
Rhizosphere Acidification as the Main Trait Characterizing the Differential In Vitro Tolerance to Iron Chlorosis in Interspecific Pyrus Hybrids

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
Statistical Analysis of the Results
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bunnag, S.; Dolcet-Sanjuan, R.; Mok, D.W.S.; Mok, M.C. Responses of two somaclonal variants of quince (Cydonia oblonga) to iron deficiency in the greenhouse and field. J. Am. Soc. Hortic. Sci. 1996, 121, 1054–1058. [Google Scholar] [CrossRef] [Green Version]
- Dolcet-Sanjuan, R.; Claveria, E.; Bonany, J.; Iglesias, I.; Asín, L.; Simard, M.H. Selection for new pear rootstocks: In vitro screening and field evaluation for tolerance to iron chlorosis. Acta Hortic. 2004, 658, 463–468. [Google Scholar] [CrossRef]
- Gonzalo, M.J.; Moreno, M.A.; Gogorcena, Y. Physiological responses and differential gene expression in Prunus rootstocks under iron deficiency conditions. J. Plant Physiol. 2011, 168, 887–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iglesias, I.; Vilardell, P.; Bonany, J.; Claveria, E.; Dolcet-Sanjuan, R. Micropropagation and field evaluation of the pear (Pyrus communis L.) ‘IGE 2002’, a new selection of the cultivar Dr. Jules Guyot. J. Am. Soc. Hortic. Sci. 2004, 129, 389–393. [Google Scholar] [CrossRef]
- Tagliavini, M.; Rombola, A.D.; Marangoni, B. Response to iron-deficiency stress of pear and quince genotypes. J. Plant Nutr. 1995, 18, 2465–2482. [Google Scholar] [CrossRef]
- Tagliavini, M.; Rombolá, D. Iron deficiency and chlorosis in orchard and vineyard ecosystems. Eur. J. Agron. 2001, 15, 71–92. [Google Scholar] [CrossRef]
- Pestana, M.; de Varennes, A.; Araújo Faria, E. Diagnosis and correction of iron chlorosis in fruit trees: A review. Food Agric. Environ. 2003, 1, 46–51. [Google Scholar]
- Chaney, R.L. Diagnostic practices to identify iron deficiency in higher plants. J. Plant Nutr. 1984, 7, 47–67. [Google Scholar] [CrossRef]
- Donnini, S.; Castagna, A.; Ranieri, A.; Zocchi, G. Differential responses in pear and quince genotypes induced by Fe deficiency and bicarbonate. J. Plant Physiol. 2009, 166, 1181–1193. [Google Scholar] [CrossRef]
- Gogorcena, Y.; Abadía, J.; Abadía, A. A new technique for screening iron-efficient genotypes in peach rootstocks: Elicitation of root ferric chelate reductase by manipulation of external iron concentrations. J. Plant Nutr. 2004, 27, 1710–1715. [Google Scholar] [CrossRef]
- Jiménez, S.; Pinochet, J.; Abadía, A.; Moreno, M.A.; Gogorcena, Y. Tolerance responses to iron chlorosis of Prunus selections as rootstocks. HortScience 2008, 43, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Simard, M.H. French contribution to rootstocks breeding and evaluation. In Proceedings of the International Conference on Fruit Tree Rootstocks, Pisa, Italy, 26 June 2009; pp. 33–43. [Google Scholar]
