
Functional Characterization of Tea Plant (Camellia sinensis L.) CsCBF2 Gene Involved in Multiple Abiotic Stress Response in Tobacco (Nicotiana tabacum L.)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
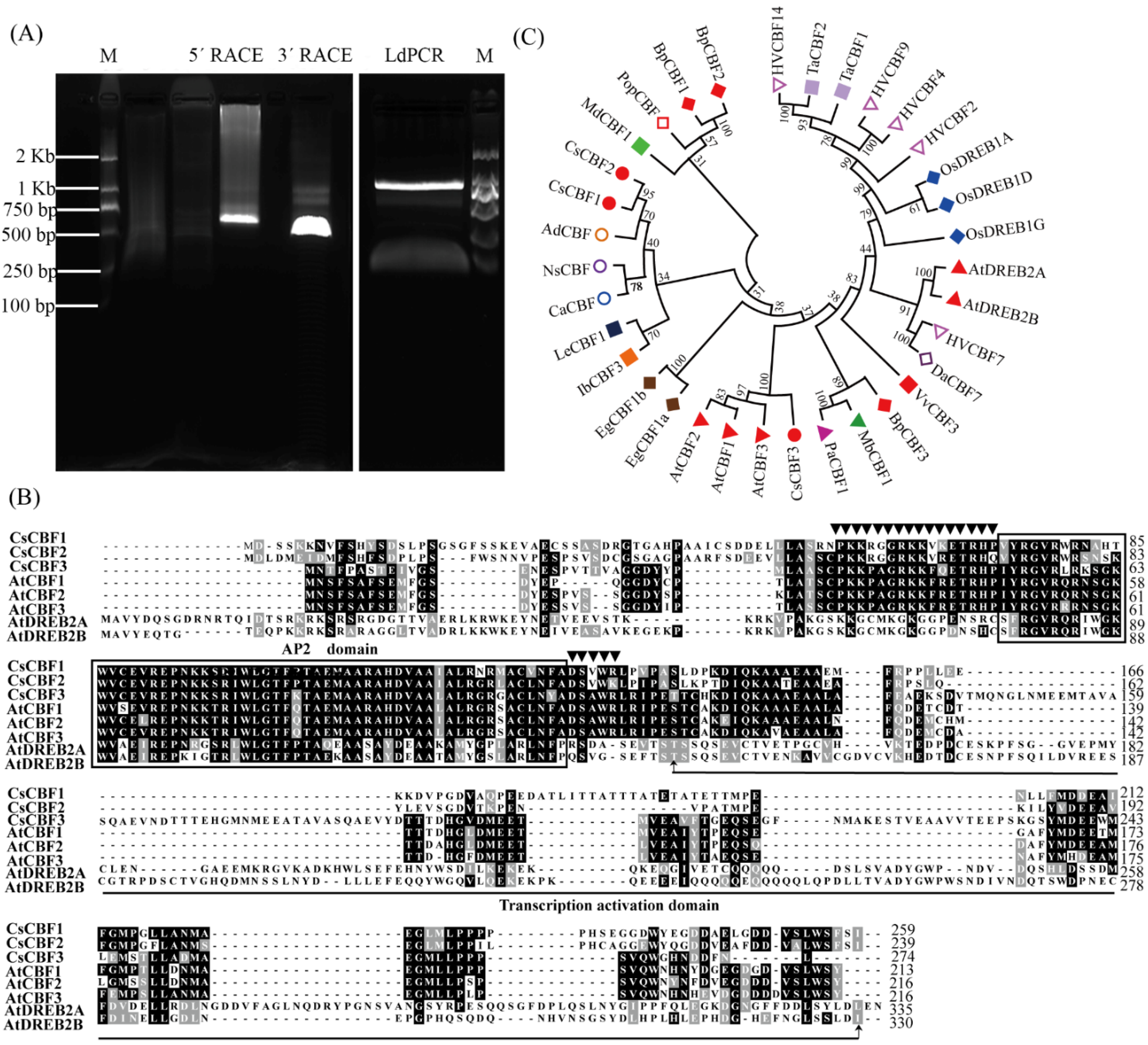
2.2. RACE Cloning of CsCBF2 Gene
2.3. Sequence Analysis and Phylogenetic Tree Building
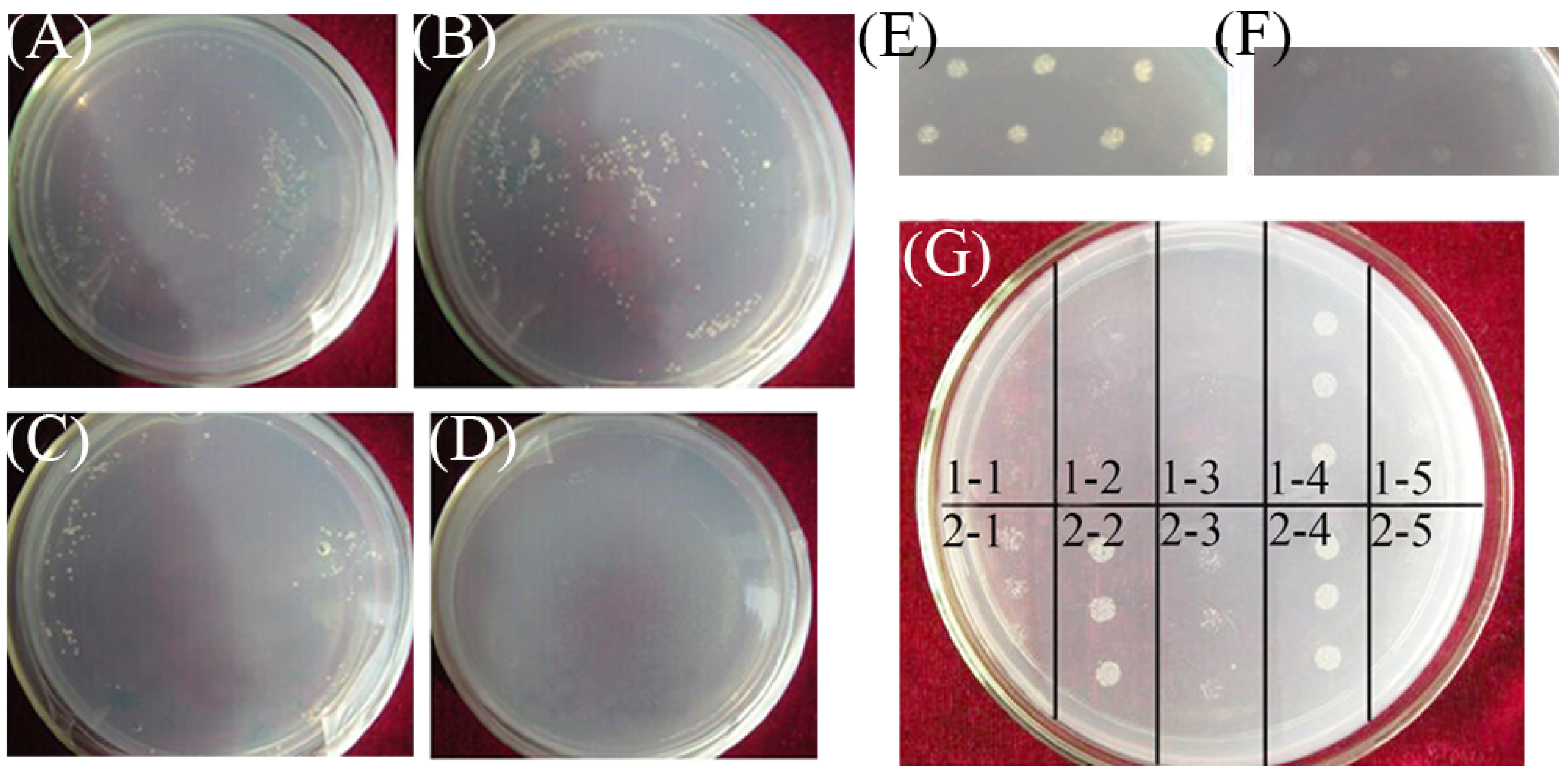
2.4. Yeast One-Hybrid and Transcription Activation Assays
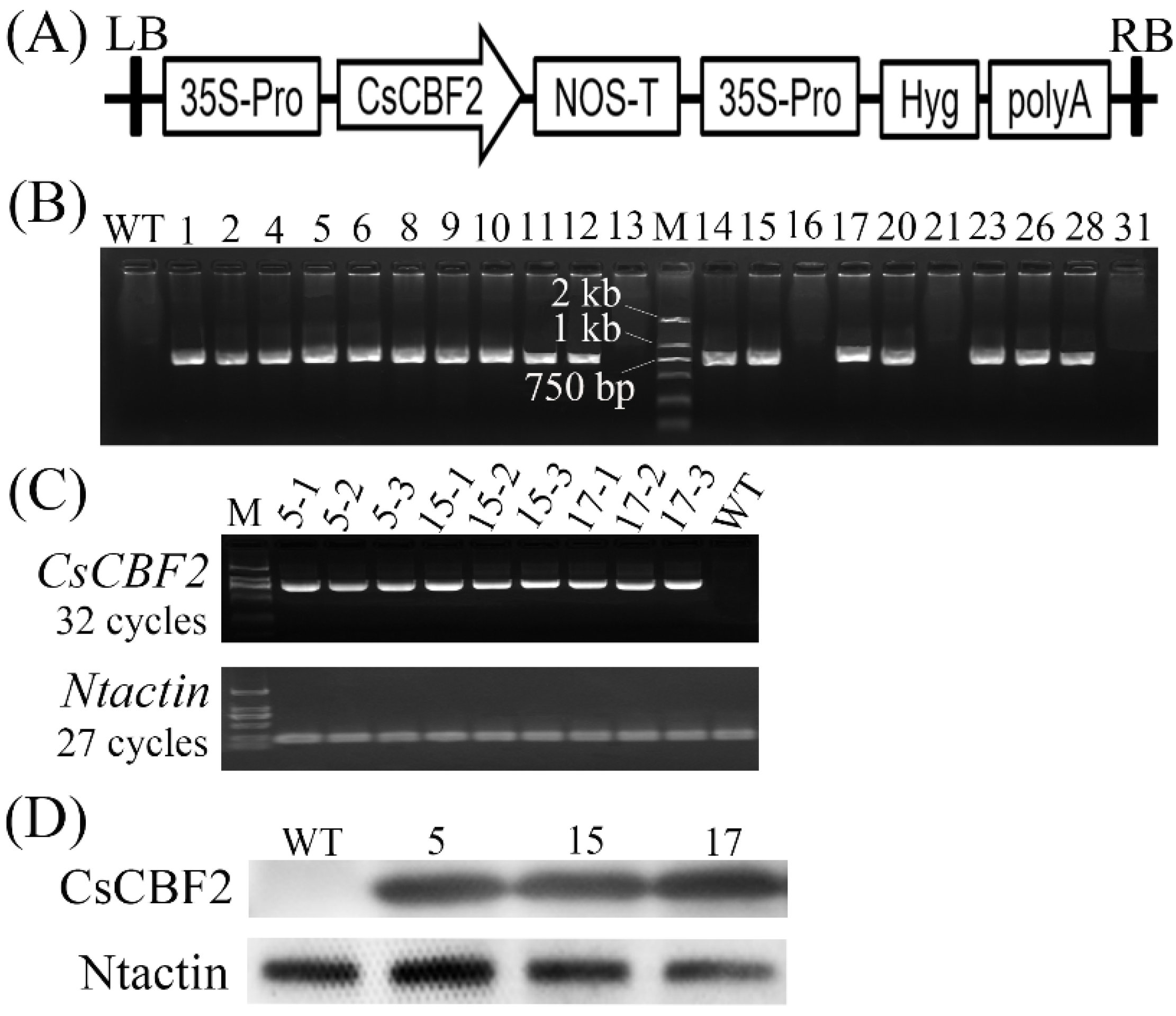
2.5. Agrobacterium-Mediated Transformation of Tobacco Plants
2.6. Molecular Characterization of CsCBF2-Overexpressing Transgenic Tobacco Plants
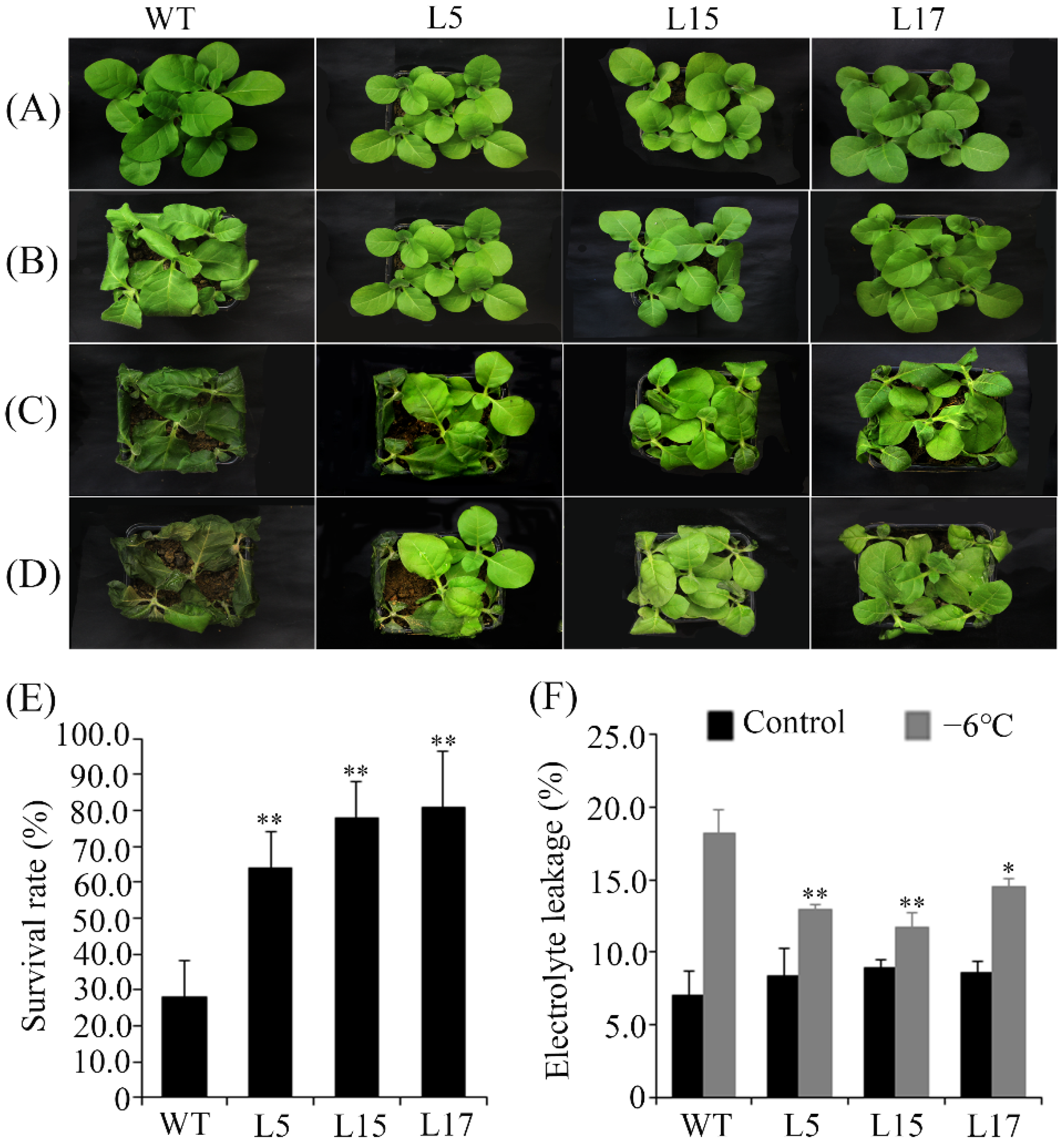
2.7. Low Temperature Tolerance Assays of CsCBF2-Overexpressing Transgenic Plants
2.8. Salt and Drought Tolerance Assays of CsCBF2-Overexpressing Transgenic Plants
2.9. Cell Membrane Stability Assay
2.10. Soluble Sugar and Proline Contents Analysis
3. Results
3.1. Isolation and Characterization of CsCBF2 Gene
3.2. CRT/DRE-Binding and Transactivation Assay of CsCBF2
3.3. Generation and Molecular Identification of CsCBF2 Transgenic Plants
3.4. Overexpression of CsCBF2 Gene Leads to Phenotypic Alterations of Tobacco Plant
3.5. Overexpression of CsCBF2 Confers Cold Tolerance in Tobacco Plant
3.6. Freezing Tolerance of CsCBF2-Overexpressing Transgenic Tobacco Plants
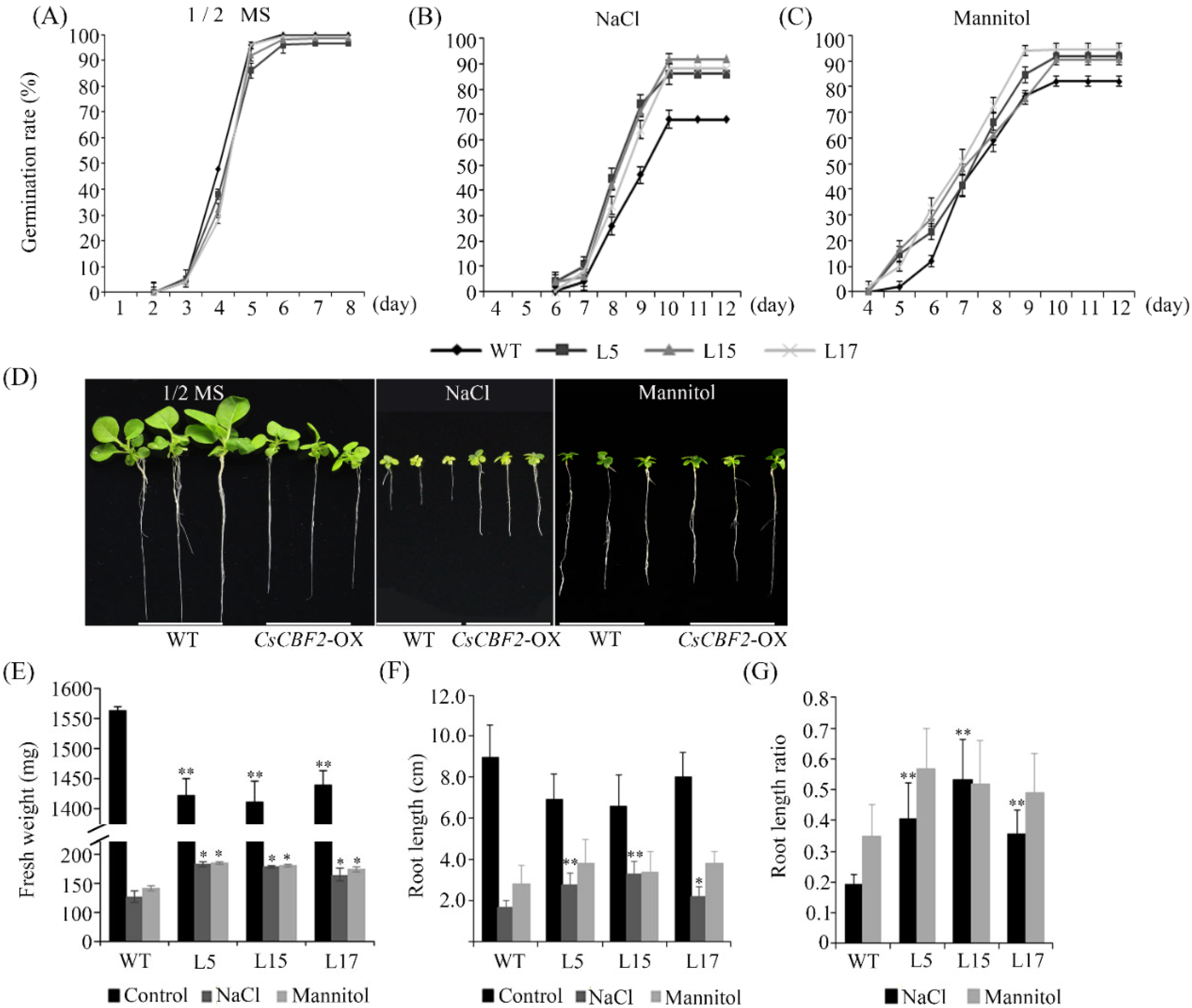
3.7. CsCBF2 Transgenic Plants Exhibit Increased Tolerance to Drought and Salt Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wei, T.; Deng, K.; Zhang, Q.; Gao, Y.; Liu, Y.; Yang, M.; Zhang, L.; Zheng, X.; Wang, C.; Liu, Z.; et al. Modulating AtDREB1C expression improves drought tolerance in Salvia miltiorrhiza. Front. Plant Sci. 2017, 8, 52. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Han, Y.; Pan, J.; Yuan, H.; Li, X.; Qin, M.; Peng, B.; Zhu, Y. Research progress in plant cold resistance mechanism. J. Xinyang Norm. Univ. (Nat. Sci. Ed.) 2019, 32, 511–516. [Google Scholar]
