
The Role of 1-methylcyclopropylene (1-MCP) and Salicylic Acid (SA) in Induced Resistance of Postharvest Fruits

Abstract
:1. Introduction
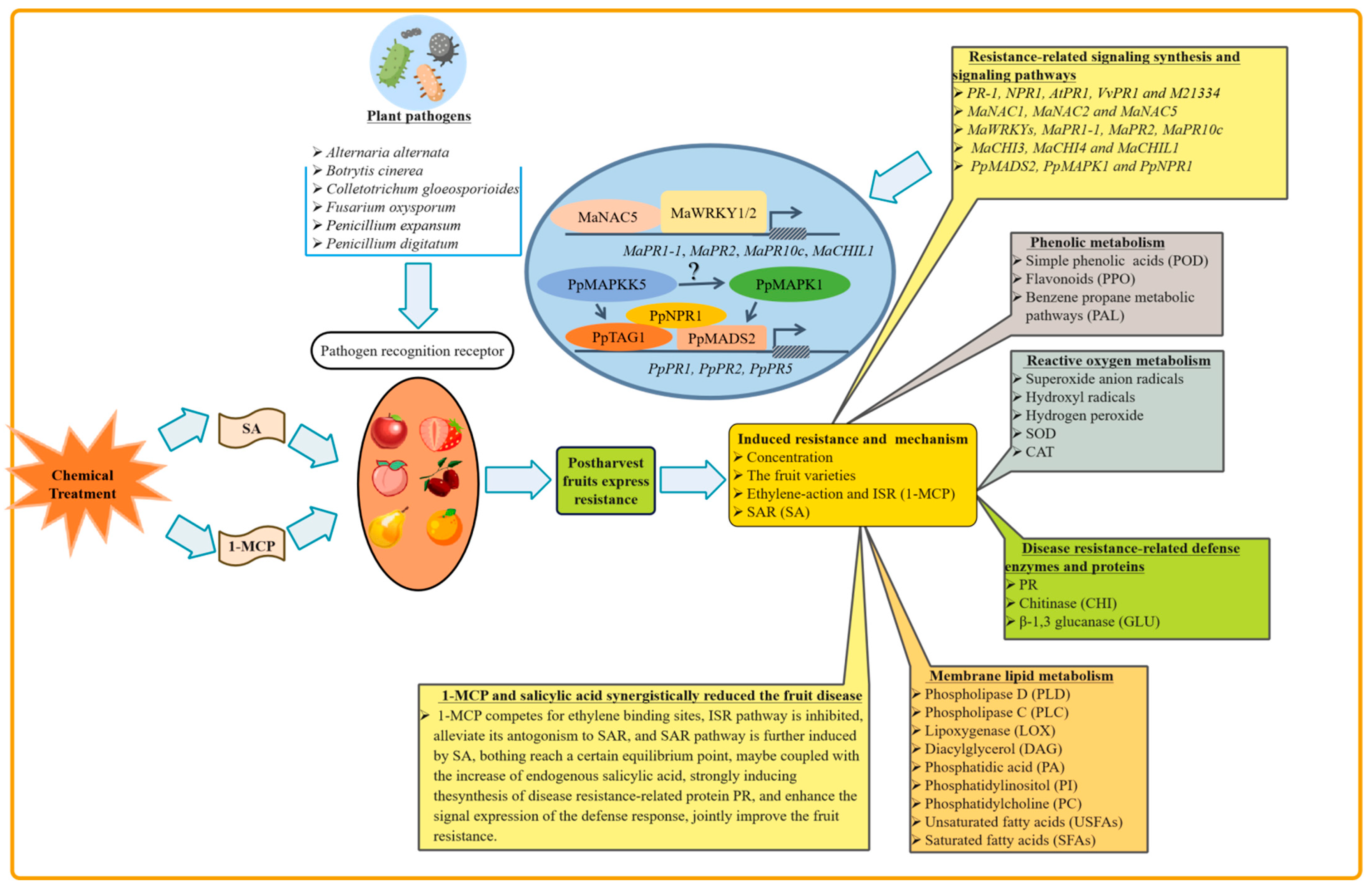
2. The Role of 1-MCP in Induced Resistance of Postharvest Fruits
2.1. Effect of 1-MCP on Induced Resistance and Its Involved Mechanism
2.2. Effect of 1-MCP on Reactive Oxygen Metabolism
2.3. Effect of 1-MCP on Phenolic Metabolism
2.4. Effect of 1-MCP on Disease Resistance-Related Defense Enzymes and Proteins
2.5. Effect of 1-MCP on Fruit Firmness
2.6. Effect of 1-MCP on Increasing Fruit Disease
3. The Role of SA in Induced Resistance of Postharvest Fruits
3.1. Effect of SA on Induced Resistance and Its Involved Mechanism
3.2. Effect of SA on Reactive Oxygen Metabolism
3.3. Effects of SA on Membrane Lipid Metabolism
3.4. Effect of SA on Phenolic Metabolism
3.5. Effect of SA on Disease Resistance-Related Signaling Synthesis and Signaling Pathways
3.6. Effect of SA on Disease Resistance-Related Defense Enzymes and Proteins
4. 1-MCP and SA Synergistically Reduced the Fruit Disease
5. Conclusions and Prospect
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Belay, Z.A.; Caleb, O.J. Role of integrated omics in unravelling fruit stress and defence responses during postharvest: A review. Food Chem. Mol. Sci. 2022, 5, 100118. [Google Scholar] [CrossRef] [PubMed]
- Chan, Z.; Tian, S. Interaction of antagonistic yeasts against postharvest pathogens of apple fruit and possible mode of action. Postharvest Biol. Technol. 2005, 36, 215–223. [Google Scholar] [CrossRef]
- Cai, X.; Xu, Z.; Li, X.; Wang, D.; Ren, X.; Kong, Q. Underlying mechanism of menthol on controlling postharvest citrus sour rot caused by Geotrichum citri-aurantii. Postharvest Biol. Technol. 2023, 196, 112160. [Google Scholar] [CrossRef]
- Cao, Z.; Zhou, D.; Ge, X.; Luo, Y.; Su, J. The role of essential oils in maintaining the postharvest quality and preservation of peach and other fruits. J. Food Biochem. 2022, 46, e14513. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Jia, X.; Li, J.; Li, X.; Jiang, J.; Li, H.; Zheng, Y.; Liu, Z.; Zhang, X.; Fan, J. Regulation effects of 1-MCP combined with flow microcirculation of sterilizing medium on peach shelf quality. Sci. Hortic. 2020, 260, 108867. [Google Scholar] [CrossRef]
- Testempasis, S.; Tanou, G.; Minas, I.; Samiotaki, M.; Molassiotis, A.; Karaoglanidis, G. Unraveling Interactions of the Necrotrophic Fungal Species Botrytis cinerea with 1-Methylcyclopropene or Ozone-Treated Apple Fruit Using Proteomic Analysis. Front. Recent Dev. Plant Sci. 2021, 12, 644255. [Google Scholar] [CrossRef]
- Palmer, I.A.; Shang, Z.; Fu, Z.Q. Salicylic acid-mediated plant defense: Recent developments, missing links, and future outlook. Front. Cell Dev. Biol. 2017, 12, 258–270. [Google Scholar] [CrossRef]
- Li, S.; Huan, C.; Liu, Y.; Zheng, X.; Bi, Y. Melatonin induces improved protection against Botrytis cinerea in cherry tomato fruit by activating salicylic acid signaling pathway. Sci. Hortic. 2022, 304, 111299. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, F.; Wang, J.; Yang, Q.; Wang, P.; Zhao, H.; Wang, J.; Wang, C.; Xu, X. Salicylic acid inhibits the postharvest decay of goji berry (Lycium barbarum L.) by modulating the antioxidant system and phenylpropanoid metabolites. Postharvest Biol. Technol. 2021, 178, 111558. [Google Scholar] [CrossRef]
- Li, J.; Lei, H.; Song, H.; Lai, T.; Xu, X.; Shi, X. 1-methylcyclopropene (1-MCP) suppressed postharvest blue mold of apple fruit by inhibiting the growth of Penicillium expansum. Postharvest Biol. Technol. 2017, 125, 59–64. [Google Scholar] [CrossRef]
- Zhou, X.; Tang, Y.; Shi, Y.; Zhou, H. Mechanism of 1-MCP treatment in induced resistance to gray mold of apples during low-temperature storage. Food Sci. 2016, 37, 254–260. [Google Scholar] [CrossRef]