- Dolcet-Sanjuan, R.; Claveria, E.; Vilardell, P.; Bonany, J.; Asin, L.; Simard, M.H. Towards the Selection of a New Pear Rootstock: In Vitro and Field Evaluation for Tolerance to Iron Chlorosis, Low Vigor and Micropropagation of Selected Clones. Acta Hortic. 2008, 800, 683–691. [Google Scholar] [CrossRef]
- Römheld, V.; Marschner, H. Mobilization of iron in the rhizosphere of different plant species. In Advances in Plant Nutrition; Tinker, A., Läuchli, A., Eds.; Praeger Scientific: New York, NY, USA, 1986; pp. 155–204. [Google Scholar]
- Li, H.; Wang, L.; Yang, Z.M. Co-expression analysis reveals a group of genes potentially involved in regulation of plant responses to iron-deficieny. Gene 2015, 554, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, W. Iron stress responses in roots of strategy I plants. In Iron Nutrition in Plants and Rhizospheric Microorganisms; Barton, L.L., Abadía, J., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 229–250. [Google Scholar]
- Zocchi, G. Metabolic changes in iron-stressed dicotyledoneous plants. In Iron Nutrition in Plants and Rhizospheric Microorganisms; Barton, L.L., Abadía, J., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 359–370. [Google Scholar]
- Satbhai, S.; Setzer, C.; Freynschlag, F.; Slovak, R.; Kerdaffrec, E.; Busch, W. Natural allelic variation of FRO2 modulates Arabidopsis root growth under iron deficiency. Nat. Commun. 2017, 8, 15603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simard, M.H.; Guisnel, R.; Demilly, D.; Billy, B.; Honoré, D.; Daguin, F. Is dwarfing in pear rootstocks due to aneuploïd genetic structures? Acta Hortic. 2011, 909, 59–66. [Google Scholar] [CrossRef]
- Asin, L.; Iglesias, I.; Dolcet-Sanjuan, R.; Claveria, E.; Vilardell, P.; Bonany, J.; Simard, M.H. INRA-IRTA Pear Rootstock Breeding Program: Aiming for Tolerance to Iron Chlorosis. In IX International Symposium on Integrating Canopy, Rootstock and Environmental Physiology in Orchard Systems 903; Robinson, T.L., Ed.; ISHS: Leuven, Belgium, 2011; pp. 207–213. [Google Scholar] [CrossRef]
- Dolcet-Sanjuan, R.; Mok, D.W.S.; Mok, M.C. Characterization and in vitro selection for iron efficiency in Pyrus and Cydonia. In Vitro Cell. Dev. Biol. 1992, 28, 25–29. [Google Scholar] [CrossRef]
- Lombard, P.B.; Westwood, M.N. Pear rootstocks. In Rootstocks for Fruit Crops; Rom, R.C., Carlson, R.F., Eds.; John Wiley & Sons, Inc.: New York, NY, USA, 1987; pp. 145–183. [Google Scholar]
- Claveria, E.; Asin, L.; Iglesias, I.; Vilardell, P.; Bonany, J.; Simard, M.H.; Dolcet-Sanjuan, R. In Vitro Screening for Tolerance to Iron Chlorosis as a Reliable Selection Tool in a Pear Rootstock Breeding Program. In XXVIII International Horticultural Congress on Science and Horticulture for People; Leitao, J.M., Ed.; ISHS: Leuven, Belgium, 2012; pp. 199–205. [Google Scholar]