- Hu, Z.; Ban, Q.; Hao, J.; Zhu, X.; Cheng, Y.; Mao, J.; Lin, M.; Xia, E.; Li, Y. Genome-Wide characterization of the C-repeat binding factor (CBF) genefamily involved in the response to abiotic stresses in tea plant (Camellia sinensis). Front. Plant Sci. 2020, 11, 921. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Lv, J.; Ma, Z.; Dong, W. The mechanism of alfalfa (Medicago sativa L.) response to abiotic stress. Plant Growth Regul. 2019, 89, 239–249. [Google Scholar] [CrossRef]
- Zhang, J.; Qian, J.-Y.; Bian, Y.-H.; Liu, X.; Wang, C.-L. Transcriptome and Metabolite Conjoint Analysis Reveals the Seed Dormancy Release Process in Callery Pear. Int. J. Mol. Sci. 2022, 23, 2186. [Google Scholar] [CrossRef]
- Nassuth, A.; Siddiqua, M.; Xiao, H.; Moody, M.A.; Carlow, C.E. Newly developed quantitative transactivation system shows difference in activation by Vitis CBF transcription factors on DRE/CRT elements. Plant Methods 2014, 10, 32. [Google Scholar] [CrossRef]
- Erpen, L.; Devi, H.S.; Grosser, J.W.; Dutt, M. Potential use of the DREB/ERF, MYB, NAC and WRKY transcription factors to improve abiotic and biotic stress in transgenic plants. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 132, 1–25. [Google Scholar] [CrossRef]
- Moon, S.-J.; Min, M.K.; Kim, J.-A.; Kim, D.Y.; Yoon, I.S.; Kwon, T.R.; Byun, M.O.; Kim, B.-G. Ectopic expression of OsDREB1G, a member of the OsDREB1 Subfamily, confers cold stress tolerance in rice. Front. Plant Sci. 2019, 10, 297. [Google Scholar] [CrossRef]
- Stockinger, E.J.; Gilmour, S.J.; Thomashow, M.F. Arabidopsis thaliana CBF1 encodes an AP2 domain-containing transcriptional activator that binds to the C-repeat/DRE, a cis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit. Proc. Natl. Acad. Sci. USA 1997, 94, 1035–1040. [Google Scholar] [CrossRef]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef]
