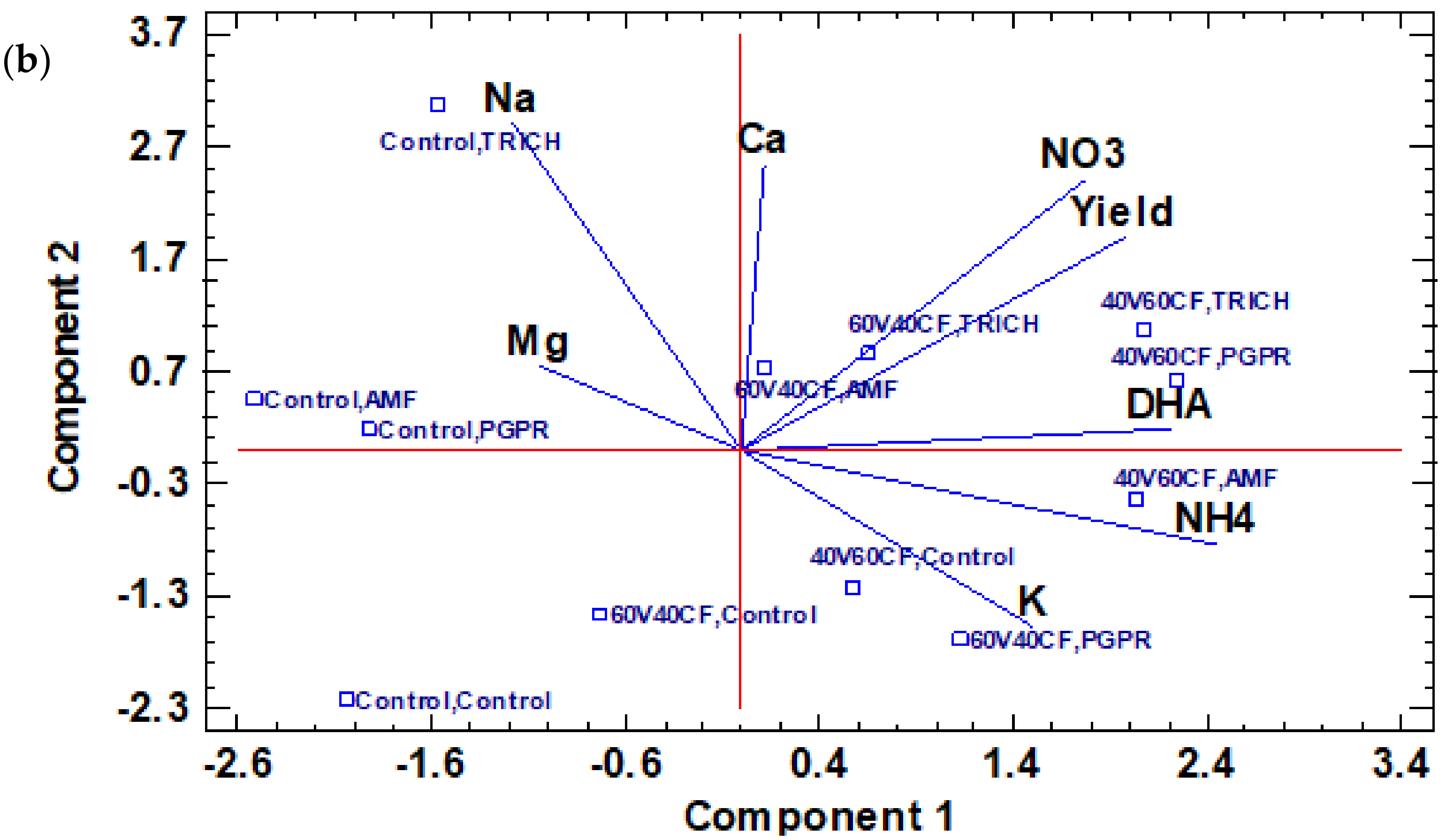
3.5. Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rogers, M.A. Organic vegetable production in controlled environments using soilless media. Hortechnology 2017, 27, 166–170. [Google Scholar] [CrossRef] [Green Version]
- Gohardoust, M.R.; Bar-Tal, A.; Effati, M.; Tuller, M. Characterization of physicochemical and hydraulic properties of organic and mineral soilless culture substrates and mixtures. Agronomy 2020, 10, 1403. [Google Scholar] [CrossRef]
- Ruiz, J.; Salas, M.C. Evaluation of organic substrates and microorganisms as bio-fertilisation tool in container crop production. Agronomy 2019, 9, 705. [Google Scholar] [CrossRef] [Green Version]
- Hussain, N.; Singh, A.; Saha, S.; Venkata, M.; Bhattacharyya, P.; Bhattacharya, S.S. Excellent N-fixing, and P-solubilizing traits in earthworm gut-isolated bacteria: A vermicompost based assessment with vegetable market waste and rice straw feed mixtures. Bioresour. Technol. 2016, 222, 165–174. [Google Scholar] [CrossRef]
- Loera, A.; Troyo, E.; Murillo, B.; Barraza, A.; Caamal-Chan, G.; Lucero, G.; Nieto, A. Effects of Vermicompost Leachate versus Inorganic Fertilizer on Morphology and Microbial Traits in the Early Development Growth Stage in Mint (Mentha spicata L.) and Rosemary (Rosmarinus officinalis L.) Plants under Closed Hydroponic System. Horticulturae 2021, 7, 100. [Google Scholar] [CrossRef]
- Saeed, Q.; Xiukang, W.; Haider, F.U.; Kuˇcerik, J.; Mumtaz, M.Z.; Holatko, J.; Naseem, M.; Kintl, A.; Ejaz, M.; Naveed, M.; et al. Rhizosphere Bacteria in Plant Growth Promotion, Biocontrol, and Bioremediation of Contaminated Sites: A Comprehensive Review of Effects and Mechanisms. Int. J. Mol. Sci. 2021, 22, 10529. [Google Scholar] [CrossRef]
- Khan, N.; Ali, S.; Shahid, M.A.; Mustafa, A.; Sayyed, R.Z.; Curá, J.A. Insights into the Interactions among Roots, Rhizosphere, and Rhizobacteria for Improving Plant Growth and Tolerance to Abiotic Stresses: A Review. Cells 2021, 10, 1551. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Naveed, M.; Mustafa, A.; Abbas, A. The good, the bad, and the ugly of rhizosphere microbiome. In Probiotic and Plant Health; Springer: Singapore, 2017; pp. 253–290. [Google Scholar] [CrossRef]
- Shutz, L.; Gattinger, A.; Meier, M.; Muller, A.; Boller, T.; Mader, P.; Mathimaran, N. Improving crop yield and nutrient use efficiency via biofertilization—A global meta-analysis. Front. Plant Sci. 2017, 8, 2204. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.T.; Hwang, S.G.; Huang, Y.M.; Huang, C.H. Effects of Trichoderma asperellum on nutrient uptake and Fusarium wilt of tomato. Crop Prot. 2018, 110, 275–282. [Google Scholar] [CrossRef]
- Sousa, L.B.; Stamford, N.P.; Oliveira, W.S.; Silva, E.V.N.; Santos, C.E.R.S.; Stamford, T.C.M. Optimization of biofertilizers enriched in N by diazotrophic bacteria. Eur. J. Sustain. Dev. 2017, 6, 448–456. [Google Scholar] [CrossRef]