- Simard, M.H.; Michelesi, J.C. ‘Pyriam’: A new pear rootstock. Acta Hortic. 2002, 596, 351–355. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Chaney, R.L.; Coulombe, B.A.; Bell, P.F.; Angle, J.S. Detailed method to screen dicot cultivars for resistance to Fe-chlorosis using FeDTPA and bicarbonate in nutrient solutions. J. Plant Nutr. 1992, 15, 2063–2083. [Google Scholar] [CrossRef]
- Solecka, D.; Boudet, A.M.; Kacperska, A. Phenylpropanoid and anthocyanin changes in low temperature treated winter oilseed rape leaves. Plant Physiol. Biochem. 1999, 37, 491–496. [Google Scholar] [CrossRef]
- Kidd, P.S.; Llugany, M.; Poschenrieder, C.; Gunsé, B.; Barceló, J. The role of root exudates in aluminium resistance and silicon-induced amelioration of aluminium toxicity in tree varieties of maize (Zea mays L.). J. Exp. Bot. 2001, 52, 1339–1352. [Google Scholar] [PubMed]
- Tolrà, R.; Barceló, J.; Poschenrieder, C. Constitutive and Aluminium-induced Patterns of Phenolic Compounds in Two Maize Varieties Differing in Aluminium Tolerance. J. Inorg. Biochem. 2009, 103, 1486–1490. [Google Scholar] [CrossRef] [PubMed]
- Singlenton, V.J.; Orthofer, R.O.; Lamuela-Raventos, M.R. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Tolrà, R.; Poschenrieder, C.; Barceló, J. Zinc hyperaccumulation in Thlaspi caerulescens. II. Influence on organic acids. J. Plant Nutr. 1996, 19, 1541–1550. [Google Scholar] [CrossRef]
- Tolrà, R.P.; Poschenrieder, C.; Lupi, B.; Barceló, J. Aluminum-induced changes in the profiles of both organic acids and phenolic substances underlie Al tolerance in Rumex acetosa. Environ. Exp. Bot. 2005, 54, 231–238. [Google Scholar] [CrossRef]
- Petersen, R.G. Design and Analysis of Experiments; Marcel Dekker Inc.: New York, NY, USA, 1985. [Google Scholar] [CrossRef]
- Sun, X.-P.; Wang, S.Y.; Tong, Y.-A.; Korcak, R.F.; Faust, M. Metabolic changes in iron-deficient apple seedlings. J. Plant Nutr. 1987, 10, 1021–1030. [Google Scholar] [CrossRef]
- Rombolà, D.; Brüggemann, W.; López-Millán, A.F.; Tagliavini, M.; Abadía, J.; Marangoni, B.; Moog, P.R. Biochemical responses to iron deficiency in kiwifruit (Actinidia deliciosa). Tree Physiol. 2002, 22, 869–875. [Google Scholar] [CrossRef] [Green Version]
- Covarrubias, J.I.; Rombolà, A.D. Organic acids metabolism in roots of grapevine rootstocks under severe iron deficiency. Plant Soil 2015, 394, 165–175. [Google Scholar] [CrossRef]
- Alhendawi, R.A.; Römheld, V.; Kirkby, E.A.; Marschner, H. Influence of increasing bicarbonate concentrations on plant growth, organic acid accumulation in roots and iron uptake by barley, sorghum, and maize. J. Plant Nutr. 1997, 20, 1731–1753. [Google Scholar] [CrossRef]
- Bienfait, H.F. Is there a metabolic link between H+ excretion and ferric reduction by roots of Fe-deficient plants?—A viewpoint. J. Plant Nutr. 1996, 19, 1211–1222. [Google Scholar] [CrossRef]
- Landsberg, E.C. Organic acid synthesis and release of hydrogen ions in response to Fe deficiency stress of mono- and dicotyledonous plant species. J. Plant Nutr. 1981, 3, 579–591. [Google Scholar] [CrossRef]