- Gutha, L.R.; Reddy, A.R. Rice DREB1B promoter shows distinct stress-specific responses, and the overexpression of cDNA in tobacco confers improved abiotic and biotic stress tolerance. Plant Mol. Biol. 2008, 68, 533–555. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Yu, B.; Song, Q.; Liu, Y.; Chen, T.H.; Li, G.; Yang, X. Ectopic expression of StCBF1 and ScCBF1 have different functions in response to freezing and drought stresses in Arabidopsis. Plant Sci. 2018, 270, 221–233. [Google Scholar] [CrossRef]
- Zhou, L.; Li, J.; He, Y.; Liu, Y.; Chen, H. Functional characterization of SmCBF genes involved in abiotic stress response in eggplant (Solanum melongena). Sci. Hortic. 2018, 233, 14–21. [Google Scholar] [CrossRef]
- Wu, J.; Folta, K.M.; Xie, Y.; Jiang, W.; Lu, J.; Zhang, Y. Overexpression of Muscadinia rotundifolia CBF2 gene enhances biotic and abiotic stress tolerance in Arabidopsis. Protoplasma 2017, 254, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Li, J.; Long, S.; Wei, S. A DREB1 gene from zoysiagrass enhances Arabidopsis tolerance to temperature stresses without growth inhibition. Plant Sci. 2019, 278, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The transcriptional regulatory network in the drought response and its crosstalk in abiotic stress responses including drought, cold, and heat. Front. Plant Sci. 2014, 5, 170. [Google Scholar] [CrossRef]
- Wang, P.; Chen, X.; Guo, Y.; Zheng, Y.; Yue, C.; Yang, J.; Ye, N. Identification of CBF transcription factors in tea plants and a survey of potential CBF target genes under low temperature. Int. J. Mol. Sci. 2019, 20, 5137. [Google Scholar] [CrossRef]
- Herath, V. Small family, big impact: In silico analysis of DREB2 transcription factor family in rice. Comput. Biol. Chem. 2016, 65, 128–139. [Google Scholar] [CrossRef]
- Hichri, I.; Muhovski, Y.; Clippe, A.; Žižková, E.; Dobrev, P.I.; Motyka, V.; Lutts, S. SlDREB2, a tomato dehydration-responsive element-binding 2 transcription factor, mediates salt stress tolerance in tomato and Arabidopsis. Plant Cell Environ. 2016, 39, 62–79. [Google Scholar] [CrossRef]