- Saftner, R.A.; Abbott, J.A.; Conway, W.S.; Barden, C.L. Effects of 1-Methylcyclopropene and Heat Treatments on Ripening and Postharvest Decay in ‘Golden Delicious’ Apples. J. Am. Soc. Hortic. Sci. 2003, 128, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Joyce, D.C.; Terry, L.A. 1-Methylcyclopropene treatment affects strawberry fruit decay. Postharvest Biol. Technol. 2001, 23, 227–232. [Google Scholar] [CrossRef]
- Wang, Z.; Yuan, G.; Pu, H.; Shan, S.; Zhang, Z.; Song, H.; Xu, X. 1-Methylcyclopropene suppressed the growth of Penicillium digitatum and inhibited the green mould in citrus fruit. J. Phytopathol. 2021, 169, 83–90. [Google Scholar] [CrossRef]
- Liu, H.; Jiang, W.; Zhou, L.; Wang, B.; Luo, Y. The effects of 1-methylcyclopropene on peach fruit (Prunus persica L. cv. Jiubao) ripening and disease resistance. Int. J. Food Sci. Technol. 2005, 40, 1–7. [Google Scholar] [CrossRef]
- Zhang, Z.; Tian, S.; Zhu, Z.; Xu, Y.; Qin, G. Effects of 1-methylcyclopropene(1-MCP) on ripening and resistance of jujube (Zizyphus jujuba cv. Huping) fruit against postharvest disease. LWT Food Sci and Technol. 2012, 45, 13–19. [Google Scholar] [CrossRef]
- Li, L.; Kashash, Y.; Goldenberg, L.; Sabag, A.; Doron-Faigenboim, A.; Porat, R. Effects of 1-methylcyclopropene on postharvest storage performance and the transcriptome of cactus pear fruit. Int. J. Food Sci. Technol. 2017, 52, 1801–1809. [Google Scholar] [CrossRef]
- Lin, Y.; Sun, J.; Chen, Y.; Lin, Y.; Jiang, X.; Lin, H. Inhibition of postharvest disease and induction of defence-related enzymes by paper containing 1-methlcyclopropene(1-MCP) in averrhoa carambola fruit. Chin. J. Trop. Crops. 2016, 37, 1172–1176. [Google Scholar]
- Cao, S.; Zheng, Y. Effect of 1-methylcyclopropene on anthracnose rot caused by Colletotrichum acutatum and disease resistance in loquat fruit. J. Sci. Food Agric. 2010, 90, 2289–2294. [Google Scholar] [CrossRef]
- Zeng, L.; Shi, L.; Lin, H.; Lin, Y.; Lin, Y.; Wang, H. Paper-containing 1-methylcyclopropene treatment suppresses fruit decay of fresh Anxi persimmons by enhancing disease resistance. Food Qual. Saf. 2021, 5, fyab007. [Google Scholar] [CrossRef]
- Hofman, P.J.; Jobin-Décor, M.; Meiburg, G.F.; Macnish, A.J.; Joyce, D.C. Ripening and quality responses of avocado, custard apple, mango and papaya fruit to 1-methylcyclopropene. Aust. J. Exp. Agric. 2001, 41, 567–572. [Google Scholar] [CrossRef]
- Sun, X.; Wang, W.; Li, Z.; Wang, Z.; Zhang, Z. Effects of 1-MCP on cold storage of dangshangsuii pears. Storage Process 2001, 6, 14–17. [Google Scholar] [CrossRef]
- Porat, R.; Weiss, B.; Cohen, L.; Daus, A.; Goren, R.; Droby, S. Effects of ethylene and 1-methylcyclopropene on the postharvest qualities of ‘Shamouti’ oranges. Postharvest Biol. Technol. 1999, 15, 155–163. [Google Scholar] [CrossRef]
- Chai, J.; Wang, Y.; Liu, Y.; Yong, K.; Liu, Z. 1-MCP extends the shelf life of ready-to-eat ‘Hayward’ and ‘Qihong’ kiwifruit stored at room temperature. Sci. Hortic. 2021, 289, 110437. [Google Scholar] [CrossRef]
- Mansoor, S.; Wani, O.A.; Lone, J.K.; Manhas, S.; Kour, N.; Alam, P.; Ahmad, A.; Ahmad, P. Reactive Oxygen Species in Plants: From Source to Sink. Antioxidants 2022, 11, 225. [Google Scholar] [CrossRef]
- Jones, A.M. Surprising Signals in Plant Cells. Science 1994, 263, 183–184. [Google Scholar] [CrossRef]
- Egea, I.; Flores, F.B.; Martínez-Madrid, M.C.; Romojaro, F.; Sánchez-Bel, P. 1-Methylcyclopropene affects the antioxidant system of apricots (Prunus armeniaca L. cv. Búlida) during storage at low temperature. J. Sci. Food Agric. 2010, 90, 549–555. [Google Scholar] [CrossRef]
- Chiriboga, M.-A.; Bordonaba, J.G.; Schotsmans, W.C.; Larrigaudière, C.; Recasens, I. Antioxidant potential of ‘Conference’ pears during cold storage and shelf life in response to 1-methylcyclopropene. LWT-Food Sci. Technol. 2013, 51, 170–176. [Google Scholar] [CrossRef]
- Xia, Y.; Zhuo, R.; Li, B.; Tian, S. Effects of 1-methylcyclopropene on disease resistance of red-fleshed kiwifruit during long-term cold storage and the possible mechanisms. N. Z. J. Crop. Hortic. 2020, 49, 182–195. [Google Scholar] [CrossRef]