- Martinez-Cuenca, M.; Primo-Capella, A.; Quiñones, A.; Bermejo, A.; Forner-Giner, M.A. Rootstock influence on iron uptake responses in Citrus leaves and their regulation under the Fe paradox effect. PeerJ 2017, 5, e3553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abadía, J.; López-Millán, A.F.; Rombolà, A.; Abadía, A. Organic acids and Fe deficiency: A review. Plant Soil 2002, 241, 75–86. [Google Scholar] [CrossRef]
- Morales, F.; Grasa, R.; Abadía, A.; Abadía, J. Iron chlorosis paradox in fruit trees. J. Plant Nutr. 1998, 21, 815–825. [Google Scholar] [CrossRef]
- Ortega-Maldonado, J.M.; Ojeda-Barrios, D.L.; Martínez-Téllez, J.J.; Hernández-Rodríguez, A.; de Jesús Ruíz-Anchondo, T.; Escudero-Almanza, D.J. Aspectos relevantes sobre la bioquímica y la fisiología del fierro en plantas. Tecnociencia Chihuah. 2014, 8, 30–38. [Google Scholar]

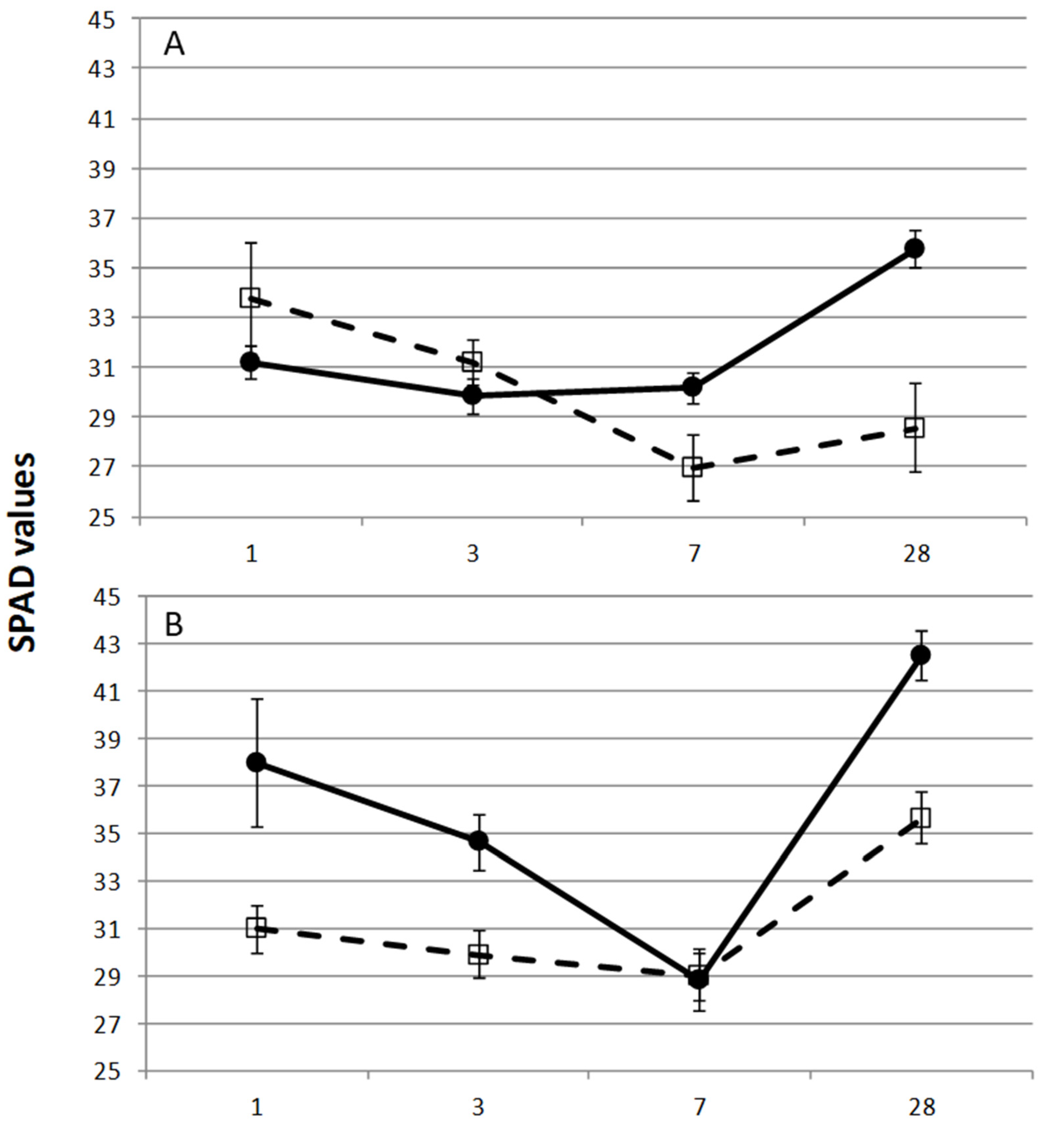
 ) have the same behavior and they are significantly different (p = 0.006) from Pcw (
) have the same behavior and they are significantly different (p = 0.006) from Pcw (  ). At 20 mM NaHCO3 (B), the Pap, Pcw, and Pe ( ) have the same behavior and they are significantly different (p = 0.034) from Pcc ( ). Mean values come from the measurements of the three upper first leaves from each plantlet, with four replicates per genotype. Bars represent the standard error.
) have the same behavior and they are significantly different (p = 0.006) from Pcw ( ). At 20 mM NaHCO3 (B), the Pap, Pcw, and Pe ( ) have the same behavior and they are significantly different (p = 0.034) from Pcc ( ). Mean values come from the measurements of the three upper first leaves from each plantlet, with four replicates per genotype. Bars represent the standard error.
). At 20 mM NaHCO3 (B), the Pap, Pcw, and Pe ( ) have the same behavior and they are significantly different (p = 0.034) from Pcc ( ). Mean values come from the measurements of the three upper first leaves from each plantlet, with four replicates per genotype. Bars represent the standard error.
) have the same behavior and they are significantly different (p = 0.006) from Pcw ( ). At 20 mM NaHCO3 (B), the Pap, Pcw, and Pe ( ) have the same behavior and they are significantly different (p = 0.034) from Pcc ( ). Mean values come from the measurements of the three upper first leaves from each plantlet, with four replicates per genotype. Bars represent the standard error.
 ) have the same behavior and they are significantly different (p = 0.027) from Pap ( ). At 20 mM NaHCO3 (B), Pcc ( ) is significantly different (p < 0.001) from Pa, Pap, Pcw, and Pe ( ), which have the same performance. Mean values come from the measures taken on the culture medium of four replicates, one plantlet each per genotype. Bars represent the standard error.