- Imade, E.E.; Babalola, O.O. Biotechnological utilization: The role of Zea mays rhizospheric bacteria in ecosystem sustainability. Appl. Microbiol. Biotechnol. 2021, 105, 4487–4500. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.; Salas, M.C. The use of plant growth promoting bacteria for biofertigation; effects on concentrations of nutrients in the inoculated aqueous vermicompost extract and on the yield and quality of tomatoes. Biol. Agric. Hortic. 2022, 1–17. [Google Scholar] [CrossRef]
- Abhilash, P.C.; Dubey, R.K.; Tripathi, V.; Gupta, V.K.; Singh, H.B. Plant growth-promoting microorganisms for environmental sustainability. Trends Biotechnol. 2016, 34, 847–850. [Google Scholar] [CrossRef] [PubMed]
- Caporale, A.G.; Vitaglione, P.; Troise, A.D.; Pigna, M.; Ruocco, M. Influence of three different soil types on the interaction of two strains of Trichoderma harzianum with Brassica rapa subsp. sylvestris cv. esculenta, under soil mineral fertilization. Geoderma 2019, 350, 11–18. [Google Scholar] [CrossRef]
- Langendries, S.; Goormachtig, S. Paenibacillus polymyxa, a Jack of all trades. Environ. Microbiol. 2021, 23, 5659–5669. [Google Scholar] [CrossRef]
- Bhardwaj, A.; Sharma, A.; Goswami, B.K.; Bhardwaj, V. Interaction Effect of Soilless Media and Organic Amendments for Eco-Friendly Root-Knot Nematode Management in Brinjal and Tomato Nursery. J. Pure Appl. Microbiol. 2021, 15, 356–367. [Google Scholar] [CrossRef]
- Owen, D.; Williams, A.P.; Griffith, G.W.; Withers, P.J.A. Use of commercial bio-inoculants to increase agricultural production through improved phosphorus acquisition. Appl. Soil Ecol. 2015, 86, 41–54. [Google Scholar] [CrossRef]
- Constable, G.A.; Rochester, I.J.; Betts, J.H.; Herridge, D.F. Prediction of nitrogen fertilizer requirement in cotton using petiole and sap nitrate. Commun. Soil Sci. Plant Anal. 1991, 22, 1315–1324. [Google Scholar] [CrossRef]
- Pardossi, A.; Incrocci, L.; Salas, M.C.; Gianquinto, G. Managing Mineral Nutrition in Soilless Culture. In Rooftop Urban Agriculture; Orsini, F., Dubbeling, M., de Zeeuw, H., Gianquinto, G., Eds.; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Saia, S.; Rappa, V.; Ruisi, P.; Abenavoli, M.R.; Sunseri, F.; Giambalvo, D.; Frenda, A.S.; Martinelli, F. Soil inoculation with symbiotic microorganisms promotes plant growth and nutrient transporter genes expression in durum Wheat. Front. Plant Sci. 2015, 6, 815. [Google Scholar] [CrossRef] [Green Version]
- Esparza-Reynoso, S.; Ruiz-Herrera, F.; Pelagio-Flores, R.; Macías-Rodríguez, L.I.; Martínez-Trujillo, M.; López-Coria, M.; Sánchez-Nieto, S.; Herrera-Estrella, A.; López-Bucio, J. Trichoderma atroviride emitted volatiles improve growth of Arabidopsis seedlings through modulation of sucrose transport and metabolism. Plant Cell Environ. 2021, 44, 1961–1976. [Google Scholar] [CrossRef]