- Zhang, X.-X.; Tang, Y.-J.; Ma, Q.-B.; Yang, C.-Y.; Mu, Y.-H.; Suo, H.-C.; Luo, L.-H.; Nian, H. OsDREB2A, a rice transcription factor, significantly affects salt tolerance in transgenic soybean. PLoS ONE 2013, 8, e83011. [Google Scholar] [CrossRef]
- Li, X.; Zhang, D.; Li, H.; Wang, Y.; Zhang, Y.; Wood, A.J. EsDREB2B, a novel truncated DREB2-type transcription factor in the desert legume Eremosparton songoricum, enhances tolerance to multiple abiotic stresses in yeast and transgenic tobacco. BMC Plant Biol. 2014, 14, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Huang, L.; Yan, H.; Zhang, X.; Xu, B.; Ma, X. Cloning and characterization of an ABA-independent DREB transcription factor gene, HcDREB2, in Hemarthria compressa. Hereditas 2016, 153, 3. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, M.; Abdullah, S.N.A.; Aziz, M.A.; Namasivayam, P. A novel CBF that regulates abiotic stress response and the ripening process in oil palm (Elaeis guineensis) fruits. Tree Genet. Genomes 2015, 11, 56. [Google Scholar] [CrossRef]
- Gao, W.; He, M.; Liu, J.; Ma, X.; Zhang, Y.; Dai, S.; Zhou, Y. Overexpression of Chrysanthemum lavandulifolium ClCBF1 in Chrysanthemum morifolium ‘White Snow’ improves the level of salinity and drought tolerance. Plant Physiol. Biochem. 2018, 124, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Ritonga, F.N.; Ngatia, J.N.; Wang, Y.; Khoso, M.A.; Farooq, U.; Chen, S. AP2/ERF, an important cold stress-related transcription factor family in plants: A review. Physiol. Mol. Biol. Plants 2021, 27, 1953–1968. [Google Scholar] [CrossRef] [PubMed]
- Byun, M.Y.; Lee, J.; Cui, L.H.; Kang, Y.; Oh, T.K.; Park, H.; Lee, H.; Kim, W.T. Constitutive expression of DaCBF7, an antarctic vascular plant Deschampsia antarctica CBF homolog, resulted in improved cold tolerance in transgenic rice plants. Plant Sci. 2015, 236, 61–74 . [Google Scholar] [CrossRef]
- Siddiqua, M.; Nassuth, A. Vitis CBF1 and Vitis CBF4 differ in their effect on Arabidopsis abiotic stress tolerance, development and gene expression. Plant Cell Environ. 2011, 34, 1345–1359. [Google Scholar] [CrossRef]