) have the same behavior and they are significantly different (p = 0.027) from Pap ( ). At 20 mM NaHCO3 (B), Pcc ( ) is significantly different (p < 0.001) from Pa, Pap, Pcw, and Pe ( ), which have the same performance. Mean values come from the measures taken on the culture medium of four replicates, one plantlet each per genotype. Bars represent the standard error.
) have the same behavior and they are significantly different (p = 0.027) from Pap ( ). At 20 mM NaHCO3 (B), Pcc ( ) is significantly different (p < 0.001) from Pa, Pap, Pcw, and Pe ( ), which have the same performance. Mean values come from the measures taken on the culture medium of four replicates, one plantlet each per genotype. Bars represent the standard error.
) have the same behavior and they are significantly different (p = 0.027) from Pap ( ). At 20 mM NaHCO3 (B), Pcc ( ) is significantly different (p < 0.001) from Pa, Pap, Pcw, and Pe ( ), which have the same performance. Mean values come from the measures taken on the culture medium of four replicates, one plantlet each per genotype. Bars represent the standard error.
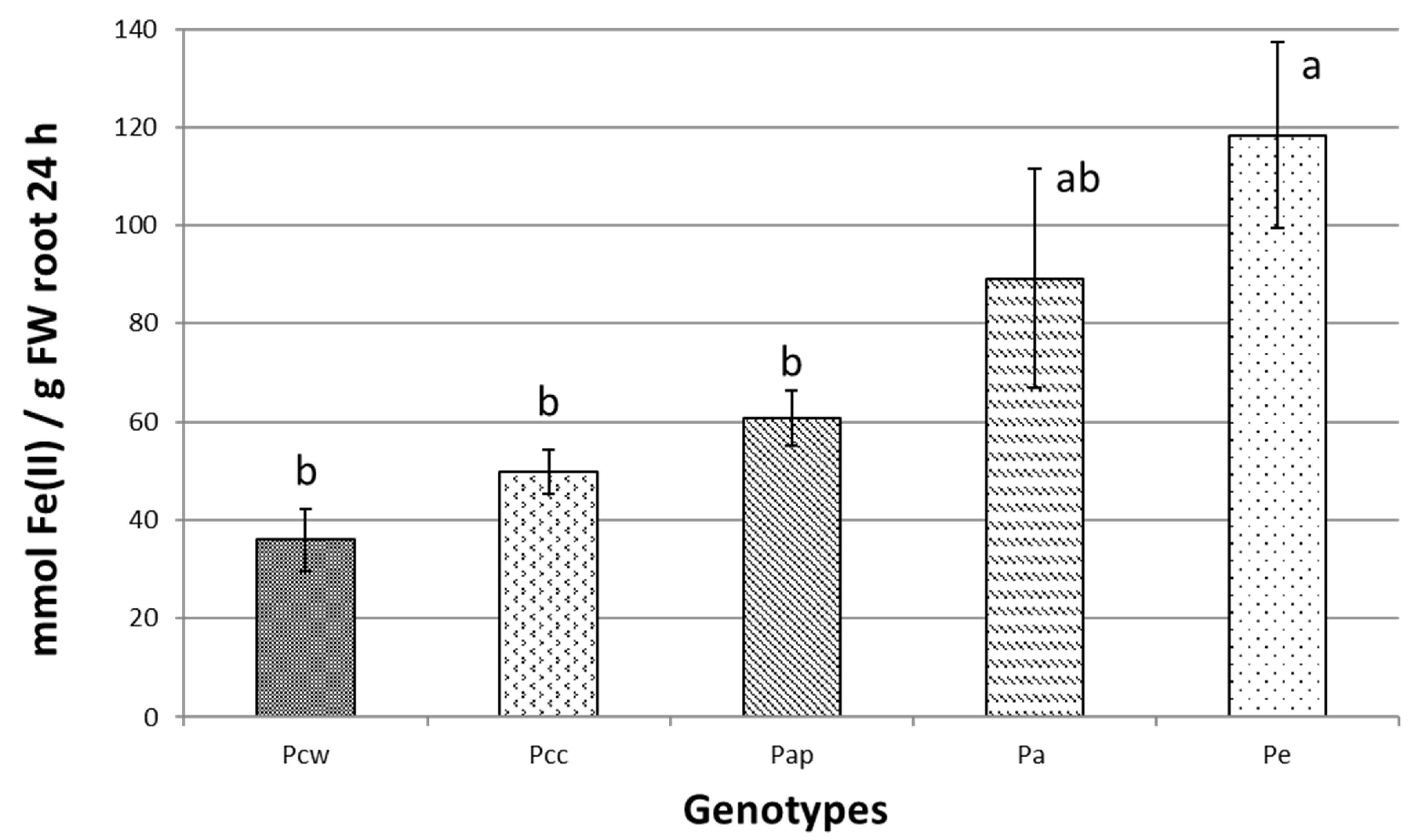
 ) is significantly different (p < 0.001) from Pa, Pap, Pcc, and Pcw ( ), which have the same reduction response. At 20 mM NaHCO3 (B), all genotypes ( ) have the same behavior (p = 0.229). Mean values come from the measures taken on the roots of four replicates, one plantlet each per genotype. Bars represent the standard error.