- Bernal-Vicente, A.; Pascual, J.A.; Tittarelli, F.; Hernández, J.A.; Diaz-Vivancos, P. Trichoderma harzianum T-78 supplementation of compost stimulates the antioxidant defense system in melon plants. J. Sci. Food Agric. 2015, 95, 2208–2214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalska, J.; Remlein-Starosta, J.; Seider-Lozykowska, K. Can Trichoderma asperellum (T1) stimulate growth of lemon balm (Melissa officinalis L.) in different systems of cultivation. Acta Sci. Pol. Hortorum Cultus 2014, 13, 91–102. [Google Scholar]
- Camacho Ferre, F. Técnicas de Producción en Cultivos Protegidos (No. 04; SB318 T4.); Caja Rural Intemediterránea: Cajamar, Brazil, 2003. [Google Scholar]
- Prasad, M.; Maher, M.J. The use of composted green waste (CGW) as a growing medium component. Acta Hortic. 2001, 549, 107–114. [Google Scholar] [CrossRef]
- Truong, H.D.; Wang, C.H. Studies on the effects of vermicompost on physicochemical properties and growth of two tomato varieties under greenhouse conditions. Commun. Soil Sci. Plant Anal. 2015, 46, 1494–1506. [Google Scholar] [CrossRef]
- Ramadhani, I.; Widawati, S.; Sudiana, I.M.; Kobayashi, M. The effect of the combination of arbuscular mycorrhiza and rhizobacteria and doses of NPK fertilizer on the growth of Sorghum bicolor (L.) Moench. IOP Conf. Ser. Earth Environ. Sci. 2019, 308, 12045. [Google Scholar] [CrossRef]
- Basavesha, K.N.; Savalgi, V.P. Effect of nitrogen fixing paenibacillus SP. Isolates on growth, yield, and nutrient uptake on maize in calcareous soil. Int. J. Agric. Stat. Sci. 2018, 14, 427–431. [Google Scholar]
- Márquez-Benavidez, L.; Rizo-León, M.Á.; Montaño-Arias, N.M.; Ruiz-Nájera, R.; Sánchez-Yáñez, J.M. Respuesta de Phaseolus vulgaris a la inoculación de diferentes dosis de Trichoderma harzianum con el fertilizante nitrogenado reducido al 50%. J. Selva Andin. Res. Soc. 2017, 8, 135–144. [Google Scholar] [CrossRef] [Green Version]
- López-González, J.A.; López, M.J.; Vargas-García, M.C.; Suárez-Estrella, F.; Jurado, M.; Moreno, J.Y. Tracking organic matter and microbiota dynamics during the stages of lignocellulosic waste composting. Bioresour. Technol. 2013, 146, 574–584. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of phosphorus in soil in connection with the vital activity of some microbial species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Wilson, P.W.; Knight, S.C. Experiments in Bacterial Physiology; Burgess: Minneapolis, MN, USA, 1952; p. 49. [Google Scholar]
- Gonzalez, K.D.; Rodriguez, M.N.; Trejo, L.I.; Garcia, J.L.; Sanchez, J. Vermicompost effluent and tea in the production of leafy vegetables in system NFT. Interciencia 2013, 38, 863–869. [Google Scholar]