- Shi, L.; Shen, L.; Yu, M.; Ouyang, L.; Fan, B.; Sheng, J. Effects of 1-methylcyclopropene treatment on the shelf-life quality of strawberry cv. Jingnong 1st. Acta Hortic. 2008, 768, 311–315. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, X.; Fu, M.; Chen, Q.; Muzammil, J.M. Chlorine dioxide fumigation generated by a solid releasing agent enhanced the efficiency of 1-MCP treatment on the storage quality of strawberry. J. Food Sci. Technol. 2018, 55, 2003–2010. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Huber, D.J.; Rao, J. Antioxidant systems of ripening avocado (Persea americana Mill.) fruit following treatment at the preclimacteric stage with aqueous 1-methylcyclopropene. Postharvest Biol. Technol. 2013, 76, 58–64. [Google Scholar] [CrossRef]
- Raji, M.R.; Lotfi, M.; Tohidfar, M.; Ramshini, H.; Sahebani, N.; Aalifar, M.; Baratian, M.; Mercati, F.; De Michele, R.; Carimi, F. Multiple fungal diseases resistance induction in Cucumis melo through co-transformation of different pathogenesis related (PR) protein genes. Sci. Hortic. 2022, 297, 110924. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Van Strien, E.A. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of Inducible Defense-related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoniw, J.F.; Ritter, C.E.; Pierpoint, W.S.; Loon, L.V.C. Comparison of Three Pathogenesis-related Proteins from Plants of Two Cultivars of Tobacco Infected with TMV. J. Gen. Virol. 1980, 47, 79–87. [Google Scholar] [CrossRef]
- Ohshima, M.; Matsuoka, M.; Yamamoto, N.; Tanaka, Y.; Kano-Murakami, Y.; Ozeki, Y.; Kato, A.; Harada, N.; Ohashi, Y. Nucleotide sequence of the PR-1 gene of Nicotiana tabacum. FEBS Lett. 1987, 225, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Tornero, P.; Gadea, J.; Conejero, V.; Vera, P. Two PR-1 Genes from Tomato Are Differentially Regulated and Reveal a Novel Mode of Expression for a Pathogenesis-Related Gene During the Hypersensitive Response and Development. Mol. Plant-Microbe Interactions 1997, 10, 624–634. [Google Scholar] [CrossRef] [Green Version]
- Maleck, K.; Levine, A.M.; Eulgem, T.; Morgan, A.; Schmid, J.; Lawton, K.A.; Dangl, J.L.; Dietrich, R.A. The transcriptome of Arabidopsis thaliana during systemic acquired resistance. Nat. Genet. 2000, 26, 403–410. [Google Scholar] [CrossRef]
- Liu, Q.; Xue, Q. Computational identification of novel PR-1-type genes in Oryza sativa. J. Genet. Genom. 2006, 85, 193–198. [Google Scholar] [CrossRef]
- Joshi, V.; Joshi, N.; Vyas, A.; Jadhav, S.K. 25—Pathogenesis-Related Proteins: Role in Plant Defense. In Biocontrol Agents and Secondary Metabolites, 1st ed.; Woodhead Publishing: Sawston, UK; Cambridge, MA, USA, 2021; p. 573. [Google Scholar]
- Mitsuhara, I.; Iwai, T.; Seo, S.; Yanagawa, Y.; Kawahigasi, H.; Hirose, S.; Ohkawa, Y.; Ohashi, Y. Characteristic expression of twelve rice PR1 family genes in response to pathogen infection, wounding, and defense-related signal compounds (121/180). Mol. Genet. Genom. Genom. 2008, 279, 415–427. [Google Scholar] [CrossRef]
- Wang, Y.; Tan, L.-X.; Xu, Z.-Q.; Jiao, Y.-X.; Zhu, D.-X.; Yang, Y.-S.; Wei, J.-F.; Sun, J.-L.; Tian, M. Identification and characterization of natural PR-1 protein as major allergen from Humulus japonicus pollen. Mol. Immunol. 2023, 153, 170–180. [Google Scholar] [CrossRef]
- Alexander, D.C.; Goodman, R.M.; Gut-Rella, M.; Glascock, C.B.; Weymann, K.; Friedrich, L.; Maddox, D.W.; Ahl-Goy, P.; Luntz, T.L.; Ward, E.R. Increased tolerance to two oomycete pathogens in transgenic tobacco expressing pathogenesis-related protein 1a. Proc. Natl. Acad. Sci. USA 1993, 90, 7327–7331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niderman, T.; Genetet, I.; Bruyere, T.B.L.; Gees, R.; Stintzi, A.; Legrand, M.; Fritig, B.; Mosinger, E. Pathogenesis-Related PR-1 Proteins Are Antifungal (Isolation and Characterization of Three 14-Kilodalton Proteins of Tomato and of a Basic PR-1 of Tobacco with Inhibitory Activity against Phytophthora infestans). Plant Physiol. 1995, 108, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, J.K.; Hwang, B.K. Induction of enhanced disease resistance and oxidative stress tolerance by overexpression of pepper basic PR-1 gene in Arabidopsis. Physiol. Plant. 2005, 124, 267–277. [Google Scholar] [CrossRef]