) is significantly different (p < 0.001) from Pa, Pap, Pcc, and Pcw ( ), which have the same reduction response. At 20 mM NaHCO3 (B), all genotypes ( ) have the same behavior (p = 0.229). Mean values come from the measures taken on the roots of four replicates, one plantlet each per genotype. Bars represent the standard error.
) is significantly different (p < 0.001) from Pa, Pap, Pcc, and Pcw ( ), which have the same reduction response. At 20 mM NaHCO3 (B), all genotypes ( ) have the same behavior (p = 0.229). Mean values come from the measures taken on the roots of four replicates, one plantlet each per genotype. Bars represent the standard error.
) is significantly different (p < 0.001) from Pa, Pap, Pcc, and Pcw ( ), which have the same reduction response. At 20 mM NaHCO3 (B), all genotypes ( ) have the same behavior (p = 0.229). Mean values come from the measures taken on the roots of four replicates, one plantlet each per genotype. Bars represent the standard error.
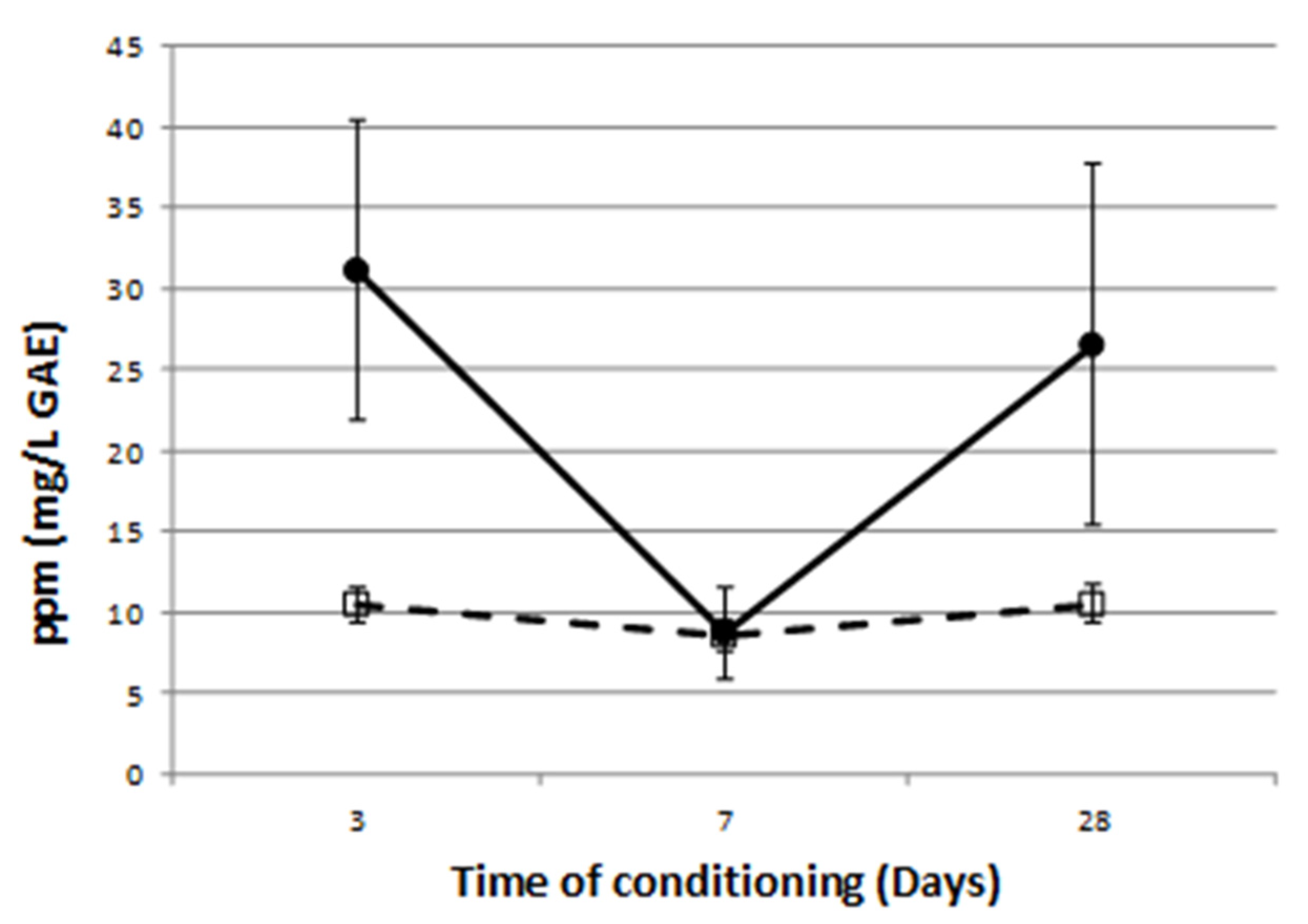
 ) is significantly different (p < 0.001) from Pa, Pap, Pcc, and Pe ( ), which have the same phenol exudation response. Since no significant interaction was observed between NaHCO3 (p = 0.713), mean values come from the measures taken on the roots of eight replicates, one plantlet each per genotype. Bars represent the standard error.
) is significantly different (p < 0.001) from Pa, Pap, Pcc, and Pe ( ), which have the same phenol exudation response. Since no significant interaction was observed between NaHCO3 (p = 0.713), mean values come from the measures taken on the roots of eight replicates, one plantlet each per genotype. Bars represent the standard error.
) is significantly different (p < 0.001) from Pa, Pap, Pcc, and Pe ( ), which have the same phenol exudation response. Since no significant interaction was observed between NaHCO3 (p = 0.713), mean values come from the measures taken on the roots of eight replicates, one plantlet each per genotype. Bars represent the standard error.