- Xiong, Y.; Fei, S.-Z. Functional and phylogenetic analysis of a DREB/CBF-like gene in perennial ryegrass (Lolium perenne L.). Planta 2006, 224, 878–888. [Google Scholar] [CrossRef]
- Jin, R.; Kim, B.H.; Ji, C.Y.; Kim, H.S.; Li, H.M.; Ma, D.F.; Kwak, S.-S. Overexpressing IbCBF3 increases low temperature and drought stress tolerance in transgenic sweetpotato. Plant Physiol. Biochem. 2017, 118, 45–54. [Google Scholar] [CrossRef]
- Jia, Y.; Ding, Y.; Shi, Y.; Zhang, X.; Gong, Z.; Yang, S. The cbfs triple mutants reveal the essential functions of CBFs in cold acclimation and allow the definition of CBF regulons in Arabidopsis. New Phytol. 2016, 212, 345–353. [Google Scholar] [CrossRef]
- Yin, S.; Han, Y.; Huang, L.; Hong, Y.; Zhang, G. Overexpression of HvCBF7 and HvCBF9 changes salt and drought tolerance in Arabidopsis. Plant Growth Regul. 2018, 85, 281–292. [Google Scholar] [CrossRef]
- Carlow, C.E.; Faultless, J.T.; Lee, C.; Siddiqua, M.; Edge, A.; Nassuth, A. Nuclear localization and transactivation by Vitis CBF transcription factors are regulated by combinations of conserved amino acid domains. Plant Physiol. Biochem. 2017, 118, 306–319. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhuang, J.; Zou, Z.; Li, Q.; Xin, H. Overexpression of a Camellia sinensis DREB transcription factor gene (CsDREB) increases salt and drought tolerance in transgenic Arabidopsis thaliana. J. Plant Biol. 2017, 60, 452–461. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhao, M.; Xing, F.; Mao, G.; Wang, Y.; Dai, Y.; Niu, M.; Yuan, H. Identification and expression analysis of CAMTA genes in tea plant reveal their complex regulatory role in stress responses. Front. Plant Sci. 2022, 13, 910768. [Google Scholar] [CrossRef]