- Dabravolski, S.A.; Frenkel, Z. Diversity and evolution of pathogenesis-related proteins family 4 beyond plant kingdom. Plant Gene 2021, 26, 100279. [Google Scholar] [CrossRef]
- Rabari, A.; Ruparelia, J.; Jha, C.K.; Sayyed, R.Z.; Mitra, D.; Priyadarshini, A.; Senapati, A.; Panneerselvam, P.; Mohapatra, P.K.D. Articulating beneficial rhizobacteria mediated plant defenses through induced systemic resistance. Pedosphere 2022. [Google Scholar] [CrossRef]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiol. Res. 2018, 212–213, 29–37. [Google Scholar] [CrossRef]
- Menezes, S.P.; Silva, E.M.D.A.; Lima, E.M.; de Sousa, A.O.; Andrade, B.S.; Lemos, L.S.L.; Gramacho, K.P.; Gesteira, A.D.S.; Pirovani, C.P.; Micheli, F. The pathogenesis-related protein PR-4b from Theobroma cacao presents RNase activity, Ca2+ and Mg2+ dependent-DNase activity and antifungal action on Moniliophthora perniciosa. BMC Plant Biol. 2014, 14, 161. [Google Scholar] [CrossRef] [Green Version]
- Sinha, M.; Singh, R.P.; Kushwaha, G.S.; Iqbal, N.; Singh, A.; Kaushik, S.; Kaur, P.; Sharma, S.; Singh, T.P. Current Overview of Allergens of Plant Pathogenesis Related Protein Families. Sci. World J. 2014, 2014, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loake, G.J.; Grant, M. Salicylic acid in plant defence—The players and protagonists. Curr. Opin. Plant Biol. 2007, 10, 466–472. [Google Scholar] [CrossRef]
- Appu, M.; Ramalingam, P.; Sathiyanarayanan, A.; Huang, J. An overview of plant defense-related enzymes responses to biotic stresses. Plant Gene 2021, 27, 100302. [Google Scholar] [CrossRef]
- Vaghela, B.; Vashi, R.; Rajput, K.; Joshi, R. Plant chitinases and their role in plant defense: A comprehensive review. Enzym. Microb. Technol. 2022, 159, 110055. [Google Scholar] [CrossRef]
- Anusuya, S.; Sathiyabama, M. Identification of defence proteins from the seed exudates of Cicer arietinum L. and its effect on the growth of Fusarium oxysporumf. sp. ciceri. Arch. Phytopathol. 2014, 47, 1611–1620. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, J.; Li, Z.; Ye, X.; Dong, W.; Jiang, M.; Huang, Y.; Cui, Z. Enzymatic properties of a multi-specific β-(1,3)-glucanase from Corallococcus sp. EGB and its potential antifungal applications. Protein Expr. Purif. 2019, 164, 105481. [Google Scholar] [CrossRef]
- Liu, L.; Fu, Y. Chitinase and its role in plant disease resistance. Mod Agric. 2003, 5, 5–7. [Google Scholar] [CrossRef]
- de Sousa, A.E.D.; Lopes, M.M.D.A.; Moreira, A.D.R.; Macedo, J.J.N.; Moura, C.F.H.; de Aragão, F.A.S.; Zocolo, G.; de Miranda, M.R.A.; Silva, E.D.O. Induction of postharvest resistance in melon using pulsed light as abiotic stressor. Sci. Hortic. 2019, 246, 921. [Google Scholar] [CrossRef]
- Kim, Y.; Youn, J.; Roh, J.; Kim, J.; Kim, S.; Kim, T. Brassinosteroids enhance salicylic acid-mediated immune responses by in-hibiting BIN2 phosphorylation of clade I TGA transcription factors in Arabidopsis. Mol. Plant. 2022, 15, 991. [Google Scholar] [CrossRef]
- Niki, T.; Mitsuhara, I.; Seo, S.; Ohtsubo, N.; Ohashi, Y. Antagonistic Effect of Salicylic Acid and Jasmonic Acid on the Expression of Pathogenesis-Related (PR) Protein Genes in Wounded Mature Tobacco Leaves. Plant Cell Physiol. 1998, 39, 500–507. [Google Scholar] [CrossRef]
- Li, Z.T.; Dhekney, S.A.; Gray, D.J. PR-1 gene family of grapevine: A uniquely duplicated PR-1 gene from a Vitis interspecific hybrid confers high level resistance to bacterial disease in transgenic tobacco. Plant Cell Rep. 2010, 30, 1–11. [Google Scholar] [CrossRef]
- Veselova, S.V.; Burkhanova, G.F.; Nuzhnaya, T.V.; Maksimov, I.V. Roles of ethylene and cytokinins in development of defense responses in Triticum aestivum plants infected with Septoria nodorum. Russ. J. Plant Physiol. 2016, 63, 609–619. [Google Scholar] [CrossRef]