- Salas, M.C.; Mejía, P.A.; Domínguez, N.; Ruiz, J.L. Evaluating the efficiency of two automatic fertigation systems in soilless crops: Substrate moisture sensors vs. timer systems. Acta Hortic. 2020, 1273, 23–30. [Google Scholar] [CrossRef]
- Bodale, I.; Mihalache, G.; Achitei, V.; Teliban, G.C.; Cazacu, A.; Stoleru, V. Evaluation of the nutrients uptake by Tomato plants in different phenological stages using an electrical conductivity technique. Agriculture 2021, 11, 292. [Google Scholar] [CrossRef]
- Bityutskii, N.; Yakkonen, K.; Petrova, A.; Nadporozhskaya, M. Xylem sap mineral analyses as a rapid method for estimation plant-availability of Fe, Zn, and Mn in carbonate soils: A case study in cucumber. J. Soil Sci. Plant Nutr. 2017, 17, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Hochmuth, G.; Maynard, D.; Vavrina, C.; Hanlon, E. Plant Tissue Analysis and Interpretation for Vegetable Crops in Florida. Fla. Horticultural Sciences. 1991. Department, UF/IFAS Extension. Original Publication Date October 1994. Reviewed April 2015. Available online: http://edis.ifas.ufl.edu (accessed on 15 January 2020).
- Garcia, C.; Hernandez, T.; Costa, F. Potential use of dehydrogenase activity as an index of microbial activity in degraded soils. Commun. Soil Sci. Plant Anal. 2008, 28, 123–134. [Google Scholar] [CrossRef]
- Batra, L.; Manna, M.C. Dehydrogenase activity and microbial biomass carbon in salt-affected soils of semiarid and arid regions. Arid Soil Res. Rehabil. 1997, 11, 295–303. [Google Scholar] [CrossRef]
- Casida, L.E. Microbial metabolic activity in soils as measured by dehydrogenase determinations. Appl. Environ. Microbiol. 1977, 34, 630–636. [Google Scholar] [CrossRef] [Green Version]
- Vozhehova, R.; Kokovikhin, S.; Lykhovyd, P.V.; Balashova, H.; Lavrynenko, Y.; Biliaieva, I.; Markovska, O. Statistical yielding models of some irrigated vegetable crops in dependence on water use and heat supply. J. Water Land Dev. 2020, 45, 190–197. [Google Scholar] [CrossRef]
- Vandecasteele, B.; Willekens, K.; Van Delm, T. Changes in chemical and physical properties of compost-amended growing media during strawberry cropping: Monitoring depletion of nutrients with petiole sap measurements. In Proceedings of the 1st International Symposium on Organic Matter Management and Compost Use in Horticulture, Adelaide, Australia, 4–7 April 2011; Volume 1018, pp. 541–549. [Google Scholar]
- Das, D.; Bhattacharyya, P.; Ghosh, B.C.; Banik, P. Bioconversion and biodynamics of Eisenia foetida in different organic wastes through microbially enriched vermiconversion technologies. Ecol. Eng. 2016, 86, 154–161. [Google Scholar] [CrossRef]
- Cadahía, C. La savia como índice de fertilización, cultivos agro-energéticos. In Horticolas, Frutales y Ornamentals; Mundi-Prensa: Madrid, Spain, 2008. [Google Scholar]