- Yin, Y.; Ma, Q.-P.; Zhu, Z.-X.; Cui, Q.-Y.; Chen, C.-S.; Chen, X.; Fang, W.-P.; Li, X.-H. Functional analysis of CsCBF3 transcription factor in tea plant (Camellia sinensis) under cold stress. Plant Growth Regul. 2016, 80, 335–343. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.; Zhao, Q.; Jing, S.; Chen, S.; Yuan, H. Identification of Genes Induced in Response to Low-Temperature Treatment in Tea Leaves. Plant Mol. Biol. Rep. 2009, 27, 257–265. [Google Scholar] [CrossRef]
- Yuan, H.; Zhu, X.; Zeng, W.; Yang, H.; Sun, N.; Xie, S.; Cheng, L. Isolation and transcription activation analysis of the CsCBF1 gene from Camellia sinensis. Acta Bot. Boreal. Occident. Sin. 2013, 33, 1717–1723. [Google Scholar]
- Zhao, P.; Zhou, X.-M.; Zhang, L.-Y.; Wang, W.; Ma, L.; Yang, L.-B.; Peng, X.-B.; Bozhkov, P.; Sun, M.-X. A Bipartite molecular module controls cell death activation in the basal cell lineage of plant embryos. PLoS Biol. 2013, 11, e1001655. [Google Scholar] [CrossRef]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Zhou, Q.; Yu, Q.; Wang, Z.; Pan, Y.; Lv, W.; Zhu, L.; Chen, R.; He, G. Knockdown of GDCH gene reveals reactive oxygen species-induced leaf senescence in rice. Plant Cell Environ. 2013, 36, 1476–1489. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, X.; Qin, Y.; Feng, B.; Wu, Y.; He, Y.; Wang, A.; Zhu, J. The chilling tolerance divergence 1 protein confers cold stress tolerance in processing tomato. Plant Physiol. Biochem. 2020, 151, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Yu, G.; Chen, H.; Wang, B.; Jiang, L.; Han, X.; Lin, G.; Cheng, X.-G. SlDREB2 gene specifically recognizing to the universal DRE elements is a transcriptional activator improving drought tolerance in tomato. Sci. Hortic. 2022, 295, 110887. [Google Scholar] [CrossRef]
- Xia, E.; Tong, W.; Hou, Y.; An, Y.; Chen, L.; Wu, Q.; Liu, Y.; Yu, J.; Li, F.; Li, R.; et al. The reference genome of tea plant and resequencing of 81 diverse accessions provide insights into its genome evolution and adaptation. Mol. Plant 2020, 13, 1013–1026. [Google Scholar] [CrossRef] [PubMed]
- Knight, A.; Piskacek, M. Cryptic inhibitory regions nearby activation domains. Biochimie 2022, 200, 19–26. [Google Scholar] [CrossRef]