- Dou, J.-F.; Kou, X.-H.; Wu, C.-E.; Fan, G.-J.; Li, T.-T.; Li, X.-J.; Zhou, D.-D.; Yan, Z.-C.; Zhu, J.-P. Recent advances and development of postharvest management research for fresh jujube fruit: A review. Sci. Hortic. 2023, 310, 111769. [Google Scholar] [CrossRef]
- Win, N.M.; Yoo, J.; Naing, A.H.; Kwon, J.-G.; Kang, I.-K. 1-Methylcyclopropene (1-MCP) treatment delays modification of cell wall pectin and fruit softening in “Hwangok” and “Picnic” apples during cold storage. Postharvest Biol. Technol. 2021, 180, 111599. [Google Scholar] [CrossRef]
- Lohani, S.; Trivedi, P.K.; Nath, P. Changes in activities of cell wall hydrolases during ethylene-induced ripening in banana: Effect of 1-MCP, ABA, and IAA. Postharvest Biol. Technol. 2004, 31, 119–126. [Google Scholar] [CrossRef]
- Wei, J.; Zhu, X.; Yuan, J.; Liu, C. Effects of 1-MCP on fruit’s starch and the components of cell-wall material during post-harvest of gala apples. Acta Agric. Boreali-Sin. 2008, 1, 121–124. [Google Scholar] [CrossRef]
- Fu, Y.; Chen, M.; Liu, K.; Chen, J. Effects of the second treatment with 1-MCP on post-harvest physiological and bio-chemical characteristics of kiwifruit. J. Fruit Sci. 2007, 1, 43–48. [Google Scholar] [CrossRef]
- Jiang, N.; Rao, J.; Fu, R. PG enzyme activity and the expression of the gene DkPG1 in the recovered softening of persimmon fruit. Hort. J. 2010, 9, 1507–1512. [Google Scholar]
- Sharma, S.; Sharma, R.R.; Pal, R.K.; Paul, V.; Dahuja, A. 1-Methylcyclopropene influences biochemical attributes and fruit softening enzymes of ‘Santa Rosa’ Japanese plum (Prunus salicina Lindl.). J. Plant Biochem. Biotechnol. 2011, 21, 295–299. [Google Scholar] [CrossRef]
- Itai, A.; Igori, T.; Fujita, N.; Egusa, M.; Kodama, M.; Murayama, H. Ethylene Analoge and 1-Methylcyclopropene Enhance Black Spot Disease Development in Pyrus pyrifolia Nakai. HortScience 2012, 47, 228–231. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Leverentz, B.; Conway, W.S.; Saftner, R.A.; Reed, A.; Camp, M.J. Control of bitter rot and blue mold of apples by integrating heat and antagonist treatments on 1-MCP treated fruit stored under controlled atmosphere conditions. Postharvest Biol. Technol. 2003, 29, 129–143. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, W.; Dong, W.; Ding, D.; Jia, X.; Zhang, Z. Research of 1-MCP treatment effects on fresh-keeping of ‘Lubaoshi’ pear. Storage Process 2007, 6, 9–12. [Google Scholar] [CrossRef]
- Endo, J.-I.; Takahashi, W.; Ikegami, T.; Beppu, T.; Tanaka, O. Induction of Flowering by Inducers of Systemic Acquired Resistance in the Lemna Plant. Biosci. Biotechnol. Biochem. 2009, 73, 183–185. [Google Scholar] [CrossRef]
- Blanco-Herrera, F.; Salinas, P.; Cecchini, N.M.; Jordana, X.; Van Hummelen, P.; Alvarez, M.E.; Holuigue, L. Early genomic responses to salicylic acid in Arabidopsis. Plant Mol. Biol. 2009, 70, 79–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, S.; Mallick, N.; Mitra, A. Salicylic acid-induced resistance to Fusarium oxysporum f. sp. lycopersici in tomato. Plant Physiol. Biochem. 2009, 47, 642–649. [Google Scholar] [CrossRef]
- Da Rocha Neto, A.C.; Luiz, C.; Maraschin, M.; Di Piero, R.M. Efficacy of salicylic acid to reduce Penicillium expansum inoculum and preserve apple fruits. Int. J. Food Microbiol. 2016, 221, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Zeier, J. Metabolic regulation of systemic acquired resistance. Curr. Opin. Plant Biol. 2021, 62, 102050. [Google Scholar] [CrossRef]
- Da Rocha Neto, A.C.; Maraschin, M.; Di Piero, R.M. Antifungal activity of salicylic acid against Penicillium expansum and its possible mechanisms of action. Int. J. Food Microbiol. 2015, 215, 64–70. [Google Scholar] [CrossRef]
- Iqbal, Z.; Singh, Z.; Khangura, R.K.; Ahmad, S. Management of citrus blue and green moulds through application of organic elicitors. Australas. Plant Pathol. 2011, 41, 69–77. [Google Scholar] [CrossRef]
- Panahirad, S.; Zaare-Nahandi, F.; Safaralizadeh, R.; Alizadeh-Salteh, S. Postharvest Control of Rhizopus stolonifer in Peach (Prunus persica L. Batsch) Fruits Using Salicylic Acid. J. Food Saf. 2012, 32, 502–507. [Google Scholar] [CrossRef]