- Zulkarami, B.; Ashrafuzzman, M.; Razi, I.M. Morpho-physiological growth, yield, and fruit quality of rock melon as affected by growing media and electrical conductivity. J. Food Agric. Environ. 2010, 8, 249–252. [Google Scholar]
- Wang, X.X.; Zhao, F.; Zhang, G.; Zhang, Y.; Yang, L. Vermicompost improves tomato yield and quality and the biochemical properties of soils with different tomato planting history in a greenhouse study. Front. Plant Sci. 2017, 21, 1978. [Google Scholar] [CrossRef] [Green Version]
- Barrett, G.E.; Alexander, P.D.; Robinson, J.S.; Bragg, N.C. Achieving environmentally sustainable growing media for soilless plant cultivation systems—A review. Sci. Hortic. 2016, 212, 220–234. [Google Scholar] [CrossRef] [Green Version]
- Vargas, M.C.; Suárez, F.; López, M.J.; Moreno, J. Microbial population dynamics and enzyme activities in composting processes with different starting materials. Waste Manag. 2010, 30, 771–778. [Google Scholar] [CrossRef] [PubMed]
- EPA. Estimated Nitrate Concentrations in Groundwater Used for Drinking. Available online: https://www.epa.gov/nutrient-policy-data/estimated-nitrate-concentrations-groundwater-used-drinking (accessed on 28 March 2022).
- Mal, S.; Chattopadhyay, G.N.; Chakrabarti, K. Microbiological integration for quantitative improvement of vermicompost. Int. J. Recycl. Org. Waste Agric. 2021, 10, 157–166. [Google Scholar] [CrossRef]
- Dominguez, S.; Belen Rubio, M.; Cardoza, R. Nitrogen metabolism and growth enhancement in tomato plants challenged with Trichoderma harzianum expressing the Aspergillus nidulans acetamidase amdS gene. Front. Microbiol. 2016, 7, 1182. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.E.; Doni, F.; Khadka, R.B.; Uphoff, N. Endophytic strains of Trichoderma increase plants’ photosynthetic capability. J. Appl. Microbiol. 2021, 130, 529–546. [Google Scholar] [CrossRef]
- Kumar, K.; Manigundan, K.; Amaresan, N. Influence of salt tolerant Trichoderma spp. On growth of maize (Zea mays) under different salinity conditions. J. Basic Microbiol. 2017, 57, 141–150. [Google Scholar] [CrossRef]
- Pandey, V.; Ansari, M.W.; Tula, S. Dose -dependent response of Trichoderma harzianum in improving drought tolerance in rice genotypes. Planta 2016, 243, 1251–1264. [Google Scholar] [CrossRef]
- Pascale, A.; Vinale, F.; Manganiello, G. Trichoderma and its secondary metabolites improve yield and quality of grapes. Crop Prot. 2017, 92, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Matata, M.; Cibulova, A.; Varecka, L. Plant waste residues as inducers of extracellular proteases for a deuteromycete fungus Trichoderma atroviridae. Chem. Pap. 2016, 70, 1039–1048. [Google Scholar] [CrossRef]