) is significantly different (p < 0.001) from Pa, Pap, Pcc, and Pe ( ), which have the same phenol exudation response. Since no significant interaction was observed between NaHCO3 (p = 0.713), mean values come from the measures taken on the roots of eight replicates, one plantlet each per genotype. Bars represent the standard error.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mora-Córdova, C.P.; Tolrà, R.; Padilla, R.; Poschenrieder, C.; Simard, M.-H.; Asín, L.; Vilardell, P.; Bonany, J.; Claveria, E.; Dolcet-Sanjuan, R. Rhizosphere Acidification as the Main Trait Characterizing the Differential In Vitro Tolerance to Iron Chlorosis in Interspecific Pyrus Hybrids. Horticulturae 2022, 8, 551. https://doi.org/10.3390/horticulturae8060551
Mora-Córdova CP, Tolrà R, Padilla R, Poschenrieder C, Simard M-H, Asín L, Vilardell P, Bonany J, Claveria E, Dolcet-Sanjuan R. Rhizosphere Acidification as the Main Trait Characterizing the Differential In Vitro Tolerance to Iron Chlorosis in Interspecific Pyrus Hybrids. Horticulturae. 2022; 8(6):551. https://doi.org/10.3390/horticulturae8060551
Chicago/Turabian StyleMora-Córdova, Claudia Paola, Roser Tolrà, Rosa Padilla, Charlotte Poschenrieder, Marie-Helene Simard, Luis Asín, Pere Vilardell, Joan Bonany, Elisabet Claveria, and Ramon Dolcet-Sanjuan. 2022. "Rhizosphere Acidification as the Main Trait Characterizing the Differential In Vitro Tolerance to Iron Chlorosis in Interspecific Pyrus Hybrids" Horticulturae 8, no. 6: 551. https://doi.org/10.3390/horticulturae8060551
APA StyleMora-Córdova, C. P., Tolrà, R., Padilla, R., Poschenrieder, C., Simard, M. -H., Asín, L., Vilardell, P., Bonany, J., Claveria, E., & Dolcet-Sanjuan, R. (2022). Rhizosphere Acidification as the Main Trait Characterizing the Differential In Vitro Tolerance to Iron Chlorosis in Interspecific Pyrus Hybrids. Horticulturae, 8(6), 551. https://doi.org/10.3390/horticulturae8060551