- Cao, J.; Yan, J.; Zhao, Y.; Jiang, W. Effects of postharvest salicylic acid dipping on Alternaria rot and disease resistance of jujube fruit during storage. J. Sci. Food Agric. 2013, 93, 3252–3258. [Google Scholar] [CrossRef]
- Yatin, Z. Induction of disease resistance and phenylpropanoid metabolism in apricot fruits by pre-harvest salicylic acid treatment. J. Food Sci. 2015, 36, 216–220. [Google Scholar] [CrossRef]
- Yang, Z.; Cao, S.; Cai, Y.; Zheng, Y. Combination of salicylic acid and ultrasound to control postharvest blue mold caused by Penicillium expansum in peach fruit. Innov. Food Sci. Emerg. Technol. 2011, 12, 310–314. [Google Scholar] [CrossRef]
- Shan, W.; Chen, J.-Y.; Kuang, J.-F.; Lu, W.-J. Banana fruit NAC transcription factor MaNAC5 cooperates with MaWRKYs to enhance the expression of pathogenesis-related genes against Colletotrichum musae. Mol. Plant Pathol. 2016, 17, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Kuang, J.-F.; Wang, F.-Y.; Chen, L.; Hong, K.-Q.; Xiao, Y.-Y.; Xie, H.; Lu, W.-J.; Chen, J.-Y. Molecular characterization of PR and WRKY genes during SA- and MeJA-induced resistance against Colletotrichum musae in banana fruit. Postharvest Biol. Technol. 2013, 79, 62–68. [Google Scholar] [CrossRef]
- Zeng, K.; Cao, J.; Jiang, W. Enhancing disease resistance in harvested mango (Mangifera indica L. cv. ‘Matisu’) fruit by salicylic acid. J. Sci. Food Agric. 2006, 86, 694–698. [Google Scholar] [CrossRef]
- He, J.; Ren, Y.; Chen, C.; Liu, J.; Liu, H.; Pei, Y. Defense Responses of Salicylic Acid in Mango Fruit Against Postharvest Anthracnose, Caused by Colletotrichum gloeosporioides and its Possible Mechanism. J. Food Saf. 2017, 37, e12294. [Google Scholar] [CrossRef]
- Mustafa, M.A.; Ali, A.; Seymour, G.B.; Tucker, G.A. The role of the ubiquitous phenolic compound ’salicylic acid’ in chilling tolerance of carambola. Acta Hortic. 2015, 1079, 679–683. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, X.; Li, B.; Zhang, Q.; Liang, W.; Wang, C. Salicylic acid confers enhanced resistance to Glomerella leaf spot in apple. Plant Physiol. Biochem. 2016, 106, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Tian, S. Effects of pre- and post-harvest application of salicylic acid or methyl jasmonate on inducing disease resistance of sweet cherry fruit in storage. Postharvest Biol. Technol. 2005, 35, 253–262. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, J.; Bian, W.; Yang, X.; Ye, L.; He, S.; Song, X. Control Efficacy of Salicylic Acid Microcapsules against Postharvest Blue Mold in Apple Fruit. Molecules 2022, 27, 8108. [Google Scholar] [CrossRef]
- Moosa, A.; Sahi, S.T.; Khan, S.A.; Malik, A.U. Salicylic acid and jasmonic acid can suppress green and blue moulds of citrus fruit and induce the activity of polyphenol oxidase and peroxidase. Folia Hortic. 2019, 31, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Wang, F.; Lu, Y.; Deng, J. Combination of chitosan and salicylic acid to control postharvest green mold caused by Penicillium digitatum in grapefruit fruit. Sci. Hortic. 2018, 233, 54–60. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, Y. Effects of salicylic acid on the expression of SOD, PPO isozymes and NPR1 in pear. Hortic. Plant J. 2013, 40, 41–48. [Google Scholar] [CrossRef]
- Zhao, X.; Zhao, Y.; Wang, H.; Song, P. Effect of salicylic acid on quality and physiological character of postharvest pear (Pyrus pyrifolia cv. xingshui) fruit. Hubei Agric. Sci. 2009, 48, 1165–1167. [Google Scholar] [CrossRef]
- Zhu, F.; Chen, J.; Xiao, X.; Zhang, M.; Yun, Z.; Zeng, Y.; Xu, J.; Cheng, Y.; Deng, X. Salicylic acid treatment reduces the rot of postharvest citrus fruit by inducing the accumulation of H2O2, primary metabolites and lipophilic polymethoxylated flavones. Food Chem. 2016, 207, 68–74. [Google Scholar] [CrossRef]
- Mo, Y.; Gong, D.; Liang, G.; Han, R.; Xie, J.; Li, W. Enhanced preservation effects of sugar apple fruits by salicylic acid treatment during post-harvest storage. J. Sci. Food Agric. 2008, 88, 2693–2699. [Google Scholar] [CrossRef]