- Ruting, G.; Ji, S.; Wang, Z.; Zhang, H.; Wang, Y.; Liu, Z. Trichoderma asperellum xylanases promote growth and induce resistance in poplar. Microbiol. Res. 2021, 248, 126767. [Google Scholar]
- Muniswami, D.M.; Buvaneshwari, K.; Fathima Rosa Mystica, L.; Naveena, T.; Pabitha, B.; Reshma, S.; Rangila, D.; Santhiya, P.; Sharmila Devi, N.; Ahamed Rasheeq, A.; et al. Comparative assessment of different biofertilizers in maize (Zea mays L.) cultivation. Biomass Convers. Biorefinery 2021, 1–19. [Google Scholar] [CrossRef]
- Tao, R.; Wakelin, S.A.; Liang, Y.; Chu, G. Response of ammonia-oxidizing archaea and bacteria in calcareus soil to mineral and organic fertilizer application and their relative contribution to nitrification. Soil Biol. Biochem. 2017, 114, 20–30. [Google Scholar] [CrossRef]
- Lang, H.J.; Elliot, G.C. Enumeration, and inoculation of nitrifying bacteria in soilless potting media. J. Am. Soc. Hortic. Sci. 1997, 122, 709–714. [Google Scholar] [CrossRef] [Green Version]
- Djukic, D.; Mandic, L.; Moracanin, S.V.; Aleksandra, S. Effect of mineral fertilizers and biofertilizers on soil microbial characteristics and Potato yield. In Proceedings of the 14th International Multidisciplinary Scientific Geoconference Sgem, Albena, Bulgaria, 17–26 June 2014; pp. 343–349. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| March | April | May | June |
|---|---|---|---|
| 14.2 | 16.8 | 19.3 | 22.8 |
| September | October | November | December | January | February | March |
|---|---|---|---|---|---|---|
| 25.1 | 21.9 | 16.1 | 14.2 | 12 | 14 | 15 |
| Parameters | Units | CF | V |
|---|---|---|---|
| pH | 5.8–6.8 | 7.73 | |
| Electric conductivity (E.C.) | dS m−1 | <0.7 | 0.9 |
| Particle size | mm | 0–12 | <5 |
| Solid particle density | g cm−3 | 0.1 | 0.77 |
| Total porosity | % | 95.4 | 67.9 |
| Cation exchange capacity | mmol 100 g−1 | 60–130 | 25–30 |
| Organic matter | % DM 1 | 94.7 | 15 |
| Sodium | mg L−1 | 253 | 431 |
| Potassium | mg L−1 | 108 | 1881 |
| Calcium | mg L−1 | 29 | 506 |
| Magnesium | mg L−1 | 28 | 192 |
| Chloride | mg L−1 | 281 | 1260 |
| Sulphates | mg L−1 | 321 | 1988 |
| Nitrates | mg L−1 | 120 | 1312 |
| Phosphates | mg L−1 | 32 | 25 |
| Sodium adsorption ratio | 11.4 | 4.1 | |
| Organic carbon | g kg−1 | 78.6 | 82.4 |
| Organic nitrogen | g kg−1 | 1.8 | 9.9 |
| C: N Ratio | 43.66 | 8.32 | |
| Humic and Fulvic acids | % w/w 2 | ND 3 | 17.50 |
| INPUTS | Units | TB * | TF * | NF * | PS * |
|---|---|---|---|---|---|
| AMF 1 | Log (CFU g−1 dw **) | 5.88 ± 0.03 | 4.92 ± 0.08 | 5.62 ± 0.06 | 5.34 ± 0.13 |
| PGPR 2 | Log (CFU g−1 dw) | 9.23 ± 0.31 | 3.77 ± 0.33 | 9.05 ± 0.27 | 0.00 |
| TRICH 3 | Log (CFU g−1 dw) | 4.14 ± 0.29 | 7.65 ± 0.50 | 0.00 | 0.00 |
| VT | Log (CFU mL−1 ***) | 4.93 ± 0.42 | 1.41 ± 0.33 | 4.83 ± 0.45 | 4.74 ± 0.06 |
| YIELD Kg m−2 | Control | AMF | PGPR | TRICH |
|---|---|---|---|---|
| Melon | ||||