- Sakuma, Y.; Maruyama, K.; Osakabe, Y.; Qin, F.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of an Arabidopsis transcription factor, DREB2A, involved in drought-responsive gene expression. Plant Cell 2006, 18, 1292–1309. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, X.; Li, M.; Yang, H.; Fu, D.; Lv, J.; Ding, Y.; Gong, Z.; Shi, Y.; Yang, S. The direct targets of CBFs: In cold stress response and beyond. J. Integr. Plant Biol. 2021, 63, 1874–1887. [Google Scholar] [CrossRef]
- Lu, J.; Wang, L.; Zhang, Q.; Ma, C.; Su, X.; Cheng, H.; Guo, H. AmCBF1 transcription factor regulates plant architecture by repressing GhPP2C1 or GhPP2C2 in Gossypium hirsutum. Front. Plant Sci. 2022, 13, 914206. [Google Scholar] [CrossRef]
- Oh, S.J.; Song, S.I.; Kim, Y.S.; Jang, H.J.; Kim, S.Y.; Kim, M.; Kim, Y.K.; Nahm, B.H.; Kim, J.K. Arabidopsis CBF3/DREB1A and ABF3 in transgenic rice increased tolerance to abiotic stress without stunting growth. Plant Physiol. 2005, 138, 341–351. [Google Scholar] [CrossRef]
- Shimazaki, T.; Endo, T.; Kasuga, M.; Yamaguchi-Shinozaki, K.; Watanabe, K.N.; Kikuchi, A. Evaluation of the yield of abiotic-stress-tolerant AtDREB1A transgenic potato under saline conditions in advance of field trials. Breed. Sci. 2016, 66, 703–710. [Google Scholar] [CrossRef]
- Navarrete-Campos, D.; Le Feuvre, R.; Balocchi, C.; Valenzuela, S. Overexpression of three novel CBF transcription factors from Eucalyptus globulus improves cold tolerance on transgenic Arabidopsis thaliana. Trees 2017, 31, 1041–1055. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Meng, Z.; Wei, Y.; Du, X.; Liang, C.; Zhang, R. Elevation of GhDREB1B transcription by a copy number variant significantly improves chilling tolerance in cotton. Planta 2021, 254, 42. [Google Scholar] [CrossRef] [PubMed]