- Cao, J.; Zeng, K.; Jiang, W. Enhancement of Postharvest Disease Resistance in Ya Li Pear (Pyrus bretschneideri) Fruit by Salicylic Acid Sprays on the Trees during Fruit Growth. Eur. J. Plant Pathol. 2005, 114, 363–370. [Google Scholar] [CrossRef]
- Huang, R.-H.; Liu, J.-H.; Lu, Y.-M.; Xia, R.-X. Effect of salicylic acid on the antioxidant system in the pulp of ‘Cara cara’ navel orange (Citrus sinensis L. Osbeck) at different storage temperatures. Postharvest Biol. Technol. 2008, 47, 168–175. [Google Scholar] [CrossRef]
- Mora-Herrera, M.E.; López-Delgado, H.A.; Castillo-Morales, A.; Foyer, C.H. Salicylic acid and H2O2 function by independent pathways in the induction of freezing tolerance in potato. Physiol. Plant. 2005, 125, 430–440. [Google Scholar] [CrossRef]
- Chen, Z.; Ricigliano, J.W.; Klessig, D.F. Purification and characterization of a soluble salicylic acid-binding protein from tobacco. Proc. Natl. Acad. Sci. USA 1993, 90, 9533–9537. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Wang, Y.; Liu, X.; Guo, X.; Wang, G.; Li, L. Effect of pre-harvest salicylic acid spray treatment on fruit quality and antioxidant capacity of raspberry during post-harvest storge. J. Food Sci. 2010, 31, 308–312. [Google Scholar]
- Chen, Y.; Sun, J.; Lin, H.; Lin, M.; Lin, Y.; Wang, H.; Hung, Y.-C. Salicylic acid treatment suppresses Phomopsis longanae Chi-induced disease development of postharvest longan fruit by modulating membrane lipid metabolism. Postharvest Biol. Technol. 2020, 164, 111168. [Google Scholar] [CrossRef]
- Hong, X.; Hou, X.; Zheng, Y.; Zhang, X. Induction effect of salicylic acid on resistance to powdery mildew of Jiulixiang. For. Pro. Des. 2013, 1, 125–127. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; War, M.Y.; Ignacimuthu, S. Role of salicylic acid in induction of plant defense system in chickpea (Cicer arietinum L.). Plant Signal. Behav. 2011, 6, 1787–1792. [Google Scholar] [CrossRef] [Green Version]
- Cheong, Y.H.; Moon, B.C.; Kim, J.K.; Kim, C.Y.; Kim, M.C.; Kim, I.H.; Park, C.Y.; Kim, J.C.; Park, B.O.; Koo, S.C.; et al. BWMK1, a Rice Mitogen-Activated Protein Kinase, Locates in the Nucleus and Mediates Pathogenesis-Related Gene Expression by Activation of a Transcription Factor1. Plant Cell Physiol. 2003, 132, 1961–1972. [Google Scholar] [CrossRef] [Green Version]
- Koo, S.C.; Moon, B.C.; Kim, J.K.; Kim, C.Y.; Sung, S.J.; Kim, M.C.; Cho, M.J.; Cheong, Y.H. OsBWMK1 mediates SA-dependent defense responses by activating the transcription factor OsWRKY33. Biochem. Biophys. Res. Commun. 2009, 387, 365–370. [Google Scholar] [CrossRef]
- Li, C.; Lei, C.; Wang, K.; Tan, M.; Xu, F.; Wang, J.; Zheng, Y. MADS2 regulates priming defence in postharvest peach through combined salicylic acid and abscisic acid signaling. J. Exp. Bot. 2022, 73, 3787–3806. [Google Scholar] [CrossRef]
- Mahesh, H.M.; Murali, M.; Pal, M.A.C.; Melvin, P.; Sharada, M. Salicylic acid seed priming instigates defense mechanism by inducing PR-Proteins in Solanum melongena L. upon infection with Verticillium dahliae Kleb. Plant Physiol. Biochem. 2017, 117, 12–23. [Google Scholar] [CrossRef]
- Shen, Y.; Yang, H. Effect of preharvest chitosan-g-salicylic acid treatment on postharvest table grape quality, shelf life, and resistance to Botrytis cinerea -induced spoilage. Sci. Hortic. 2017, 224, 367–373. [Google Scholar] [CrossRef]
- Zhang, X.C.; Ren, A.; Zhang, P.; Yang, H.H.; Zhao, T.T.; Xu, X.Y.; Jiang, J.B.; Li, J.F. Identification of salicylic acid conferred resistance genes against gray leaf spot disease in tomato. Int. J. Agric. Biol. 2020, 23, 109–116. [Google Scholar] [CrossRef]
- Li, L.; Zou, Y. Induction of disease resistance by salicylic acid and calcium ion against Botrytis cinerea in tomato (Lycopersicon esculentum). Emir. J. Food Agric. 2017, 29, 78. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-J.; Noh, S.-I.; Choi, C.; Lim, B.-S.; Ahn, Y.-J.; Chun, J.-P. Effects of Salicylic Acid and 1-Methylcyclopropene on Physiological Disorders and Berry Quality in ‘Campbell Early’ Table Grapes. Prot. Hortic. Plant Fact. 2019, 28, 218–224. [Google Scholar] [CrossRef]