| Control | 4.27 a A | 4.31 a A | 5.03 * B | 5.18 a B |
| 40V60CF | 4.75 b A | 5.43 b B | 4.98 * B | 5.51 b B |
| 60V40CF | 4.92 b A | 5.58 b B | 5.08 * A | 5.55 b B |
| Tomato | ||||
| Control | 5.03 a A | 5.12 a A | 5.08 a A | 5.90 b B |
| 40V60CF | 5.74 b B | 5.96 ab B | 5.96 ab B | 6.12 b B |
| 60V40CF | 5.21 a A | 5.79 b B | 5.27 a A | 5.43 a A |
| 20V80CF | 40V60CF | 60V40CF | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | AMF | PGPR | TRICH | Control | AMF | PGPR | TRICH | Control | AMF | PGPR | TRICH | |
| NO3− | 520.1 a A | 538.3 a A | 523.7 a A | 817.9 b B | 658.3 b B | 546.0 a A | 874.2 c D | 785.1 ab C | 488.7 a A | 683.7 b C | 593.2 b B | 767.3 a D |
| NH4+ | 108.3 a B | 98.3 a A | 100.3 a A | 105.2 a AB | 131.0 b A | 132.8 c A | 139.8 b B | 146.2 b B | 134.0 c B | 121.8 b A | 147.2 b C | 144.7 b C |
| Mg2+ | 127.2 b C | 92.8 b B | 74.0 b A | 124.2 b C | 87.0 a B | 73.2 a A | 70.0 b A | 79.2 a AB | 120.0 b C | 73.2 a B | 56.8 a A | 134.0 b D |
| K+ | 2018.8 a B | 1937.2 a AB | 1731.7 a A | 1864.1 a A | 2524.8 b C | 1752.0 a A | 2074.9 b B | 2092.8 b B | 2601.9 b C | 1921.7 a A | 2016.1 b AB | 2147.7 b B |
| Ca2+ | 362.2 a A | 581.2 a C | 424.8 a B | 552.9 b C | 513.3 b C | 520.0 a C | 403.2 a A | 459.3 a B | 510.1 b B | 539.2a B | 425.1 a A | 555.2 b B |
| Na+ | 184.0 b A | 331.7 a B | 343.2 ab B | 466.8 b C | 131.8 a A | 269.7 a B | 272.0a B | 301.2 a B | 163.0 b A | 259.2 a B | 308.7 b C | 296.8 a C |
| 20V80CF | 40V60CF | 60V40CF | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | AMF | PGPR | TRICH | Control | AMF | PGPR | TRICH | Control | AMF | PGPR | TRICH | |
| NO3− | 1138.6 a A | 1178.6 a A | 1365.6 a B | 1571.8 a C | 1392.7 c A | 1571.6 b B | 1600.3 b B | 1705.4 b B | 1070.0 b A | 1497.0 b C | 1298.8 b B | 1680.0 b D |
| NH4+ | 152.0 a * | 137.0 a * | 140.0 a * | 147.0 a * | 179.3 b * | 195.7 c * | 190.6 b * | 182.5 b * | 177.0 b * | 167.2 b * | 201.9 b * | 198.5 b * |
| Mg2+ | 553.7 b C | 404.1 b B | 322.1 b A | 540.5 b C | 339.5 a B | 333.7 a B | 281.9 a A | 391.3 a C | 348.2 a A | 362.0 a A | 377.9 c A | 583.3 a B |
| K+ | 3396.4 a B | 3259.1 a B | 2913.4 a A | 3136.2 a AB | 3520.8 ab AB | 3879.8 b C | 3566.9 b B | 3239.7 a A | 3704.6 b B | 3233.1 a A | 3476.0 b A | 3613.3 b AB |
| Ca2+ | 608.7 a A | 976.8 c C | 713.9 a B | 929.3 b C | 641.1 a A | 778.7 a B | 945.3 b C | 821.0 a B | 857.3 b B | 906.2 b BC | 714.4 a A | 933.1 b C |
| Na+ | 770.0 a A | 1387.9 b B | 1436.0 c B | 1953.5 c C | 940.6 ab A | 1041.8 a B | 1049.8 b B | 1046.2 a B | 1100.6 b B | 1084.5 a B | 873.2 a A | 1242.1 b B |
| (a) | |||||||
| Factor | NO3−-N mg L−1 | NH4+ mg L−1 | Mg2+ mg L−1 | K+ mg L−1 | Cl− mg L−1 | Na+ mg L−1 | Ca2+ mg L−1 |
| Substrate | |||||||
| 20V80CF | 1.76 a | 25.44 a | 29.65 a | 213.09 a | 255.62 a | 203.69 a | 298.18 a |