- do Rego, T.F.C.; Santos, M.P.; Cabral, G.B.; de Moura Cipriano, T.; de Sousa, N.L.; de Souza Neto, O.A.; Aragão, F.J.L. Expression of a DREB 5-A subgroup transcription factor gene from Ricinus communis (RcDREB1) enhanced growth, drought tolerance and pollen viability in tobacco. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 146, 493–504. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, Z.; Xie, S.; Si, T.; Li, Y.; Zhu, J.-K. Mutational evidence for the critical role of CBF transcription factors in cold acclimation in Arabidopsis. Plant Physiol. 2016, 171, 2744–2759. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Li, X.; Zhang, D.; Gao, B.; Yang, H.; Wang, Y.; Guan, K.; Wood, A.J. ScDREB8, a novel A-5 type of DREB gene in the desert moss Syntrichia caninervis, confers salt tolerance to Arabidopsis. Plant Physiol. Biochem. 2017, 120, 242–251. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Q.; Liu, D.; Wei, Y.; Ma, N.; Zhang, R.; Zhang, Z.; Jiang, C.; Yuan, H. Functional Characterization of Tea Plant (Camellia sinensis L.) CsCBF2 Gene Involved in Multiple Abiotic Stress Response in Tobacco (Nicotiana tabacum L.). Horticulturae 2022, 8, 853. https://doi.org/10.3390/horticulturae8090853
Zhou Q, Liu D, Wei Y, Ma N, Zhang R, Zhang Z, Jiang C, Yuan H. Functional Characterization of Tea Plant (Camellia sinensis L.) CsCBF2 Gene Involved in Multiple Abiotic Stress Response in Tobacco (Nicotiana tabacum L.). Horticulturae. 2022; 8(9):853. https://doi.org/10.3390/horticulturae8090853
Chicago/Turabian StyleZhou, Qiying, Dongxiao Liu, Yiwan Wei, Ning Ma, Ruijiao Zhang, Zengya Zhang, Changjun Jiang, and Hongyu Yuan. 2022. "Functional Characterization of Tea Plant (Camellia sinensis L.) CsCBF2 Gene Involved in Multiple Abiotic Stress Response in Tobacco (Nicotiana tabacum L.)" Horticulturae 8, no. 9: 853. https://doi.org/10.3390/horticulturae8090853
APA StyleZhou, Q., Liu, D., Wei, Y., Ma, N., Zhang, R., Zhang, Z., Jiang, C., & Yuan, H. (2022). Functional Characterization of Tea Plant (Camellia sinensis L.) CsCBF2 Gene Involved in Multiple Abiotic Stress Response in Tobacco (Nicotiana tabacum L.). Horticulturae, 8(9), 853. https://doi.org/10.3390/horticulturae8090853