- Xu, F.; Liu, Y.; Xu, J.; Fu, L. Influence of 1-methylcyclopropene (1-MCP) combined with salicylic acid (SA) treatment on the postharvest physiology and quality of bananas. J. Food Process. Preserv. 2019, 43, 13880. [Google Scholar] [CrossRef]
- Luo, D.; Zhai, G.; Cao, S.; Zhu, T.; Chen, R.; Xiao, Q.; Zhang, D.; Ba, L. Effect of preharvest salicylic acid combined with postharvest 1-MCP treatment on quality and antioxidant ablity of plum fruit during storage. Science 2022, 43, 327–333. [Google Scholar] [CrossRef]
- Li, Y.; He, H.; Hou, Y.Y.; Kelimu, A.; Wu, F.; Zhao, Y.; Shi, L.; Zhu, X. Salicylic acid treatment delays apricot (Prunus armeniaca L.) fruit softening by inhibiting ethylene biosynthesis and cell wall degradation. Sci. Hortic. 2022, 300, 111061. [Google Scholar] [CrossRef]


{kind=link}
| Fruits | Treatment Concentration | Fungi | Reference |
|---|---|---|---|
| “Fuji” apple | 1 μL/L | Botrytis cinerea | [6] |
| “Everest” strawberry | 1 μL/L | Botrytis cinerea | [13] |
| Jujube | 1 μL/L | Botrytis cinerea | [16] |
| “Golden Delicious” apple | 0.3 μL/L | Colletotrichum acutatum | [12] |
| Cactus pear | 1 μL/L | Colletotrichum gloeosporioides | [17] |
| Carambola | 0.6 μL/L | Colletotrichum gloeosporioides | [18] |
| Loquat | 50 nL/L | Colletotrichum gloeosporioides | [19] |
| “Anxi” persimmon | 1.35 μL/L | Gloeosporium kaki | [20] |
| “Dangshan” pear | 1 μL/L | Penicillium digitatum | [21] |
| “Emerald” pear | 1 μL/L | Penicillium digitatum | [22] |
| Citrus | 5 μL/L | Penicillium digitatum | [14] |
| “Fuji” apple | 5 μL/L | Penicillium expansum | [10] |
| “Golden Delicious” apple | 0.3 μL/L | Penicillium expansum | [23] |
| Peach | 0.6 μL/L | Penicillium expansum | [15] |
| “Hongyang” kiwifruit | 0.8 μL/L | Phomopsis sp. | [24] |
| Fruits | Treatment Concentration | Fungi | Reference |
|---|---|---|---|
| Apricot | 1 mmol/L | Alternaria alternata | [82] |
| Jujube | 2 mmol/L | Alternaria alternata | [81] |
| Tomato | 2 mmol/L | Botrytis cinerea | [83] |
| Banana | 2 mmol/L 2 mmol/L | Colletotrichum musae Colletotrichum gloeosporioides | [84] [85] |
| Mango | 1 mmol/L 5 mmol/L | Colletotrichum gloeosporioides Colletotrichum gloeosporioides | [86] [87] |
| Carambola | 1 mmol/L | Colletotrichum gloeosporioides | [88] |
| Tomato | 200 μmol/L | Fusarium oxysporum | [76] |
| Apple | 0.2 mmol/L | Glomerella cingulata | [89] |
| Cherry | 2 mmol/L | Monilinia fructicola | [90] |
| Apple | 0.3 mmol/L | Penicillium expansum | [91] |
| Peach | 0.05 mmol/L | Penicillium expansum | [83] |
| Citrus | 2.5 mmol/L | Penicillium expansum | [92] |
| Apple | 2.5 mmol/L | Penicillium digitatum | [76,78] |
| Citrus | 3 mmol/L | Penicillium digitatum | [79] |
| Grapefruit | 2 mmol/L | Penicillium digitatum | [93] |
| Pear | 0.2 mmol/L | Physalospora piricola | [94] |
| Peach | 5 mmol/L | Rhizopus stolonifer | [80] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, X.; Fang, J.; Fu, M.; Jiao, W.; Ren, P.; Yang, X. The Role of 1-methylcyclopropylene (1-MCP) and Salicylic Acid (SA) in Induced Resistance of Postharvest Fruits. Horticulturae 2023, 9, 108. https://doi.org/10.3390/horticulturae9010108
Meng X, Fang J, Fu M, Jiao W, Ren P, Yang X. The Role of 1-methylcyclopropylene (1-MCP) and Salicylic Acid (SA) in Induced Resistance of Postharvest Fruits. Horticulturae. 2023; 9(1):108. https://doi.org/10.3390/horticulturae9010108
Chicago/Turabian StyleMeng, Xiangzhi, Jiazhen Fang, Maorun Fu, Wenxiao Jiao, Pengfei Ren, and Xiaoying Yang. 2023. "The Role of 1-methylcyclopropylene (1-MCP) and Salicylic Acid (SA) in Induced Resistance of Postharvest Fruits" Horticulturae 9, no. 1: 108. https://doi.org/10.3390/horticulturae9010108
APA StyleMeng, X., Fang, J., Fu, M., Jiao, W., Ren, P., & Yang, X. (2023). The Role of 1-methylcyclopropylene (1-MCP) and Salicylic Acid (SA) in Induced Resistance of Postharvest Fruits. Horticulturae, 9(1), 108. https://doi.org/10.3390/horticulturae9010108