| 40V60CF | 1.30 a | 11.18b | 42.78 a | 260.79 a | 394.59 ab | 269.67 ab | 376.73 ab |
| 60V40CF | 1.44 a | 11.37 b | 41.32 a | 270.95 a | 470.46 b | 297.03 b | 460.9 b |
| DAT | |||||||
| 30 | 1.07 a | 17.14 b | 81.66 c | 467.23 a | 485.00 b | 231.97 ab | 368.72 a |
| 60 | 0.94 a | 9.02 a | 38.65 b | 199.4 b | 267.67 a | 217.25 a | 380.74 a |
| 75 | 2.27 b | 37.53 c | 13.61 a | 236.94 b | 412.67 b | 315.19 b | 416.01 a |
| 90 | 3.03 c | 12.27 ab | 25.28 ab | 255.31 b | 748.77 c | 420.71 c | 369.92 a |
| (b) | |||||||
| Factor | NO3−-N mg L−1 | NH4+ mg L−1 | Mg2+ mg L−1 | K+ mg L−1 | Cl− mg L−1 | Na+ mg L−1 | Ca2+ mg L−1 |
| Substrate | |||||||
| 20V80CF | 1.63 b | 7.38 a | 8.02 a | 327.57 a | 229.36 a | 152.42 a | 69.60 b |
| 40V60CF | 4.48 a | 7.56 a | 11.66 a | 230.63 a | 199.58 a | 203.69 a | 119.60 a |
| 60V40CF | 3.83 a | 7.20 a | 9.48 a | 210.69 a | 263.39 a | 238.17 a | 144.00 a |
| DAT | |||||||
| 30 | 2.71 b | 4.5 b | 0.24 b | 427.25 a | 263.39 bc | 78.16 c | 199.20 ab |
| 60 | 2.21 b | 10.98 a | 40.09 a | 392.46 a | 180.08 d | 222.54 b | 231.20 a |
| 90 | 5.85 a | 7.2 ab | 1.46 b | 345.94 a | 246.73 bcd | 159.78 b | 106.00 c |
| 120 | 5.12 a | 11.88 a | 0 b | 343.60 a | 341.38 a | 228.29 b | 131.60 bc |
| 150 | 8.57 a | 3.96 ab | 53.95 a | 87.17 b | 146.05 cd | 142.53 bc | 150.00 abc |
| 180 | 6.70 a | 1.8 b | 1.46 b | 58.24 b | 275.80 ab | 303.47 a | 110.00 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mejía, P.A.; Ruíz-Zubiate, J.L.; Correa-Bustos, A.; López-López, M.J.; Salas-Sanjuán, M.d.C. Effects of Vermicompost Substrates and Coconut Fibers Used against the Background of Various Biofertilizers on the Yields of Cucumis melo L. and Solanum lycopersicum L. Horticulturae 2022, 8, 445. https://doi.org/10.3390/horticulturae8050445
Mejía PA, Ruíz-Zubiate JL, Correa-Bustos A, López-López MJ, Salas-Sanjuán MdC. Effects of Vermicompost Substrates and Coconut Fibers Used against the Background of Various Biofertilizers on the Yields of Cucumis melo L. and Solanum lycopersicum L. Horticulturae. 2022; 8(5):445. https://doi.org/10.3390/horticulturae8050445
Chicago/Turabian StyleMejía, Pedro A., José Luis Ruíz-Zubiate, Amelia Correa-Bustos, María José López-López, and María del Carmen Salas-Sanjuán. 2022. "Effects of Vermicompost Substrates and Coconut Fibers Used against the Background of Various Biofertilizers on the Yields of Cucumis melo L. and Solanum lycopersicum L." Horticulturae 8, no. 5: 445. https://doi.org/10.3390/horticulturae8050445
APA StyleMejía, P. A., Ruíz-Zubiate, J. L., Correa-Bustos, A., López-López, M. J., & Salas-Sanjuán, M. d. C. (2022). Effects of Vermicompost Substrates and Coconut Fibers Used against the Background of Various Biofertilizers on the Yields of Cucumis melo L. and Solanum lycopersicum L. Horticulturae, 8(5), 445. https://doi.org/10.3390/horticulturae8050445